
Nur Kusmiyati
The Graduate School of Biotechnology, Universitas Gadjah Mada, Yogyakarta, Indonesia
Tutik Dwi Wahyuningsih
Department of Chemistry, Faculty of Mathematics and Natural Science, Universitas Gadjah Mada, Yogyakarta, Indonesia
Widodo
Faculty of Animal Science, Universitas Gadjah Mada, Yogyakarta, Indonesia
LiveDNA: 62.23055
Pakistan Journal of Nutrition
Year: 2018 | Volume: 17 | Issue: 8 | Page No.: 405-410







ABSTRACT
Background and Objective: A prebiotic is a non-digestible food component that supports the growth of probiotics in the gastrointestinal tract, thus providing beneficial effects to the host. One commercially available prebiotic is inulin, produced from Jerusalem artichokes and chicory. A previous study reported that the inulin from dahlia tubers is of better quality due to the presence of soluble and insoluble fibres. Due to its potential, inulin from dahlia tubers has been increasingly tested for its capability as a prebiotic. The aim of this study was to investigate the prebiotic effects of inulin and inulin extract from dahlia tubers on the viability of Lactobacillus casei AP and representative Enterobacteriaceae and to measure the prebiotic index and production of short-chain fatty acids. Methodology: This study focused on the combined effect of a prebiotic (inulin or inulin extract from dahlia tubers) and a probiotic, L. casei AP, which originated in a human gastrointestinal tract. This combination resulted in the growth suppression of pathogenic bacteria, the growth promotion of the probiotic and a high prebiotic index. The combination of L. casei AP and inulin/inulin extract, which was defined as a synbiotic, was also evaluated for the growth suppression of H. pylori and S. dysenteriae. Results: The L. casei AP had a higher growth rate in the presence of inulin extract or inulin than MRS did. Co-culturing E. coli and L. casei AP in media containing inulin extract suppressed the growth of E. coli and simultaneously resulted in the highest prebiotic index, at 4.70. The synbiotic effect of inulin extract together with L. casei AP was stronger in suppressing the growth of S. dysenteriae than of H. pylori. This inhibition was concomitant with the production of short-chain fatty acids, lactate, propionate, propionate and butyrate, which contributed to lowering the pH in the media. During fermentation, lactate was produced at the highest concentration (33.83-35.53%), followed by acetate (9.83-10.35%), propionate (0.05-0.45%) and butyrate (0.03-0.49%). Conclusion: Inulin extract from dahlia tubers acts very well as a prebiotic by supporting the growth of probiotic L. casei AP and suppressing the growth of the pathogens S. dysenteriae and H. pylori, resulting in a high prebiotic effect.
PDF Abstract XML References Citation
Received: March 06, 2018;
Accepted: May 24, 2018;
Published: July 15, 2018
Copyright: © 2018. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Nur Kusmiyati, Tutik Dwi Wahyuningsih and Widodo, 2018. Prebiotic Effect of Inulin Extract from Dahlia Tubers (Dahlia Pinnata L.) On the Growth Performance of Intestinal-origin Lactobacillus Casei AP. Pakistan Journal of Nutrition, 17: 405-410.
DOI: 10.3923/pjn.2018.405.410
URL: https://scialert.net/abstract/?doi=pjn.2018.405.410
DOI: 10.3923/pjn.2018.405.410
URL: https://scialert.net/abstract/?doi=pjn.2018.405.410
INTRODUCTION
Inulin is a storage carbohydrate that is found in more than 36.000 plant species1. Three main plant species [Jerusalem artichoke (Helianthus tuberosus L.), dahlia (Dahlia pinnata L.) and chicory (Cichorium intybus)] are currently employed to produce inulin. Inulin produced from Jerusalem artichoke and chicory is the main source for the inulin available on the market, whereas inulin produced from dahlia tubers has not yet been developed or made available on the market. A previous study reported that the inulin from dahlia tubers is of better quality due to the presence of soluble and insoluble fibres2. Kusmiyati et al.3 previously reported a selective method, ultrasonic- and microwave-assisted extraction (UMAE), that yielded high concentrations of inulin from dahlia tubers with a degree of polymerization of 19-20 and a molecular weight of 3.173-3.316.
