
Afriani
Department of Animal Production, Faculty of Animal Science, Jambi University, Indonesia
LiveDNA: 62.20625
Arnim
Department of Animal Production, Faculty of Animal Science, Andalas University, Padang, Indonesia
Yetti Marlida
Department of Animal Nutrition, Faculty of Animal Science, Andalas University, Padang, Indonesia
LiveDNA: 62.22962
Yuherman
Department of Animal Production, Faculty of Animal Science, Andalas University, Padang, Indonesia
Pakistan Journal of Nutrition
Year: 2018 | Volume: 17 | Issue: 8 | Page No.: 361-367







ABSTRACT
Background and Objective: Proteases are important enzymes and have high economic value due to their wide applications in the food industry as a meat tenderizer. Protease use in the food industry necessitates an understanding of the capabilities and influencing factors of these enzymes to accelerate enzymatic reactions. This study aimed to isolate and characterize the proteases of lactic acid bacteria (LAB) from Bekasam. Methodology: The samples were obtained from the third, fifth, seventh, ninth and eleventh day of fermentation to isolate the proteolytic LAB. Characterization of proteases includes the incubation time, casein substrate concentration, optimum temperature and pH, metal ion contents and stability. The LAB with the highest protease activity is identified molecularly and isolated through 16S rDNA sequencing and phylogenetic analysis based on the Neighbor Joining method. Results: The results showed that the best isolate was BAF-715 because it had the highest protease activity (18.84 U mL–1) at 40 h of incubation. The optimum activity of this protease on a casein substrate at 2.5% occurred at an incubation temperature of 40°C at pH 7 and in the presence of Mg2+ and Mn2+ (5 mM) as activators. Based on molecular DNA identification, the BAF-715 isolate is determined to be Pediococcus pentosaceus. Conclusion: A protease produced by Pediococcus pentosaceus showed the highest proteolytic activity, making it the best protease for application as a beef tenderizer.
PDF Abstract XML References Citation
Received: September 27, 2017;
Accepted: May 04, 2018;
Published: July 15, 2018
Copyright: © 2018. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Afriani, Arnim, Yetti Marlida and Yuherman, 2018. Isolation and Characterization of Lactic Acid Bacteria Proteases from Bekasam for use as a Beef Tenderizer. Pakistan Journal of Nutrition, 17: 361-367.
DOI: 10.3923/pjn.2018.361.367
URL: https://scialert.net/abstract/?doi=pjn.2018.361.367
DOI: 10.3923/pjn.2018.361.367
URL: https://scialert.net/abstract/?doi=pjn.2018.361.367
INTRODUCTION
Bekasam is a traditional salt-fermented product from fish, which are found in many areas in Indonesia. Many microorganisms contain lactic acid bacteria (LAB)1. LAB can produce several metabolites; they can also produce proteolytic enzymes around cell walls, in cytoplasmic membranes and in cells2. Protease is an enzyme that breaks the peptide bonds in proteins to produce simpler compounds, such as small peptides and amino acids.
Protease is a very important enzyme in food and non-food industries. Some uses of proteases in the food industry include reducing turbidity in the beer industry, reducing gluten in the bread industry, agglomerating milk in the cheese industry and tenderizing meat in the meat processing industry3,4. Proteases can be easily isolated from various sources of plants, animals and microbes through fermentation processes5.
Microbes used as an enzyme source are more advantageous than plant and animal sources because microbes can grow rapidly on inexpensive substrates and can be easily enhanced by manipulating the growth conditions and applying genetic engineering6 and biotechnology7. The existence of a superior microbe is an important factor in the production of enzymes.
Several types of LAB have proteolytic activity, such as L. plantarum, L. brevis, L. pentosus, Pediococcus acidilactici, Pediococcus pentosaceus and Lactobacillus spp8-10. LAB are safe microbes and enzymes produced from LAB can be used directly on food.
The ability of proteases to accelerate reactions is influenced by several factors, such as pH, temperature and metals serving as activators or inhibitors, that change enzymatic characteristics. Proteases from Pediococcus acidilactici have an optimum activity at pH 4, an incubation temperature of 28°C and a 2% casein concentration11. The protease activity from Lactobacillus plantarum was highest at 4.43 U mg–1, pH 7, an incubation temperature of 47°C and with a Mg2+ metal activator12. The 220 proteases from P. pentosaceus stabilized at pH 4-7, an incubation temperature of 20-40°C and with metal activators/inhibitors of Ca2+, Zn2+, Mg2+ and Fe3+; the protease activity was inhibited by EDTA and sodium dodecyl sulfate10.
