
Iyiola Oluwakemi Owolabi
Interdisciplinary Graduate School of Nutraceutical and Functional Food, Prince of Songkla University, Hat-Yai, 90112 Songkla, Thailand
Chutha Takahashi Yupanqui
Interdisciplinary Graduate School of Nutraceutical and Functional Food, Prince of Songkla University, Hat-Yai, 90112 Songkla, Thailand
Sunisa Siripongvutikorn
Department of Food Technology, Faculty of Agro-Industry, Prince of Songkla University, Hat-Yai, 90112 Songkla, Thailand
LiveDNA: 66.22917
Pakistan Journal of Nutrition
Year: 2018 | Volume: 17 | Issue: 9 | Page No.: 411-420







ABSTRACT
New discoveries on the benefits of bioactive compounds derived from plants have prompted the modification and enrichment of foods and food components with the aim of promoting health benefits. The process of elicitation is beneficial as it helps trigger physiological modifications and stimulates stress-induced protective responses in plants. This process occurs not only in plant cells but also in human cells and can activate bioactive compounds responsible for defensive strategies which prevent the oxidation that causes neurodegenerative heart diseases and some types of cancer. The aim of this study was to illustrate the numerous elicitors derived from different origins which have the capacity to enhance phenolic and antioxidant properties in plants and shed some light on the importance of plant metabolomics as a powerful tool for finding information on detecting secondary metabolites, which may help in natural product discovery. This paper reviews the literature related to the elicitation process and metabolomics, providing an evaluation of their results. Elicitation, in both biotic and abiotic forms, is an innovative tool which activates and improves the production of secondary metabolites in plants, though some metabolites behave differently. On the other hand, metabolomics provides insight into how these metabolites respond to elicitation. Full exploration of these elicitors in plants will provide academic and scientific information to support the development of functional and nutraceutical foods with the potential to improve the quality of human health and alleviate the effects of prevalent non-communicable diseases.
PDF Abstract XML References Citation
Received: December 15, 2017;
Accepted: June 20, 2018;
Published: August 15, 2018
Copyright: © 2018. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Iyiola Oluwakemi Owolabi, Chutha Takahashi Yupanqui and Sunisa Siripongvutikorn, 2018. Enhancing Secondary Metabolites (Emphasis on Phenolics and Antioxidants) in Plants through Elicitation and Metabolomics. Pakistan Journal of Nutrition, 17: 411-420.
DOI: 10.3923/pjn.2018.411.420
URL: https://scialert.net/abstract/?doi=pjn.2018.411.420
DOI: 10.3923/pjn.2018.411.420
URL: https://scialert.net/abstract/?doi=pjn.2018.411.420
INTRODUCTION
It has been generally established that when plants are treated with certain living (biotic) or non-living (abiotic) factors called elicitors, they activate component function within cells by synchronously changing primary and secondary metabolic processes1. Not only do plants generate different secondary metabolites to respond to environmental changes but certain elicitors also improve the production of metabolites and/or signal transduction2. A synergy between the stimulators/elicitors and the metabolites produced help the stimulated plant to overcome biological and environmental stresses. Several plant elicitors have been found to stimulate defence responses, while others stimulate growth responses, leading to increase morpho-physiological and yield parameters3.
Many studies on this topic have reported the benefits of consuming food with an abundance of phenolics, which are important to human health4. As reviewed by Randhir et al.5, phenolics are secondary metabolites which are known to be partially generated in response to the interaction between plants and the environment. These components have garnered attention due to their established capacity to mitigate the risks of certain ailments and to their therapeutic properties against tumours, viruses, microbes, inflammation, hypotension and oxidants6.
Improving the nutritional and nutraceutical value of plants will be greatly beneficial to human health. This can be attained in two major ways: (1) Genetic manipulation and (2) Modification of metabolism, by elicitors7. There have, however been various controversies in relation to the genetic engineering of plants, whereas modification of the chemical composition and selected bioactivities of plant foods by elicitors is less costly and more socially acceptable worldwide8. Thus, elicitation of seeds and sprouts appears to be a promising and innovative alternative to other biological techniques used to enhance the yields of plant metabolites and nutraceuticals in food4.
Both biotic and abiotic forms of elicitors exist that stimulate metabolism and there have been many studies of their potential use in improving seed germination. Recently, elicitors have been intensively studied due to their ability to enhance phenolics and antioxidants in plants. This paper will review several types of elicitation studies that examine a wide range of food plants with important dietary roles.
ELICITATION
Elicitation is a process of increasing the production of secondary metabolites in plants to improve their survival and competitiveness9, while elicitors are components that stimulate any type of physiological response in plants10. The active role of secondary metabolites in the adaptations that plants make in the face of environmental stress cannot be overemphasized and elicitors are known to trigger biochemical systems in plants which increase the production of secondary metabolites11. Elicitation could thus be a low-risk strategy to increase the presence of these compounds in plants12. Based on their source, these elicitors can be categorized into two groups; biotic and abiotic (Fig. 1).