Inulin fermentation in vivo contributes significantly to the increase of certain metabolites. This prebiotic can be selectively fermented by certain probiotics and other beneficial gastrointestinal microbes into short-chain fatty acids (SFCAs), mainly lactate, acetate, propionate and butyrate. The production of these metabolites eventually decreases the luminal pH and exerts an antagonistic activity on a large number of enteric pathogenic bacteria4 apart from competition for nutrients or for niche establishment5. In this study, the inulin extracted from dahlia tubers, together with commercial inulin, was assessed for its capacity to support the growth of probiotics and suppress pathogens and was measured for its prebiotic index (PI). Lactobacillus casei AP was used to measure PI due to its capability as a probiotic6. Widodo et al.6 reported that Lactobacillus casei AP, isolated from an Indonesian’s gastrointestinal tract, was capable of utilizing inulin as a carbon source. This present study investigated the capabilities of the probiotic L. casei AP and inulin extract from dahlia tubers to synergistically control the growth of S. dysenteriae ATCC 11835 and H. pylori ATCC 26695. The objectives of this study were to investigate the prebiotic effects of inulin and inulin extract from dahlia tubers and to measure the prebiotic index and production of short chain fatty acids.
MATERIAL AND METHODS
Bacterial cultures and media preparation: This study was a laboratory-based experiment carried out at the Research Centre for Biotechnology, Universitas Gadjah Mada, Yogyakarta, Indonesia. Bacterial cultures used in this study were Lactobacillus casei AP7, Enteropathogenic Escherichia coli (EPEC), Shigella dysenteriae ATCC 11835 and Heliobacter pylori ATCC 26695. The media used were inulin extract from dahlia tubers (Dahlia pinnata L.), inulin (Sigma-Aldrich, MO, USA), commercial de-Mann Rogosa Sharpe (MRS), synthetic de-Mann Rogosa Sharpe (glucosa-MRS), Cook Meat Medium (CMM) (Sigma-Aldrich, MO, USA), Plate Count Agar (PCA) (Oxoid, Hampshire, UK) and Trypticase Soy Broth/Agar (TSB/A) (Merck, Darmstadt, Germany).
Growth media for L. casei AP was MRS broth: The TSB was used for the propagation of H. pylori and S. dysenteriae. L. casei AP growth was microaerophilic/anaerobic (Oxoid, Hampshire, UK). The media used as the control and as the base for inulin media were sterilized by autoclaving at 121°C for 15 min. In the case of modified MRS broth, filter-sterilized L-cysteine hydrochloride was added to the autoclaved media. Filter-sterilized carbohydrates (inulin extract, inulin and glucose) were added to the modified MRS broth to final concentrations of 1.0% (w/v)8.
Bacterial growth measurement in inulin-containing media: To investigate the effects of inulin and inulin extract on the growth of L. casei AP, the bacteria were cultivated overnight in the appropriate selective media at 37°C. The activated cultures were harvested for 15 min with 2.500 rpm at 4°C using a Hettich centrifuge (Fisher Scientific, NH, USA). Then, the precipitate was washed twice with phosphate-buffered saline (0.1 mol L–1 phosphate buffer pH = 7.4 and 0.9% saline) and the final pellet was suspended in phosphate-buffered saline and diluted to approximately 106 cells mL–1. The ability of strains to utilize inulin as a carbon source was examined by growing them on media with inulin or inulin extract as the only carbon source, as a control, cells were grown in MRS. The growth of bacterial cells at 37°C was monitored for 24 h and measured using UV Spectrophotometry (Shimadzu, Kyoto, Japan) at an optical density (OD) of 620 nm. All measurements were repeated twice. The changes in pH were measured using an Inolab pH meter (Thomas Scientific, NJ, USA).