From the description above, this study was conducted to determine the optimum conditions for the production of protease enzymes by LAB.
MATERIALS AND METHODS
Isolation of lactic acid bacteria: LAB was isolated from Bekasam samples on the third, fifth, seventh, ninth and eleventh fermentation days of 2 replications and the bacteria were grown on MRS agar medium. From each sample, five different colonies were grown on MRS agar medium+CaCO3 using the pour plate method. The isolate, which formed a clear zone, was then purified by the streak plate method on the same medium.
Test of proteolytic lactic acid bacteria9: A LAB proteolytic test was used with an agar skim milk (ASM) medium with 3% skim and 3% agar composition. The proteolytic isolates were characterized by the formation of a clear zone on the agar skim medium and then, the clear zone was measured. The proteolytic index was calculated by measuring the ratio of the diameter of the clear zone/colony diameter (R).
Production and isolation of protease enzymes: The production and isolation of protease enzymes are performed by adding one dose of isolate to 50 mL MRS broth and incubating the sample in a shaker incubator with an agitation speed of 150 rpm at 37.5°C for 24 h. The inoculum was then inoculated on protease production media with a composition of 2% skim milk, 0.5% peptone, 0.1% yeast extract, 2% glucose, 0.1% NaCl, 0.008% KH2PO4, 0.01% MgSO4.7H2O and 0.04% (NH4)2SO413. The sample was incubated in a shaker incubator with an agitation speed of 150 rpm at 37.5°C with various production times, namely, 8, 16, 24, 32, 40, 48 and 56 h. The sample was centrifuged at a rate of 12000 rpm for 15 min at 4°C to separate the supernatant and its sediment. The supernatant is a crude extract of the protease and is used for testing the protease activity.
Protease activity measurements: Protease activity was measured by the method of Bergmeyer et al.14 using casein substrate Hammerstein 2% (b/v). A unit of protease activity is defined as the amount of enzyme that can produce one μmol of tyrosine product per minute under the measurement conditions. Isolates with the highest proteolytic activity were tested further.
Protease characterization: Characterization of proteases includes the effect of the casein substrate concentration on enzyme activity, which was tested by reacting an enzyme solution with a casein concentration of 0.5, 1, 1.5, 2, 2.5 and 3%.
The pH effect on the enzyme activity was tested by using different buffer treatments. The universal buffers used had pH values of 3, 4, 5, 6, 7, 8 and 9.
The effect of temperature on enzyme activity was tested on the first incubation carried out at different temperatures, i.e., 30, 40, 50, 60, 70, 80 and 90°C.
The effect of the metal ions Mg2+, Cu2+, Ca2+, Fe2+, Zn2+ and Mn2+ in sulfate salts with a concentration of 5 mM as an activator or inhibitor on protease activity was tested using an optimum pH buffer and optimum temperature. As a benchmark, the treatment used buffer without the addition of metal ions (nonmetal).
Molecular identification of selected LAB isolates: LAB isolates with the highest proteolytic activity were molecularly identified by extracting DNA encoding 16S rRNA. DNA sequencing of the encoded 16S rRNA was performed by 1st Base, PT Genetics of Indonesia. An analysis of the sequencing results was performed using the BLASTing sequence of nucleotides from the sequencing results of 16S rRNA with the database available at www.ncbi.nlm.nih.gov. Phylogenetic analysis was performed with the Neighbor Joining method using the program Molecular Evolutionary Genetics Analysis (MEGA) version 6.0 with Multiple Sequence Comparison by Log Expectation (MUSCLE).
Amplification of DNA encoding 16S rRNA with PCR (polymerase chain reaction): The amplification reaction of the DNA sample was carried out in 0.2 mL PCR tubes. To each PCR reaction tube was added 0.25 μL RBC Taq (5 U mL–1), 10×5 μL Taq buffer (containing Mg2+), 4 μL total of 2.5 mM dNTP and universal primer 63F (5'-CAGGCCTAACACATGCAAGTC-3') and universal primer 1387R (5 'GGGCGGWGTGTACAAGGC-3') at 1.25 μL (20 pmol) and 1.25 μL (20 pmol), respectively; 2.5 μL (100 ng) of genomic extract and ddH2O up to 50 μL were also added. PCR amplification was performed using a PTC 100 PCR instrument (MJ Research, Inc.) at an initial denaturation temperature of 95°C for 5 min, followed by 30 cycles of annealing at 94°C for 30 sec, as well as an extension at 50°C for 1 min, 72°C for 2 min and a final stage of 72°C for 5 min. The PCR products were stored at 4°C for further examination using 1% agarose electrophoresis in 1×TAE, 100 V for 30 min15.