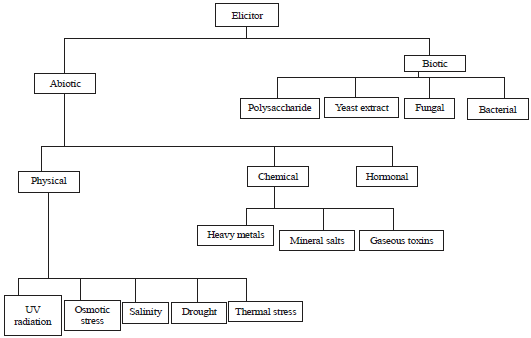 | |
| Fig. 1: | Classification of elicitors based on their origin |
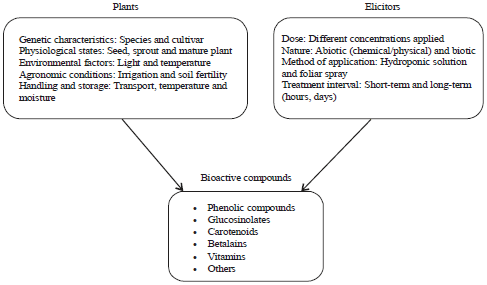 | |
| Fig. 2: | Factors influencing plant responses to bioactive compounds |
Secondary metabolites produced during elicitation depend upon the receptor (plant) itself and the type of elicitor involved (Fig. 2). The receptor perceives the elicitor, which leads to a variety of responses, including reversible phosphorylation and dephosphorylation of plasma membrane proteins and cytosolic proteins, cytosolic [Ca2+] enhancement, Cl‾ and K+ efflux/H+ influx, extra cellular alkalinisation and cytoplasmic acidification, mitogen-activated protein kinase (MAPK) activation, NADPH oxidase activation and the production of reactive oxygen and nitrogen species (ROS and RNS), early defence response gene expression, jasmonate production, late defence response gene expression and secondary metabolite accumulation (Fig. 3).
Some plant hormones have also been defined as elicitors because of their capacity to induce the genetic expression of various photosynthetic pathways. This review focused particularly on studies of elicitation in plants that aimed at enhancing their nutrition with the specific aim of stimulating the production of phenolics and antioxidants. The review investigated the use of both living and non-living (i.e., biotic and abiotic) elicitors.
Abiotic elicitation: Abiotic elicitation involves triggering the synthesis of phytochemical compounds in plants using chemical or physical stimuli. Abiotic elicitors are derived from non-living sources that can be categorized into i) Physical factors and ii) Compounds9. These elicitors have been tested individually and in combination, in water culture (hydroponics) or sprays and at different growth stages or even after harvesting13. This section reviews studies on the effects of abiotic elicitors on the bioactivities of different plant species.
Inorganic salts: Some antioxidant compounds have been enhanced by halo-elicitation in several plant species with NaCl, NaNO3, MnSO4, MgCl2, K3PO4 and KNO3 salts being the most commonly used to boost phytochemical and antioxidant properties14. For example, Daiponmak et al.15 reported that the total phenolic content, antioxidant capacity and cyanidin-3-glucoside content all significantly increased in the leaves of Thai rice cultivars grown for 16 days and subjected to 11 days at a salinity of 60 nM NaCl. Furthermore, positive end products of osmopriming were correlated with the recovery and upsurge of nucleic acids, improved amalgamation of proteins and the repair of membranes16.
In addition, enzymes responsible for antioxidation such as ascorbate peroxidase (APX) and catalase (CAT), which play a key role in scavenging reactive oxygen species (ROS), have been reported to increase in rice grown under saline conditions17. Some other heavy metal salts such as AgNO3 or CdCl2 have also been found to frequently activate the synthesis of phytoalexin as in the case of Brugmansia candida (angel’s trumpet) where overproduction of two tropane alkaloids, scopolamine and hyoscyamine, was elicited18. Additionally, the rare-earth metal lanthanum significantly enhanced the production of paclitaxel (Taxol) (280%×normal yield) in an in vivo experiment involving Taxus spp.19. Paclitaxel is a diterpenoid compound found in the bark of Taxus trees and is recognized by the Food and Drug Administration (FDA) to play a role in curing ovarian and breast cancer20.
Sound waves: Sound wave technology has been applied to different plants and plant development has been successfully controlled by sound waves with various frequencies, pressures, times of exposure and distances from the source21.
