
Indri Juliyarsi
Laboratory of Technology Animal Product Processing, Faculty of Animal Science, Andalas University, Limau Manis, Padang, Indonesia
LiveDNA: 62.16274
Puji Hartini
Department of Animal Science, Andalas University, Limau Manis, Padang, Indonesia
LiveDNA: 62.23061
Yuherman
Laboratory of Technology Animal Product Processing, Faculty of Animal Science, Andalas University, Limau Manis, Padang, Indonesia
Akmal Djamaan
Laboratory Biota Sumatera, Andalas University, Limau Manis, Padang, Indonesia
Arief
Laboratory of Technology Animal Product Processing, Faculty of Animal Science, Andalas University, Limau Manis, Padang, Indonesia
LiveDNA: 62.24390
Hendri Purwanto
Laboratory of Technology Animal Product Processing, Faculty of Animal Science, Andalas University, Limau Manis, Padang, Indonesia
Salam N. Aritonang
Laboratory of Technology Animal Product Processing, Faculty of Animal Science, Andalas University, Limau Manis, Padang, Indonesia
LiveDNA: 62.21859
James Hellyward
Laboratory of Technology Animal Product Processing, Faculty of Animal Science, Andalas University, Limau Manis, Padang, Indonesia
Endang Purwati
Laboratory of Technology Animal Product Processing, Faculty of Animal Science, Andalas University, Limau Manis, Padang, Indonesia
LiveDNA: 62.23209
Pakistan Journal of Nutrition
Year: 2018 | Volume: 17 | Issue: 10 | Page No.: 506-511







ABSTRACT
Background and Objective: Tempoyak is a traditional fermented condiment made from durian (Durio zibethinus) pulp. This condiment is made by mixing the durian pulp with salt and fermenting under partially anaerobic conditions at ambient temperature in a closed container. This study aimed to determine the characteristics of Lactic Acid Bacteria (LAB) in tempoyak from Padang Pariaman District, West Sumatra. Methodology: Experimental methods of measuring the chemical composition of tempoyak; isolation and purification of LAB; characterization of selected isolates; isolation of genomic 16S rRNA; microscopic, macroscopic, molecular identification and determination of antimicrobial action by the agar well diffusion method using E. coli, S. aureus and L. monocytogenes as indicator bacteria. Results: Tempoyak is composed of water, protein, fat and has an acidic taste due to its low pH (3.89) affects the water content (70.21%), protein content (5.04%) and fat content (6.11%). Gram staining showed that the isolated bacteria (bacil) are Gram-positive and catalase-negative. Moreover, antimicrobial activity was tested by the agar well diffusion method using E. coli, S. aureus and L. monocytogenes as indicator bacteria. The largest inhibitory zone was observed between the isolated Tempoyak Original (TO) sample and S. aureus (19.3 mm), followed by TO against L. monocytogenes (17.3 mm) and the smallest inhibitory zone was observed between the TO sample and E. coli (12.3 mm). Conclusion: The length of the PCR amplified DNA fragment was 1482 bp. The sequencing results from the isolated tempoyak TO showed that the LAB isolate was Lactobacillus fermentum strain CAU6337.
PDF Abstract XML References Citation
Received: January 17, 2018;
Accepted: July 12, 2018;
Published: September 15, 2018
Copyright: © 2018. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Indri Juliyarsi, Puji Hartini, Yuherman, Akmal Djamaan, Arief, Hendri Purwanto, Salam N. Aritonang, James Hellyward and Endang Purwati, 2018. Characterization of Lactic Acid Bacteria and Determination of Antimicrobial Activity in Tempoyak from Padang Pariaman District, West Sumatra, Indonesia. Pakistan Journal of Nutrition, 17: 506-511.
DOI: 10.3923/pjn.2018.506.511
URL: https://scialert.net/abstract/?doi=pjn.2018.506.511
DOI: 10.3923/pjn.2018.506.511
URL: https://scialert.net/abstract/?doi=pjn.2018.506.511
INTRODUCTION
Tempoyak is fermented durian pulp with a distinctive durian smell and creamy yellow colour and is widely consumed in both Malaysia and Indonesia as a side dish and condiment1-3. As a condiment, tempoyak is consumed with fish and vegetable dishes. This condiment is made by mixing the durian pulp with salt and fermenting under partially anaerobic conditions at ambient temperature in a closed container. Fermentation usually takes 4-7 days and the durian pulp texture changes from a solid to a semisolid with an acidic odour and dominant taste. Tempoyak acidity was reported to be approximately 2.8-3.6%4. Its sour taste is attributed to the acid produced by Lactic Acid Bacteria (LAB) during fermentation. Earlier studies have shown that LAB is the predominant microorganisms in tempoyak5,6. Lactobacillus species have been reported as the main LAB isolated from tempoyak made in Indonesia and Malaysia. It is expected that LAB and other microorganism strains vary depending on where the product is prepared. Wirawati7 isolated L. casei and L. corynebacterium from tempoyak in Indonesia.