Prebiotic index (PI): The test was initiated by adding 1 mL inoculum separately into medium containing 1% inulin, 1% inulin extract or 1% sterile MRS. The media were then incubated at 37°C for 24 h. Total viable cells were measured by plating on specific media at 0, 12 and 24 h after bacterial growth using the pour plate method. For the enumeration of L. casei, the AP medium used was MRS agar and TSA was used for E. coli, the media were incubated for 48 h at 37°C. The Prebiotic Index was calculated using the formula described by Sanz et al.9. The PI was calculated using formula 1:
| (1) |
Short-chain fatty acid analysis: Short-chain fatty acid analysis was performed using the GC-MS chromatogram (Shimadzu, Kyoto, Japan) with a modified method by Conti e Silva et al.10. The column used was SP 1200, GP 10, SP-1200/1% H3PO4 on 80/100 chromosorb W AW (Sigma-Aldrich, MO, USA), with a column length of 1.5 m and a diameter of 3 mm. The injector temperature was 240°C, the detector temperature was 240°C, the column temperature was 140°C, the gas pressure was H2 = 0.6 kg cm–2 and the air pressure = 0.5 kg cm–2. The carrier gas was N2 gas at a pressure of N2 = 1.25 kg cm–2.
Growth inhibition of pathogenic strains: Growth inhibition of pathogenic strains in response to synbiotic products was evaluated by using the agar diffusion method11. Briefly, the pathogenic strains S. dysenteriae and H. pylori were inoculated on 5 mL of TSA at a final concentration of 107 cfu mL–1. Then, the synbiotic products were inserted into a well in the TSA. The plates were incubated in conditions suitable for each pathogen for 24-48 h. All of the petri dishes (Fisher Scientific, NH, USA) were duplexed. A clear zone around the wells was measured using calipers.
RESULT AND DISCUSSION
Effects of inulin extract on bacterial populations: In vitro experiments on three different growth media showed that both inulin (commercial standard) and the inulin extract from dahlia tubers supported the growth of L. casei AP (Fig. 1). The ability of L. casei AP to grow in inulin media was consistent with previous findings of Widodo et al.6, who demonstrated the ability of L. casei AP to utilize inulin as a carbon source. The growth pattern of L. casei AP in inulin extract was similar to that observed in inulin-containing and MRS media, in which all bacteria grew rapidly after approximately 6 h and reached the maximum growth at approximately 24 h. The optical density of the media containing inulin extract was significantly higher (p<0.05) than that of media containing inulin, which indicated that inulin extract was more effective at supporting the growth of L. casei AP than were inulin-containing or MRS media. Supplementation with inulin extract was found to have a significantly better effect on the viability of bacteria compared to media without inulin extract (p<0.05).
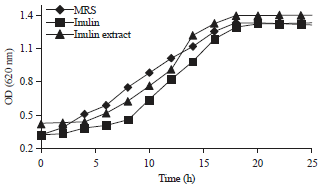 | |
| Fig. 1: | Growth of L. casei AP in three different media |
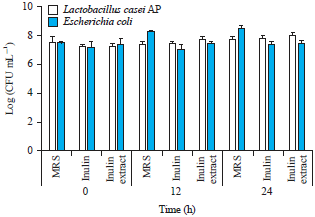 | |
| Fig. 2: | Bacterial populations at 0, 12 and 24 h in MRS, inulin and inulin extract as media |
Watson et al.12 demonstrated that lactulose, maltodextrin, fructooligosaccharides, galactooligosaccharides and inulin stimulate the growth performance of twelve different Lactobacillus species.