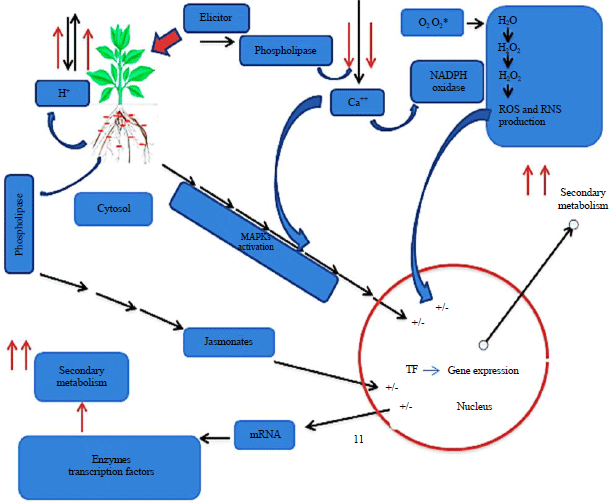 | |
| Fig. 3: | Diagram representing the potential responses of plant cells to elicitation. R: Receptor, PL: Phospholipase, MAPKs: Mitogen activated protein kinases, ROS: Reactive oxygen species, RNS: Reactive nitrogen species, TF: Transcription factors |
The lowest frequency class in the acoustic spectrum is infrasound, which has been used in various diagnostic and therapeutic applications. Acoustic waves at frequencies greater than 20 kHz are known as ultrasound and this frequency has been widely used in medical practice for more than 50 years as both a diagnostic and a therapeutic tool. Recently, its potential for food production has attracted research interest. These sounds can interact with biological tissues through thermal and mechanical processes22-24. The effects of sound waves have been studied on several plants. For example, a study of Actinidia chinensis (kiwi fruit) revealed that root activity was stimulated and moreover, the total length and root number increased. In addition, the capacity of soluble proteins together with superoxide dismutase (SOD) activity in the plant increased when it was exposed to sound waves with a frequency of 1 kHz at 100 dB. However, those indexes decreased when the sound waves exceeded 1 kHz at 100 dB25,26. Other studies have also been carried out on chrysanthemum (Dendranthema morifolium) calluses27, chlorella (Chlorella pyrenoidosa)28,29, onion (Allium cepa L.)30 and sea grass (Thalassia testudinum)31.
Drought: Previous studies of the effect of drought on plants have revealed its effects on antioxidant enzyme activities and oxidative stress. Two genotypes of wheat (Triticum aestivum L.), namely, Heshanhtou (HST, drought resistant) and Longchun 15 (LC15, drought susceptible), subjected to osmotic stress for 2 days demonstrated a rise in lipid peroxidation SOD and CAT activities32. Another study on barley (Hordeum vulgare L.) showed that SOD activity was improved in a drought-tolerant genotype subjected to increasing drought for 12 days33. The results of a study on Populus cathayana showed that watering at 25, 40, 40, 40 and 25% of field capacity amplified the actions of CAT, SOD, POD, APX and glutathione reductase (GR), with increased values of 3.7, 71.4, 77.8, 92.8 and 29%, respectively, compared with those at 100% field capacity. In contrast, 40, 75, 75, 75 and 55% gave values of 5.6, 112.3, 130.4, 115.4 and 55% of those at 100% field capacity, respectively, when evaluated in Populus kangdingensis34. To further affirm the findings from the previous studies, water stress-resistant and water stress-susceptible genotypes of Kentucky bluegrass (Poa pratensis) were exposed to progressive drought for 18 and 22 days and the APX gene expression was observed to increase35.
Brine mineral water: Brine mineral water (BMW) generally refers to seawater at depths equal to those at which photosynthesis occurs36. This water contains an abundance of minerals such as Ca, Sr, Mn, Zn, Fe, Cu, Ni, V and Se and is more similar to human body fluids than is deep sea water (DSW)37. Moon et al.38 observed that in Rehmannia glutinosa plants treated with BMW, the RgAPX gene was stimulated with 5% BMW after 48 h and PAL gene transcription advanced appreciably at 5% BMW after both 1 and 48 h. Additionally, the gene expression responsible for rbcL was higher when treated with BMW compared with control plants.
Osmo-elicitation: This is a process in which osmotic solutions having low water potential are used in improving secondary metabolite production in plants. The most commonly used substances are inorganic salts, polyethylene glycol (PEG), mannitol and glycerol15. A study on rice seedlings subjected to mild and severe osmotic stress using PEG (6000) at -0.5 and -2.0 Mpa for 24 h revealed that SOD and APX expression increased consistently with increases in the severity of water stress, but CAT declined. Sharma and Durbey39 reported that osmo-elicitation enhanced the activation and synthesis of some enzymes that catalyze the catabolic reaction and mobilization of stored substances. These occurrences have been addressed in various species in which there was an increase in CAT expression, most notably in Arabidopsis40 and sunflower (Helianthus annuus L.)41 and in CAT activity, such as in maize seeds42. In sunflower seeds elicited by PEG (6000) at -2 Mpa, there was an increase in SOD, CAT and GR activity as a result of metabolic actions43,44. Higher SOD and POD activity were recorded when rye seeds were treated with 30% PEG (6000) for 24 h45.
Hormonal elicitation: This process involves subjecting seeds or plant materials to various hormones, such as salicylic acid, ascorbate, kinetin, etc., which help to promote the development and improvement of seedlings. There are various plant hormones which act as elicitors. The most commonly used plant hormones are salicylic acid and jasmonic acid, which are important signals for the expressions of genes. Salicylic acid is useful in regulating resistance to some fungal, viral and bacterial pathogens, whereas jasmonic acid regulates protein synthesis via the octadecanoid pathway9. In the report by Wang and Zheng46, treating young red and black varieties of raspberry fruits with 0.01 mM methyl jasmonate (MeJA) led to an increase in their levels of anthocyanins and phenolic compounds.
Biotic elicitation: Biotic elicitors are compounds generated within living organisms, either from pathogens or the plant itself 9. The earliest biotic elicitors were discovered in the early 1970s and since then, there has been a great deal of evidence for compounds derived from pathogens which induce resistance feedback in both whole plants and cultures from plant cells. Examples include some compounds among the lipo- and glycoproteins or oligosaccharides. Albersheim et al.47 were the first to isolate oligosaccharides that activated a range of genes responsible for defence. The following sections provide examples of biotic elicitors and their role in the production of secondary metabolites in plants.