The LAB provides several potential health and nutritional benefits and are known as "probiotics". Bacteria that live in human intestines and control the balance of intestinal microflora elicit physiological and beneficial health effects to the host and are referred to as "probiotics". Probiotics are living microorganisms, which upon ingestion in certain numbers, exert health benefits beyond inherent basic nutrition7. Probiotics are suggested to have the following properties and functions: Adhere to host epithelial tissues, acid resistance and bile tolerance, pathogen elimination or reduced pathogen adherence, acid production and production of hydrogen peroxide and bacteriocin that antagonize pathogen growth, safety, non-pathogenic and non-carcinogenic properties and improve intestinal microflora8.
Durian can be eaten fresh, used as a flavouring agent for ice cream or processed into candies and other delicacies; however, with consumers’ current desire for natural and healthy foods, isolating useful microorganisms from traditional fermented products has become the target of several investigations. Thus, this study aimed to identify the predominant lactic acid bacteria present in tempoyak made in Padang Pariaman District of West Sumatra, Indonesia, based on LAB phenotypic characteristics and antimicrobial activity.
MATERIALS AND METHODS
Fully ripened durian fruits were obtained from Padang Pariaman District of West Sumatra, Indonesia.
Fermented durian preparation: Tempoyak was prepared using durian pulp under anaerobic conditions, placed in a bottle and allowed to naturally ferment for 14 days at room temperature (28-34°C).
Tempoyak chemical composition: After naturally fermenting for 14 days at room temperature (28-34°C), the pH and moisture, protein and fat content in tempoyak samples were tested.
Lab isolation and purification: The LAB were isolated and purified according to the method described by Purwati et al.9. A tempoyak sample (1 g) was diluted in 9 mL of MRS broth. A 10–1 dilution was placed into the anaerobic jar and incubated for 48 h at 37°C. The 10–1 dilution was then placed into a test tube containing 9 mL of MRS broth solution and serially diluted in 10–7. Next, 100μL samples of the 107 dilution were spread on petri dishes containing MRS agar and incubated at 37°C for 48 h.
A single colony was then transferred by an ose needle to MRS agar to isolate the colony by the streak method and incubated for 24 h at 37°C.
Isolate characterization: Isolates were examined by Gram staining and catalase reaction tests and the cell shape was evaluated microscopically. Gas production from glucose was tested using a Durham tube and MRS broth to determine the fermentation type.
Antimicrobial activity: Antimicrobial resistance testing was performed with three test bacteria: Escherichia coli O157, Listeria monocytogenes and Staphylococcus aureus. The LAB culture (1 mL) was centrifuged at 10,000 rpm for 5 min at 27°C and the supernatant was used to test antimicrobial resistance. Nutrient Agar (NA) was added at 0.2% to test bacteria that were replicated, homogenized and incubated. Isolates were used and labelled. The LAB supernatant (50μL) was injected by micropipette and incubated at 37°C anaerobically. Clear, circular zones of inhibition were measured after 24 h.
Isolation of LAB genomic DNA and 16S rRNA: Bacteria that formed single colonies were inoculated in MRS broth and incubated at 37°C for 24 h at 80 rpm. The inoculant was transferred to a 2 mL microtube and centrifuged at 10,000 rpm for 3 min. Supernatants were discarded and the pellets were used for genomic DNA isolation. Pellets were added to 1 mL of Tris-EDTA (TE) (1 M Tris HCl, 0.5 M EDTA, pH 8) and centrifuged at 12,000 rpm for 5 min. The supernatant was transferred to a new Eppendorf tube and 1 mL of cold absolute ethanol was added. The supernatant was removed and the pellet was resuspended in distilled water. The DNA was stored at -20°C until use10.