Co-culturing L. casei AP and E. coli in different growth media: To evaluate the effects of different media on the growth of L. casei AP and E. coli, both bacteria were co-cultured in various media (MRS, inulin, inulin extract) for 24 h. The growth of L. casei AP and E. coli was measured based on the enumeration of viable bacterial cells by means of total plate count (Fig. 2). After being grown in MRS as the growth medium for 12 and 24 h, E. coli had a faster growth rate than L. casei AP did. Meanwhile, in inulin- and inulin extract-containing media, E. coli had a slower growth rate than L. casei AP did (Fig. 2). Several studies have reported that fructooligosaccharides can support the growth of E. coli, Enterobacter and Salmonella13. In contrast, other studies have indicated that E. coli is unable to utilize fructooligosaccharides14. Van Laere et al.15 reported that arabino-oligosaccharides could support the growth of E. coli but that fructooligosaccharides could not do the same.
| Table 1: | Concentration of SCFA in media |
 | |
Different letters show significant differences, according to the BNT test (p<0.05) | |
Prebiotic index: The prebiotic indices (PIs) of inulin and inulin extracts were higher than that of MRS (Fig. 3). When grown in MRS media, the PI was 1.1. In contrast, when grown in inulin-containing and inulin extract media, the PI was 4.3 and 4.7, respectively (Fig. 3). The high prebiotic index indicated a larger probiotic population of L. casei AP than that of enterobacteriaceae/pathogen (E. coli) when co-cultured in the same growth medium, as seen in Fig. 2, suggesting that L. casei AP has a higher capability to consume inulin than E. coli does. Studies have reported differences in the PIs due to the differences in the prebiotics and probiotic strains used. It was reported that each bacterium has an unequal bacterial activity score. The PI value of dahlia tubers has not been reported previously but that of galactooligosaccharide was reported to be 0.1816. The PI score from static batch culture fermentation of FOS was 2.7817. In this study, inulin and inulin extracts were able to provide optimal carbon source requirements for the growth of L. casei AP by suppressing the growth of E. coli.
Synbiotic effect on the growth of S. dysenteriae and H. pylori: The data on the size of the inhibition zone when H. pylori and S. dysenteriae were grown in the presence of probiotic L. casei AP and prebiotic inulin or inulin extract are presented in Fig. 4. The synbiotic interaction between L. casei AP and inulin or inulin extract inhibited the growth of H. pylori and S. dysenteriae. The highest growth inhibition of S. dysenteriae by L. casei AP was obtained when both bacterial species were grown in inulin extract as the sole carbon source. S. dysenteriae and H. pylori experienced the same level of growth inhibition by L. casei AP when both were grown in inulin-containing media. Meanwhile, the lowest growth inhibition of S. dysenteriae and H. pylori was obtained when either species was grown with L. casei AP in MRS media (Fig. 4). Figure 4 also shows that S. dysenteriae was more easily inhibited by the synbiotic interaction between L. casei AP and inulin extract than H. pylori was (p<0.05). Kabir et al.18 reported that L. casei and L. acidophilus were capable of producing large amounts of lactic acid, thus completely inhibiting the growth of H. pylori in a mixed culture.
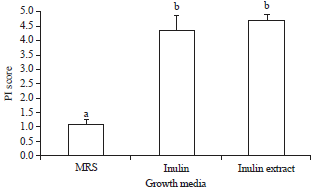 | |
| Fig. 3: | Prebiotic index in three different media. Different letters show significant differences (p<0.05) |
Findings of the current study revealed that the survival of S. dysenteriae and H. pylori when co-cultured with probiotic L. casei AP depended on the media utilized. Widodo et al.6 demonstrated the ability of L. casei AP to inhibit the growth of Shigella flexnerii, E. coli, Staphylococcus aureus and Enterococcus faecalis.