Elicitation with fish protein hydrolysates (FPH), lactoferrin (LF) and oregano extract (OE): Randhir et al.48 and Shetty49 reported that elicitation of fava bean (Vicia faba) seeds with fish protein hydrolysates (FPH), lactoferrin (LF) and oregano extract (OE) enhanced the production of phenolic antioxidants with the ability to quench free radicals. Fava bean seedlings are abundant in levo dihydroxy phenylalanine (L-DOPA), the precursor of dopamine that is used in the treatment of Parkinson’s disease. These elicitors were found to stimulate the phenylpropanoid pathway through the pentose phosphate and shikimate pathway and enhance the synthesis of phenolics. The L-DOPA content present in fava beans demonstrated a 40% increase following elicitation by LF, while elicitation with FPH and OE resulted in a 20% increase compared to an untreated control. The highest stimulation in the L-DOPA content was observed on day 2 for fava beans treated with 2 m L–1 of FPH, which recorded a value 100% higher than that of the control seedlings.
Ulvan (a sulphated polysaccharide sourced from the green alga Ulva fasciata): Ulvan, combined with exogenous elicitors extracted from the cell walls of fungi e.g., chitin fragment50, oligomers and polymers of chitosan51 or oligoglucans52, is known to enhance antioxidant activities in plants. According to study carried out by Paulert et al.53, pre-treating wheat cells with ulvan and chitin led to a five- to six-fold increase in oxidative bursts, whereas ulvan and chitosan only doubled the response. The H2O2 synthesis elicited by chitin and chitosan increased by 150 and 80 times, respectively. Another advantage of pre-treating entire plants with ulvan alone was an appreciable decrease in the severity of the symptoms of Blumeria graminis in wheat (by 45%) and in barley (by 80%).
Plant growth promoting rhizobacteria (PGPR): Many studies have shown that fungi or pathogenic bacteria or other natural elicitors can trigger the production of secondary metabolites with pharmacological activity54-57. Manero et al.58 reported that among the terpenes, cardenolides are known to be present in highly productive varieties of Digitalis lanata growing under controlled conditions. These cardenolides were increased by strains from the rhizosphere of uncultivated populations of Digitalis. Nine PGPR strains were tested on soybean seedlings and resulted in differential alterations of isoflavones (IF) with four behaviours based on changes in daidzein and genistein IF families59. Additionally, six rhizobacterial strains were tested on Hypericum perforatum seedlings, leading to a statistically significant increase in hypericin and a moderate as well as significant increase in pseudohypericin60.
Arachidonic acid: Arachidonic acid (AA; 5, 8, 11, 14-cis-eicosatetraenoic acid) is identified with the omega-6-set of polyunsaturated fatty acids that cannot be synthesized by animals and humans but are vital to their health and thus should be added to their diet. Dedyukhina et al.61 evaluated the role of AA as an elicitor on potato, tomato, sugar beet and vine plants and showed that it was able to increase the plants’ resistance to diseases. However, the study found that high concentrations of AA (over 10–5 M) induced necrosis of plant tissues and the accumulation of antimicrobial compounds (phytoalexins), whereas low AA concentrations (10–7-10–7 M) elicited systemic and prolonged resistance to phytopathogenic infections in a way that is analogous to that of the immunization process.
Yeast: In the report of Mu et al.62, three-months-old Lycoris chinensis sprouts were elicited by a yeast elicitor (YE) at 0.15 g L–1, which enhanced the galantamine (GAL) content by approximately 1.62 times over the control. Additionally, the maximum level of lycorine, which was 1.38 times higher than that observed in the control, was attained in the sprouts on the 10th day when elicited by 0.01 g L–1 YE.
Saccharomyces cerevisiae (SC) and Salix daphnoides bark (SD) extracts: Recently, metabolomics has been gaining attention; it has not only been utilized by plant breeders to improve plant yield and quality but also been used in functional foods and nutraceuticals to analyze and quantify the secondary metabolites in plant systems. Information derived from metabolomics may help to provide insights into how secondary metabolite-producing systems react to chemical and biological stimuli. This in turn can offer a medium for investigating the biological mechanism of natural products, newly isolated antibiotics and chemotherapeutics as a result of the metabolomic changes generated within treated organisms63.
Secondary metabolites and metabolomics: Metabolomics can be defined as a study that entails all aspects of small molecules contained in a biological system and gives a precise measurement of the secondary metabolite production detected within a selected organism63. Secondary metabolites are regarded as small biomolecules that are non-essential for the life of the organism producing them64.
These metabolites provide several types of survival benefits to the organism producing them. For example, metabolites can act as a metabolic defence mechanism (e.g., plant flavonoids and alkaloid toxins), protecting plants against environmental stress, e.g., pigments and osmoprotectants. Several secondary metabolites have been of huge significance to humans as some are used over an extended period as active drug ingredients in medicine. For example, many antibiotics, antitumor agents and antivirals are derived from secondary metabolites, as are antipyretics such as aspirin, hallucinogens such as lysergic acid diethylamide (LSD), cholesterol-lowering drugs such as lovastatin65,66, herbicides and phytotoxins used in agriculture67 and food additives (colouring, flavouring and sweetening agents)68.