The 16S rRNA reaction was performed. Genomic DNA from pure bacterial colonies was isolated and amplified by PCR. The DNA amplification reactions were performed in the Thermocycler Mupid-exU from IndoLab Utama, Jakarta used bacteria and archaea primers. According to Sunarto and Mahajoeno11, the Arch109f forward primer (5’-ACK GCT CAG TAA CAC GT-3’) and Ar912r reverse primer (5’-CTC CCC CGC CAA TTC CTT TA-3’) were utilized. The contents of one PCR reaction were 38.5μL of ddH2O, 0.5μL of F and R primers, 2μL of 2.5 mm dNTP, 0.5μL of Taq-polymerase, 5μL of buffer and 3μL of DNA template in an 0.5 mL Eppendorf tube. The PCR protocol was 35 cycles of predenaturation at 96°C for 5 min, denaturation at 96°C for 1 min, annealing at 55°C for 1 min, extension at 72°C for 3 min and a final extension at 72°C for 7 min. The PCR products were analyzed on a 1% agarose gel and visualized by ultraviolet illumination after adding 5μL to the gel. The DNA bands on the agar were cut and purified using the Protocol Promo Kit.
Purified products were sequenced by the Genetika Science Indonesia Laboratory in Jakarta. Subsequently, the 16S rRNA gene sequences of the isolate were submitted to the National Center for Biotechnology Information for a BLAST search. The MEGA version 6.0 (http://www.megasoftware.net) was used to create phylogenetic trees using the neighbour-joining (NJ) method.
RESULTS AND DISCUSSION
Tempoyak chemical composition: The pH and water, protein and fat content of the tempoyak are shown in Table 1.
Tempoyak is composed of protein, fat and has an acidic taste due to its low pH (Table 1). This low pH (3.89) affects the water content (70.21%), protein content (5.04%) and fat content (6.11%) of the tempoyak and is associated with the long shelf life of tempoyak. The low pH is due to the lactic acid produced during lactic acid bacteria growth. According to Usmiati and Setiyanto12, in traditional spontaneous fermentation, the microbes that are naturally present in the food will grow, leading to the succession or the substitution of the dominant microbes in the next process. The LAB produces more acid, thus lowering the pH to approximately 3.89.
LAB isolation and purification: The total LAB counts for the tempoyak from Padang Pariaman District were 16×108 CFU/g. Some bacteria were selected randomly from the MRS agar and further characterized for taxonomic properties.
| Table 1: | Tempoyak chemical composition |
 | |
Next, LAB with round, sleek and creamy characteristics were isolated and LAB morphology was evaluated macroscopically and microscopically.
Compared with previous reports on other natural dairy foods12, LAB appeared to be highly viable in tempoyak products, which is important for further functional food development. Therefore, it may be useful to further characterize the microbial diversity and composition of these conventional dairy products.
Characterizing selected isolates: Macroscopic observations (shapes, sizes and colours) showed that LAB formed round, creamy white colonies and convex elevations with slippery edges on MRS agar (MERCK).
Next, LAB was purified twice and refined as a bacterial stock for microscopic identification and to observe the physiological properties of the isolates. A Gram test was performed to determine whether the bacteria were Gram-positive or Gram-negative, as characterized by the bacteria absorbing colour reagents. Gram-positive bacteria (+) absorb the crystal violet reagent, whereas Gram-negative bacteria (-) absorb the safranin reagent and are stained red. The Gram stain results revealed that the LAB isolated from the durian pulp from Padang Pariaman District were rods (bacilli).
The Gram stain results for all LAB isolates were Gram-positive (+) because the bacteria absorbed the crystal violet and were rod-shaped. This is consistent with Unus13, who stated that Gram-positive bacteria will be stained purple with crystal violet and remain purple even after being washed with alcohol and exposed to safranin red. Gram-negative bacteria will stain red.
The LAB biochemical testing was performed using a catalase test to determine the ability of the bacteria to produce the catalase enzyme for oxygen tolerance. This biochemical test is performed by dropping hydrogen peroxide (H2O2) onto bacterial preparations on glass. The appearance of gas bubbles indicates that the bacteria are catalase-positive, while the absence of gas indicates that they are catalase-negative. The LAB are typically Gram-positive, catalase-negative and non-spore-forming. Catalase-negative isolates were characterized by the absence of gas bubbles, indicating H2O2 depletion. The results of this study were similar to those of Romadhon and Margino14, who reported catalase-negative LAB isolates from shrimp intestines. Ibrahim et al.15 also found catalase-negative LAB isolates from mangoes.
In this research study, we found that the LAB in tempoyak from Padang Pariaman are homofermentative. The catalase test, which is a biochemical test that indicates the type of fermentation, was used to classify LAB as homofermentative or heterofermentative.
Suryani et al.16 described two types of LAB fermentation: Homofermentative and heterofermentative. Homofermentative LAB produces only lactic acid as the main product of fermentation, while heterofermentative LAB also produce ethanol, other acids, such as acetic acid and CO2 gas. Gas production from LAB are tested in Durham tubes; LAB that produce bacterial lactic acid are homofermentative, while isolating that produce gas are heterofermentative. Devi et al.17 conducted a study of bactericidal activity in 6 Lactobacillus delbreukii isolates from commercial yogurt and found that they were homofermentative.