Production of short-chain fatty acids and pH changes: The ability of L. casei AP to produce short-chain fatty acids (SCFAs) was analysed by measuring the concentrations of lactate, acetate, propionate and butyrate. The concentrations of these SCFAs, as percentages (%) are presented in Table 1. The data show that differences in the growth media (MRS, inulin and inulin extract) did not affect the SCFA concentrations. Lactate was present at the highest concentration (33.83-33.53%), followed by acetate (9.83-12.42%), propionate (0.05-0.43%) and butyrate (0.03-0.49%). Lactate and acetate were already known to be the main metabolites produced by lactic acid bacteria13.
The high production of lactate and acetate consequently decreased the pH of the media (Fig. 4). A longer fermentation time resulted in a greater reduction of pH and the lowest pH was observed in growth media containing the inulin extract, followed by inulin-containing media and MRS (Fig. 5). This decreased pH was related to the increased activity of L. casei AP in producing short chain fatty acids. Decreased pH was likely due to the ionization of potential cytotoxic components, such as biogenic amines and ammonia19.
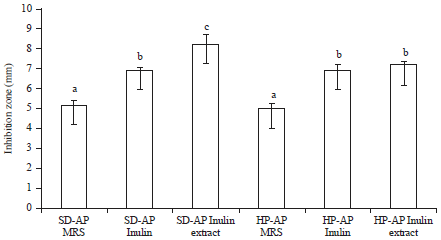 | |
| Fig. 4: | Competition between pathogens and a symbiotic L. casei AP with inulin/inulin extract (SD: S. dysenteriae ATCC 11835, HP: H. pylori ATCC 26695, AP: L. casei AP). Different letters show statistically significant differences (p<0.05) |
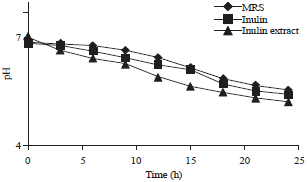 | |
| Fig. 5: | Changes in pH of three different media after inoculation with L. casei AP |
Lactic acid bacteria, particularly of the genus Lactobacillus are acid-producing bacteria and tolerate acidic conditions in the medium, with an optimum pH (5.5-6.4). In addition to the activity of L. casei AP in producing SCFA, it was proposed that E. coli was able to produce acids. Muza-Moons et al.20 reported that enteropathogenic E. coli was also able to produce acids, thus lowering the pH of the media.
This study reveals the ability of inulin extracts obtained from dahlia tubers to support the growth of probiotics and, accordingly, suppress pathogenic bacteria.
CONCLUSION
It is concluded that inulin extracts obtained from dahlia tubers can act as prebiotics that support the growth of probiotics L. casei AP.
The implication of these findings supports the utilization of dahlia tubers as inulin sources and will thus aid in the development of inulin-based food industries, particularly in tropical countries.
SIGNIFICANCE STATEMENT
This study discovers the prebiotic effect of inulin extract produced from local dahlia tubers, which can be beneficial for supporting probiotics in a host’s gastrointestinal tract. This study will support the utilization of dahlia tubers as inulin sources, thus aiding the development of inulin-based food industries, particularly in tropical countries.
ACKNOWLEDGMENT
This study was supported by a Program Pasacasarjana Dalam Negeri (B-PPDN) DIKTI scholarship and by research funding via Hibah Penelitian Unggulan Perguruan Tinggi (PUPT-UGM, 689/UN1-P.III/LT/DIT-LIT/2016). Thanks to Ari Surya Sukarno for preparation of L. casei AP and inulin (Sigma-Aldrich).
REFERENCES
- Nair, K.K., S. Kharb and D.K. Thompkinson, 2010. Inulin dietary fiber with functional and health attributes-A review. Food Rev. Int., 26: 189-203.
CrossRefDirect Link - Kusmiyati, N., T.D. Wahyuningsih and Widodo, 2018. Extraction and identification of inulin-type fructo-oligosaccharides from Dahlia pinnata L. Asian J. Chem., 30: 355-358.
CrossRef - Liong, M.T., 2008. Roles of probiotics and prebiotics in colon cancer prevention: Postulated mechanisms and in-vivo evidence. Int. J. Mol. Sci., 9: 854-863.