However, secondary metabolites are regarded as biosynthetic end-products that, contrary to primary metabolites, are capable of accumulating at higher levels than the fluxes shown in central metabolism. Therefore, researchers may say that relatively abundant secondary metabolites are suitable for comparative metabolomics workflows64.
Methodologies and instrumentations of plant metabolomics seem to be developing rapidly, though precise and exhaustive analysis of the whole metabolome of a biological system appears currently impossible69. There have been extensive analyses of mixtures that are extremely complex and are enhanced through a series of integrated technologies and methodologies, for example, non-destructive NMR (Nuclear Magnetic Resonance spectroscopy) and Mass Spectrometry (MS)-based methods including GC-MS (Gas Chromatography-MS), LC-MS (Liquid Chromatography-MS) and CE-MS (Capillary Electrophoresis- MS) and FI-ICR-MS (Fourier Transform Ion Cyclotron Resonance-MS)70,71. These methods have proven their potential utility in plant metabolomic studies in several plant species, including tomato, rice, wheat and maize72,73.
Nonetheless, there appears to be an indelible limitation of each of the analytical platforms and as a result, combining approaches in metabolomic analysis is gradually gaining attention.
Further studies in metabolomics should focus more on the following: (1) Improving the metabolomics platform in order to facilitate the precise, effective identification and quantification of various metabolites (emphasis on secondary metabolites) and accurate interpretation of data generated from rapid incorporation with other omics platforms and (2) All-encompassing probing into molecular and biochemical mechanisms of metabolomic differences in plants by using both non-targeted and targeted approaches. These recommendations will help expand and enrich the understanding of plant metabolism for better crop yield, quality and health benefits.
Looking beyond: Over the last decade, interest in functional foods has been rising as greater numbers of people become more anxious about their nutrition and diets. The food industry continues to seek new and exceptional ingredients, thereby, creating an increased demand for functional food compounds. It is, therefore, a matter of the utmost priority to identify naturally occurring compounds or collections of them and to investigate claims for their therapeutic properties. Thus, the use of both biotic and abiotic elicitors should be regarded as a strategy complementing breeding programmes, or even as an alternative to employing genetic modifications. Elicitation stress induced in a short-term, controlled, pre or post-harvest setting presents a promising tool which can be applied by the fresh produce industry to boost the nutraceutical contents of their products. However, in the production of new or improved plants, functional property approval are vital to ensure the safety of substances introduced into food materials for human consumption. Although biotic and abiotic elicitors may provide new prospects for averting certain diseases, monitoring their effects in clinical trials is therefore essential for the health and safety of consumers.
CONCLUSION
Elicitation, whether biotic or abiotic, applied alone or in combination with other approaches, is a strategy which offers many potential benefits through enhancing phytochemicals in foods. When analyzed and quantified through plant metabolomics, the products and ingredients produced through elicitation could lead to the development of functional foods or nutraceuticals, potentially providing numerous benefits to improving basic nutrition as well as health benefits.
SIGNIFICANCE STATEMENT
This study examines the economically affordable and natural elicitation process of plants as well as the importance of metabolomics in detecting and quantifying metabolites that are potentially beneficial for improving plant quality and human health. This study will help researchers to uncover critical areas of plant metabolomics that researchers have not yet been able to explore. Thus, new theories on both plant elicitation and metabolomics may be proposed.
ACKNOWLEDGMENT
The support of the Higher Education Research Promotion and Thailand’s Education Hub for the Southern Region of Asian Countries Project office of the Higher Education Commission in granting a scholarship to pursue post-graduate studies is acknowledged. We also acknowledge the graduate school of Prince of Songkla University, Thailand for awarding the ‘Graduate school Dissertation Funding’.
REFERENCES
- Jeandet, P., 2015. Phytoalexins: Current progress and future prospects. Molecules, 20: 2770-2774.
CrossRefDirect Link - Swieca, M., 2015. Elicitation with abiotic stresses improves pro-health constituents, antioxidant potential and nutritional quality of lentil sprouts. Saudi J. Biol. Sci., 22: 409-416.
CrossRefDirect Link - Randhir, R., P. Shetty and K. Shetty, 2002. L-DOPA and total phenolic stimulation in dark germinated fava bean in response to peptide and phytochemical elicitors. Process Biochem., 37: 1247-1256.
CrossRefDirect Link - Kelly, W.B., J.E. Esser and J.I. Schroeder, 1995. Effects of cytosolic calcium and limited, possible dual, effects of G protein modulators on guard cell inward potassium channels. Plant J., 8: 479-489.
CrossRefDirect Link - Gawlik-Dziki, U., M. Swieca, D. Dziki and D. Sugier, 2013. Improvement of nutraceutical value of broccoli sprouts by natural elicitors. Acta Scient. Polonorum Hortorum Cultus, 12: 129-140.
Direct Link - Gawlik-Dziki, U., M. Jezyna, M. Swieca, D. Dziki, B. Baraniak and J. Czyz, 2012. Effect of bioaccessibility of phenolic compounds on in vitro anticancer activity of broccoli sprouts. Food Res. Int., 49: 469-476.