Antimicrobial activity: To select and obtain the most potent LAB isolates of the probiotic candidates and to determine their ability to suppress the growth of pathogenic bacteria such as L. monocytogenes, S. aureus and E. coli O157, the isolates were screened by using the diffusion well method. This study did not use positive controls, such as antibiotics, because the tests were performed to determine the ability of the isolates to inhibit the growth of pathogenic bacteria. The diameters of the clear zones formed on the isolates are shown in Table 2.
The largest inhibitory zone was obtained from the isolate TO against S. aureus (19.3 mm). The inhibitory zone was 17.3 mm against L. monocytogenes and the lowest inhibitory zone was observed against E. coli (12.3 mm). These three test bacteria have different characteristics. Based on Theron and Lues18, each test bacterium has its own resistance to different organic acids. L. monocytogenes is more susceptible to lactic acid than acetic acid; E. coli is highly susceptible to lactic acid and acetic acid and S. aureus exhibits higher acid resistance than the other bacteria. As reported by Lee et al.19, the ability of LAB to inhibit the growth of pathogenic bacteria is shown by the extent of the clear zone produced during antimicrobial testing and is related to the ability of LAB to produce secondary metabolites, such as lactate and bacteriocin.
The results showed that LAB isolates from Padang Pariaman District inhibited the growth of pathogenic bacteria that endanger humans, especially S. aureus. Based on the inhibitory zone for pathogenic bacteria, LAB probiotics can be used in treatments.
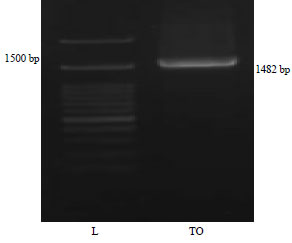 | |
| Fig. 1: | Electrophoresis of the PCR product from the tempoyak isolates from Padang Pariaman district |
| L = Ladder, TO = Isolated LAB | |
| Table 2: | Clear zone diameter, indicating antimicrobial activity (mm) |
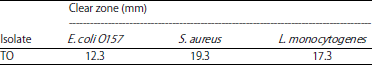 | |
Genomic isolation and 16S rRNA reaction of LAB: The electrophoresis results in Fig. 1 indicate that the 16S rRNA gene region of the fermented durian pulp of Padang Pariaman District was successfully amplified.
Successful amplification of the 16S rRNA gene was indicated by the appearance of a 1.5 kB PCR product, which was the expected fragment size when using the 27F forward primer AGAGTTTGATCCTGGCTGAG with the reverse primer 1492 R GTTTACCTTACGACTT. The sequencing result using a reverse primer obtained up to 885 nucleotides. A phylogenetic tree based on 16S rRNA gene sequence analysis is shown in Fig. 2.
Based on the BLAST search result, the bacterial isolate TO was confirmed to be 100% Lactobacillus fermentum strain CAU6337. The phylogenetic tree shows that the nearest distant neighbour is the Lactobacillus fermentum strain CAU6337. This indicates that the LAB isolated from tempoyak were Lactobacillus fermentum strain CAU6337. Hagstrom et al.20 suggested that isolates with a 16S rRNA sequence similarity over 97% may represent the same species, while sequence similarity between 93 and 97% indicates the same genus but different species.
Gram staining, PCR and BLAST analyses were performed on isolates of tempoyak from Padang Pariaman District. Gram staining showed that all isolates were Gram-positive bacilli. The catalase and fermentation type results were also uniform. PCR and BLAST results showed similarity with Lactobacillus fermentum.
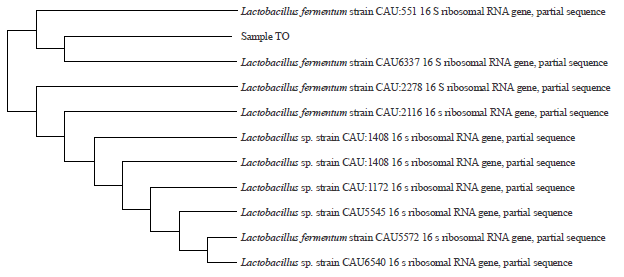 | |
| Fig. 2: | Phylogenetic tree based on the 16S rRNA gene sequence analysis |
This finding is consistent with Khumalawati21, who stated that bacteria live in vegetable waste and Lactobacillus fermentum is also found in vegetable waste. Lactic acid can be produced through spontaneous fermentation in the presence of Lactobacillus fermentum and occurs naturally under anaerobic conditions. Di Cagno et al.22 also previously stated that Lactobacillus fermentum is one of the most common species isolated from raw vegetables and fermented fruit.