CrossRefDirect Link - Nakakuki, T., 2002. Present status and future of functional oligosaccharide development in Japan. Pure Applied Chem., 74: 1245-1251.
CrossRefDirect Link - Widodo, N.S. Anindita, T.T. Taufiq and T.D. Wahyuningsih, 2014. Evaluation of two Lactobacillus strains as probiotics with emphasis in utilizing prebiotic inulin as energy source. Int. Res. J. Microbiol., 5: 33-40.
CrossRefDirect Link - Widodo, T.T. Taufiq, E. Aryati, A. Kurniawati and W. Asmara, 2012. Human origin Lactobacillus casei isolated from Indonesian infants demonstrating potential characteristics as probiotics in vitro. Indonesian J. Biotechnol., 17: 79-89.
Direct Link - Sanz, M.L., G.R. Gibson and R.A. Rastall, 2005. Influence of disaccharide structure on prebiotic selectivity in vitro. J. Agric. Food Chem., 53: 5192-5199.
CrossRefDirect Link - Conti e Silva, A.C., D.H.M. Bastos, J.A.G. Areas, R. Facanali and M.O.M. Marques, 2008. Volatile compounds in the thermoplastic extrusion of bovine rumen. Quim. Nova, 31: 1990-1993.
CrossRefDirect Link - Satirapathkul, C. and T. Leela, 2011. Growth inhibition of pathogenic bacteria by extract of Quercus infectoria galls. Int. J. Biosci. Biochem. Bioinform., 1: 26-31.
Direct Link - Watson, D., M.O. Motherway, M.H.C. Schoterman, R.J.J. van Neerven, A. Nauta and D. van Sinderen, 2013. Selective carbohydrate utilization by lactobacilli and bifidobacteria. J. Applied Microbiol., 114: 1132-1146.
CrossRefDirect Link - Holt, S.M., C.M. Miller‐Fosmore and G.L. Cote, 2005. Growth of various intestinal bacteria on alternansucrase-derived oligosaccharides. Lett. Applied Microbiol., 40: 385-390.
CrossRefDirect Link - Van Laere, K.M., R. Hartemink, M. Bosveld, H.A. Schols and A.G.J. Voragen, 2000. Fermentation of plant cell wall derived polysaccharides and their corresponding oligosaccharides by intestinal bacteria. J. Agric. Food Chem., 48: 1644-1652.
CrossRefDirect Link - Rycroft, C.E., M.R. Jones, G.R. Gibson and R.A. Rastall, 2001. A comparative in vitro evaluation of the fermentation properties of prebiotic oligosaccharides. J. Applied Microbiol., 91: 878-887.
CrossRefDirect Link - Depeint, F., G. Tzortzis, J. Vulevic, K. I'Anson and G.R. Gibson, 2008. Prebiotic evaluation of a novel galactooligosaccharide mixture produced by the enzymatic activity of Bifidobacterium bifidum NCIMB 41171, in healthy humans: A randomized, double-blind, crossover, placebo-controlled intervention study. Am. J. Clin. Nutr., 87: 785-791.
CrossRefDirect Link - Kabir, A.M., Y. Aiba, A. Takagi, S. Kamiya, T. Miwa and Y. Koga, 1997. Prevention of Helicobacter pylori infection by lactobacilli in a gnotobiotic murine model. Gut, 41: 49-55.
CrossRefDirect Link - Topping, D., 2007. Cereal complex carbohydrates and their contribution to human health. J. Cereal Sci., 46: 220-229.
CrossRefDirect Link - Muza-Moons, M.M., E.E. Schneeberger and G.A. Hecht, 2004. Enteropathogenic Escherichia coli infection leads to appearance of aberrant tight junctions strands in the lateral membrane of intestinal epithelial cells. Cell. Microbiol., 6: 783-793.
CrossRefDirect Link