CrossRefDirect Link - Patel, H. and R. Krishnamurthy, 2013. Elicitors in plant tissue culture. J. Pharmacogn. Phytochem., 2: 2278-4136.
Direct Link - Radman, R., T. Saez, C. Bucke and T. Keshavarz, 2003. Elicitation of plants and microbial cell systems. Biotechnol. Applied Biochem., 37: 91-102.
CrossRefPubMedDirect Link - Zhao, J., L.C. Davis and R. Verpoorte, 2005. Elicitor signal transduction leading to production of plant secondary metabolites. Biotechnol. Adv., 23: 283-333.
CrossRefPubMedDirect Link - Garcia-Mier, L., S.N. Jimenez-Garcia, R.G. Guevara-Gonzalez, A.A. Feregrino-Perez, L.M. Contreras-Medina and I. Torres-Pacheco, 2015. Elicitor mixtures significantly increase bioactive compounds, antioxidant activity and quality parameters in sweet bell pepper. J. Chem., Vol. 2015.
CrossRefDirect Link - Baenas, N., C. Garcia-Viguera and D.A. Moreno, 2014. Elicitation: A tool for enriching the bioactive composition of foods. Molecules, 19: 13541-13563.
CrossRefDirect Link - Tiryaki, I. and Y. Buyukcingil, 2009. Seed priming combined with plant hormones: Influence on germination and seedling emergence of sorghum at low temperature. Seed Sci. Technol., 37: 303-315.
CrossRefDirect Link - Daiponmak, W., P. Theerakulpisut, P. Thanonkao, A. Vanavichit and P. Prathepha, 2010. Changes of anthocyanin cyanidin-3-glucoside content and antioxidant activity in Thai rice varieties under salinity stress. ScienceAsia, 36: 286-291.
CrossRefDirect Link - Umnajkitikorn, K., B. Faiyue and K. Saengil, 2013. Enhancing antioxidant properties of germinated Thai rice (Oryza sativa L.) cv. Kum Doi Saket with salinity. J. Rice Res., Vol. 1.
Direct Link - Stintzi, A., H. Weber, P. Reymond, J. Browse and E.E. Farmer, 2001. Plant defense in the absence of jasmonic acid: The role of cyclopentenones. Proc. Natl. Acad. Sci. USA., 98: 12837-12842.
CrossRefDirect Link - Wu, L., S.W. Hallgren, D.M. Ferris and K.E. Conway, 2001. Effects of moist chilling and solid matrix priming on germination of loblolly pine (Pinus taeda L.) seeds. New For., 21: 1-16.
CrossRefDirect Link - Kovacs, P., G. Csaba, E. Pallinger and R. Czaker, 2007. Effects of taxol treatment on the microtubular system and mitochondria of Tetrahymena. Cell Biol. Int., 31: 724-732.
CrossRefDirect Link - Hassanien, R.H.E., T.Z. Hou, Y.F. Li and B.M. Li, 2014. Advances in effects of sound waves on plants. J. Integr. Agric., 13: 335-348.
CrossRefDirect Link - O'Brien, Jr. W.D., 2007. Ultrasound-biophysics mechanisms. Prog. Biophys. Mol. Biol., 93: 212-255.
CrossRefDirect Link - Whittingham, T.A., 2007. Medical diagnostic applications and sources. Prog. Biophys. Mol. Biol., 93: 84-110.
CrossRefDirect Link - Rokhina, E.V., P. Lens and J. Virkutyte, 2009. Low-frequency ultrasound in biotechnology: State of the art. Trends Biotechnol., 27: 298-306.
CrossRefDirect Link - Yang, X.C., B.C. Wang and C.R. Duan, 2003. Effects of sound stimulation on energy metabolism of Actinidia chinensis callus. Colloids Surf. B: Biointerfaces, 30: 67-72.
CrossRefDirect Link - Zhao, H.C., J. Wu, L. Zheng, T. Zhu and B.S. Xi et al., 2003. Effect of sound stimulation on Dendranthema morifolium callus growth. Colloids Surf. B Biointerfaces, 29: 143-147.
CrossRefDirect Link - Jiang, S., J. Huang, X. Han and X. Zeng, 2011. Influence of audio frequency mixing of music and cricket voice on growth of edible mushrooms. Trans. Chin. Soc. Agric. Eng., 27: 300-305.
Direct Link - Jiang, S., H. Rao, Z. Chen, M. Liang and L. Li, 2012. Effects of sonic waves at different frequencies on propagation of Chlorella pyrenoidosa. Agric. Sci. Technol., 13: 2197-2201.
Direct Link - Ekici, N., F. Dane, L. Mamedova, I. Metin and M. Huseyinov, 2007. The effects of different musical elements on root growth and mitosis in onion (Allium cepa) root apical meristem (Musical and biological experimental study). Asian J. Plant Sci., 6: 369-373.
CrossRefDirect Link - Wilson, P.S. and K.H. Dunton, 2009. Laboratory investigation of the acoustic response of seagrass tissue in the frequency band 0.5-2.5 kHz. J. Acoust. Soc. Am., 125: 1951-1959.