Arief23 explained that Lactobacillus fermentum can ferment a few simple sugars, such as galactose, glucose, lactose, raffinose, sucrose and xylose. Khumalawati21 stated that simple sugars in fruit and vegetable waste become food for Lactobacillus fermentum, supporting bacterial growth and fermentation.
SIGNIFICANCE STATEMENT
This study identified the predominant lactic acid bacteria present in tempoyak. This study will help the researchers to uncover the benefits of indigenous LAB isolated from tempoyak and to determine their ability to suppress the growth of pathogenic bacteria such as L. monocytogenes, S. aureus and E. coli O157. Food industry can use these LABs for producing functional food products like fermented milk and other food stuffs.
CONCLUSION
The LAB was isolated from tempoyak from Padang Pariaman District. Gram staining showed that the isolates were Gram-positive, catalase-negative bacilli. The largest inhibitory zone was obtained from the isolate TO against S. aureus (19.3 mm), followed by that against L. monocytogenes (17.3 mm) and the smallest inhibitory zone was produced against E. coli O157 at 12.3 mm. The PCR results showed that the DNA fragment was 1482 bp. The sequencing results from the isolated tempoyak sample TO showed that the LAB isolate was Lactobacillus fermentum strain CAU6337.
ACKNOWLEDGMENT
The authors would like to thank the Post Graduate Research Team at Andalas University, Ministry of Research, Technology and Higher Education of the Republic of Indonesia for funding this study as well as the Andalas University Research and Community Services Institute.
REFERENCES
- Wandi, I.R. and Y.B. Che Man, 1996. Durian leather: Development, properties and storage stability. J. Food Qual., 19: 479-489.
CrossRefDirect Link - Leisner, J.J., M. Vancanneyt, K. Lefebvre, K. Vandemeulebroecke and B. Hoste et al., 2002. Lactobacillus durianis sp. nov., isolated from an acid-fermented condiment (Tempoyak) in Malaysia. Int. J. Syst. Evol. Microbiol., 52: 927-931.
CrossRefDirect Link - Leisner, J.J., M. Vancanneyt, G. Rusul, B. Pot, K. Lefebvre, A. Fresi and L.K. Tee, 2001. Identification of lactic acid bacteria constituting the predominating microflora in an acid-fermented condiment (Tempoyak) popular in Malaysia. Int. J. Food Microbiol., 63: 149-157.
CrossRefDirect Link - Amiza, M.A., J. Zakiah, L.K. Ng and K.W. Lai, 2006. Fermentation of tempoyak using isolated tempoyak culture. Res. J. Microbiol., 1: 243-254.
CrossRefDirect Link - Saito, T., 2004. Selection of useful probiotic lactic acid bacteria from the Lactobacillus acidophilus group and their applications to functional foods. Anim. Sci. J., 75: 1-13.
CrossRefDirect Link - Ibrahim, A., A. Fridayanti and F. Delvia, 2015. Isolation and identification of lactic acid bacteria (LAB) from mango (Mangifera indica). J. Ilmiah Manuntung, 1: 159-163.
Direct Link - Devi, M., L.J. Rebecca and S. Sumathy, 2013. Bactericidal activity of the lactic acid bacteria Lactobacillus delbreukii. J. Chem. Pharm. Res., 5: 176-180.
Direct Link - Lee, J.S., K.C. Lee, J.S. Ahn, T.I. Mheen, Y.R. Pyun and Y.H. Park, 2002. Weissella koreensis sp. nov., isolated from kimchi. Int. J. Syst. Evol. Microbiol., 52: 1257-1261.
CrossRefDirect Link - Hagstrom, A., J. Pinhassi and U.L. Zweifel, 2000. Biogeographical diversity among marine bacterioplankton. Aquat. Microb. Ecol., 21: 231-244.
CrossRefDirect Link - Di Cagno, R., M. de Angelis, A. Limitone, F. Minervini and P. Carnevali et al., 2006. Glucan and fructan production by sourdough Weissella cibaria and Lactobacillus plantarum. J. Agric. Food Chem., 54: 9873-9881.
CrossRefDirect Link - Arief, I.I., 2005. Characteristics and nutritional value of Dark Firm Dry beef (DFD) beef protein from fermented Lactobacillus plantarum isolated from beef. Final Report of Research Activity of Universities Competitive Grant XIII/I, Institute for Research and Community Empowerment, Bogor Agricultural University, Bogor.