CrossRefDirect Link - Acar, D., L. Turkan and F. Ozdemir, 2001. Superoxide dismutase and peroxidase activities in drought sensitive and resistant barley (Hordeum vulgare L.) varieties. Acta Physiol. Planta., 3: 351-356.
CrossRefDirect Link - Yang, F., X. Xu, X. Xiao and C. Li, 2009. Responses to drought stress in two poplar species originating from different altitudes. Biol. Planta., 53: 511-516.
CrossRefDirect Link - Xu, L., L. Han and B. Huang, 2011. Antioxidant enzyme activities and gene expression patterns in leaves of Kentucky bluegrass in response to drought and post-drought recovery. J. Am. Soc. Hortic. Sci., 136: 247-255.
Direct Link - Kim, W.J., H. Li, T.J. Yoon, J.M. Sim, S.K. Choi and K.H. Lee, 2009. Inhibitory activity of brine mineral water on cancer cell growth, metastasis and angiogenesis. Korean J. Food Nutr., 22: 542-547.
Direct Link - Moon, D.S., D.H. Jung, H.J. Kim and P.K. Shin, 2004. Comparative analysis on resources characteristics of deep ocean water and brine groundwater. J. Korean Soc. Mar. Environ. Energy, 7: 42-46.
Direct Link - Sharma, P. and R.S. Dubey, 2005. Drought induces oxidative stress and enhances the activities of antioxidant enzymes in growing rice seedlings. Plant Growth Regul., 46: 209-221.
CrossRefDirect Link - Gallardo, K., C. Job, S.P.C. Groot, M. Puype, H. Demol, J. Vanderkerckhove and D. Job, 2001. Proteomic analysis of Arabidopsis seed germination and priming. Plant Physiol., 126: 835-848.
CrossRefDirect Link - Kibinza, S., J. Bazin, C. Bailly, J.M. Farrant, F. Corbineau and H. El-Maarouf-Bouteau, 2011. Catalase is a key enzyme in seed recovery from ageing during priming. Plant Sci., 181: 309-315.
CrossRefDirect Link - Chiu, K.Y., C.L. Chen and J.M. Sung, 2002. Effect of priming temperature on storability of primed sh-2 sweet corn seed. Crop Sci., 42: 1996-2003.
Direct Link - Bailly, C., A. Benamar, F. Corbineau and D. Come, 1998. Free radical scavenging as affected by accelerated ageing and subsequent priming in sunflower seeds. Physiol. Planta., 104: 646-652.
CrossRefDirect Link - Bailly, C., A. Benamar, F. Corbineau and D. Come, 2000. Antioxidant systems in sunflower (Helianthus annuus L.) seeds as affected by priming. Seed Sci. Res., 10: 35-42.
CrossRefDirect Link - Liu, J., G.S. Liu, D.M. Qi, F.F. Li and E.H. Wang, 2002. Effect of PEG on germination and active oxygen metabolism in wildrye (Leymus chinensis) seeds. Acta Pratacult. Sin., 11: 59-64.
Direct Link - Wang, S.Y. and W. Zheng, 2005. Preharvest application of methyl jasmonate increases fruit quality and antioxidant capacity in raspberries. Int. J. Food Sci. Technol., 40: 187-195.
CrossRefDirect Link - Albersheim, P., A.R. Ayers Jr., B.S. Valent, J. Ebel, M. Hahn, J. Wolpert and R. Carlson, 1977. Plants interact with microbial polysaccharides. J. Cell. Biochem., 6: 599-616.
CrossRefDirect Link - Randhir, R., Y.T. Lin and K. Shetty, 2004. Stimulation of phenolics, antioxidant and antimicrobial activities in dark germinated mung bean sprouts in response to peptide and phytochemical elicitors. Process Biochem., 39: 637-646.
CrossRefDirect Link - Shetty, K., 2004. Role of proline-linked pentose phosphate pathway in biosynthesis of plant phenolics for functional food and environmental applications: A review. Process Biochem., 39: 789-804.
CrossRefDirect Link - Ortmann, I. and B.M. Moerschbacher, 2006. Spent growth medium of Pantoea agglomerans primes wheat suspension cells for augmented accumulation of hydrogen peroxide and enhanced peroxidase activity upon elicitation. Planta, 224: 963-970.
CrossRefDirect Link - Dos Santos, A.L.W., N.E. El Gueddari, S. Trombotto and B.M. Moerschbacher, 2008. Partially acetylated chitosan oligo-and polymers induce an oxidative burst in suspension cultured cells of the gymnosperm Araucaria angustifolia. Biomacromolecules, 9: 3411-3415.
CrossRefDirect Link - Shinya, T., R. Menard, I. Kozone, H. Matsuoka and N. Shibuya et al., 2006. Novel β-1,3-, 1,6-oligoglucan elicitor from Alternaria alternate 102 for defense responses in tobacco. FEBS J., 273: 2421-2431.
CrossRefDirect Link - Paulert, R., D. Ebbinghaus, C. Urlass and B.M. Moerschbacher, 2010. Priming of the oxidative burst in rice and wheat cell cultures by ulvan, a polysaccharide from green macroalgae and enhanced resistance against powdery mildew in wheat and barley plants. Plant Pathol., 59: 634-642.
CrossRefDirect Link - Cai, Z., A. Kastell, I. Mewis, D. Knorr and I. Smetanska, 2012. Polysaccharide elicitors enhance anthocyanin and phenolic acid accumulation in cell suspension cultures of Vitis vinifera. Plant Cell Tissue Organ Cult., 108: 401-409.
CrossRefDirect Link - Korsangruang, S., N. Soonthornchareonnon, Y. Chintapakorn, P. Saralamp and S. Prathanturarug, 2010. Effects of abiotic and biotic elicitors on growth and isoflavonoid accumulation in Pueraria candollei var. candollei and P. candollei var. mirifica cell suspension cultures. Plant Cell. Tissue Organ Cult., 3: 333-342.
CrossRefDirect Link - Coste, A., V. Laurian, A. Halmagyi, C. Deliu and G. Coldea, 2011. Effects of plant growth regulators and elicitors on production of secondary metabolites in shoot cultures of Hypericum hirsutum and Hypericum maculatum. Plant Cell Tissue Organ Cult., 106: 279-288.
CrossRefDirect Link - Pawar, K.D., A.V. Yadav, Y.S. Shouche and S.R. Thengane, 2011. Influence of endophytic fungal elicitation on production of inophyllum in suspension cultures of Calophyllum inophyllum L. Plant Cell Tissue Organ Cult., 106: 345-352.
CrossRefDirect Link - Manero, F.J.G., B. Ramos, J.A.L. Garcia, A. Probanza and M.L.B. Casero, 2003. Systemic induction of the biosynthesis of terpenic compounds in Digitalis lanata. J. Plant Physiol., 160: 105-113.
CrossRefPubMedDirect Link - Ramos-Solano, B., E. Algar, A. Garcia-Villaraco, J. Garcia-Cristobal, J.A.L. García and F.J. Gutierrez-Manero, 2010. Biotic elicitation of isoflavone metabolism with plant growth promoting rhizobacteria in early stages of development in Glycine max var. Osumi. J. Agric. Food Chem., 58: 1484-1492.
CrossRefDirect Link - Gutierrez-Manero, F.J., A. Garcia-Villaraco, J.A. Lucas, E. Gutierrez and B. Ramos-Solano, 2015. Inoculant/elicitation technology to improve bioactive/phytoalexin. Int. J. Curr. Microbiol. Applied Sci., 4: 224-241.
Direct Link - Dedyukhina, E.G., S.V. Kamzolova and M.B. Vainshtein, 2014. Arachidonic acid as an elicitor of the plant defense response to phytopathogens. Chem. Biol. Technol. Agric., Vol. 1, No. 1.
CrossRefDirect Link - Mu, H.M., R. Wang, X.D. Li, Y.M. Jiang and C.Y. Wang et al., 2009. Effect of abiotic and biotic elicitors on growth and alkaloid accumulation of Lycoris chinensis seedlings. Zeitschrift Naturforschung C, 64: 541-550.
CrossRefDirect Link - Covington, B.C., J.A. McLean and B.O. Bachmann, 2017. Comparative mass spectrometry-based metabolomics strategies for the investigation of microbial secondary metabolites. Natl. Prod. Rep., 34: 6-24.
CrossRefDirect Link - Mousa, W.K. and M.N. Raizada, 2013. The diversity of anti-microbial secondary metabolites produced by fungal endophytes: An interdisciplinary perspective. Front Microbiol., Vol. 4.
CrossRef - Khazir, J., B.A. Mir, S.A. Mir and D. Cowan, 2013. Natural products as lead compounds in drug discovery. J. Asian Nat. Prod. Res., 15: 764-788.
CrossRefDirect Link - Duke, S.O., F.E. Dayan, J.G. Romagni and A.N. Rimando, 2000. Natural products as sources of herbicides: Current status and future trends. Weed Res., 40: 99-111.
CrossRefDirect Link - Rao, S.R. and G.A. Ravishankar, 2002. Plant cell cultures: Chemical factories of secondary metabolites. Biotechnol. Adv., 20: 101-153.
CrossRefPubMedDirect Link - Hegeman, A.D., 2010. Plant metabolomics-meeting the analytical challenges of comprehensive metabolite analysis. Brief. Funct. Genom., 9: 139-148.
CrossRefDirect Link - Okazaki, Y. and K. Saito, 2012. Recent advances of metabolomics in plant biotechnology. Plant Biotechnol. Rep., 6: 1-15.
CrossRefDirect Link - Khakimov, B., S. Bak and S.B. Engelsen, 2014. High-throughput cereal metabolomics: Current analytical technologies, challenges and perspectives. J. Cereal Sci., 59: 393-418.
CrossRefDirect Link - Francki, M.G., S. Hayton, J.P.A. Gummer, C. Rawlinson and R.D. Trengove, 2016. Metabolomic profiling and genomic analysis of wheat aneuploid lines to identify genes controlling biochemical pathways in mature grain. Plant Biotechnol. J., 14: 649-660.
CrossRefDirect Link - Benard, C., S. Bernillon, B. Biais, S. Osorio and M. Maucourt et al., 2015. Metabolomic profiling in tomato reveals diel compositional changes in fruit affected by source-sink relationships. J. Exp. Bot., 66: 3391-3404.
CrossRefDirect Link