Analysis of the DNA sequence of the genes encoding 16S rRNA: The DNA sequencing of the genes encoding 16S rRNA was performed by using 1st Base, PT, Indonesian Genetics. The sequencing analysis was performed by BLAST to obtain the nucleotide sequence from the 16S rRNA sequencing results with the available database at www.ncbi.nlm.nih.gov. Then, a multiple alignment was performed by using the Clustal W Program. Furthermore, visualization of kinship was performed using a phylogenetic tree combination via the Neighbor Joining method16 with the MEGA 6.0 program17.
RESULTS AND DISCUSSION
Isolation of lactic acid bacteria and proteolytic test of protease-producing bacteria: The isolation process obtained 50 bacterial colonies. Then, from these 50 colonies, which provided clear zones in the MRS and CaCO3 media, 25 isolates showed the characteristics of LAB. The formation of clear zones indicated that the bacteria produced an excess of secondary metabolites (lactic acid) so that an excess of lactic acid was shown by the clear zones around the bacterial colonies.
The 25 isolates of LAB were tested for proteolytic activity, as shown by their ability to form a clear zone on the agar skim medium. When viewed from the protease activity of the clear zone/colony diameter (R) size, the number of isolates exhibiting high protease activity (R>2) was less than that of isolates with R<2 (Table 1). Three isolates of LAB with a high proteolytic index (R>2) were BAF-514, BAF-715 and BAF-1121 (Table 2).
Protease enzyme activity: A qualitative test of protease activity showed that the BAF-514, BAF-715 and BAF-1121 isolates could produce extracellular protease by forming a large clear zone around the cell colony. The larger clear zone that was formed indicated that the microbes were highly capable of changing the substrate that was contained in the medium18.
The proteolytic index of isolates BAF-514, BAF-715 and BAF-1121 was 2.06, 2.29 and 2.02, respectively. The protease activity of the BAF715 isolate in the skim milk agar medium was demonstrated by the presence of clear zones around the colony (Fig. 1). The results reported by Wikandari et al.9 indicated that the LAB with a proteolytic index >2 were L. plantarum N2352, L. plantarum B1765, L. plantarum T2565, L. plantarum B1465, L. pentosus B2555 and Pediococcus pentosaceus B1666.
| Table 1: | Number of isolates of lactic acid bacteria and proteolytic lactic acid bacteria from Bekasam |
 | |
| Table 2: | Chosen isolates of lactic acid bacteria and the proteolytic index |
 | |
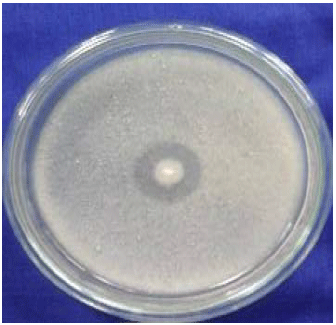 | |
| Fig. 1: | Clear zone as a qualitative indicator of LAB proteases |
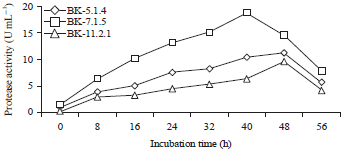 | |
| Fig. 2: | Activity of isolate enzymes BAF-514, BAF-715 and BAF-1121 for different incubation times |
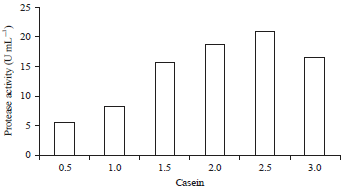 | |
| Fig. 3: | Effect of the casein substrate concentration on the protease activity of the BAF-715 isolate |
Interestingly, research by Yusmarini et al.8 proved that LAB from the genus Lactobacillus R.1.3.2 and R.11.1.2 has a proteolytic zone of 1.725 and 1.650 cm, respectively.
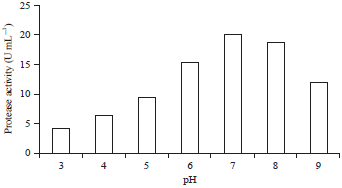 | |
| Fig. 4: | Effect of pH on the protease activity of lactic acid bacteria BAF-715 |
The results for the protease activity as a function of incubation time (Fig. 2) showed that the optimum activity of the BAF-514 and BAF-1121 isolate proteases at 48 h of incubation time was 11.32 and 9.65 U mL–1, respectively. This result indicates that the proteases were produced at a stationary phase, namely, the peak cell growth phase. Meanwhile, the isolate protease activity of BAF-715 at 40 h of incubation time was 18,844 U mL–1, revealing that the protease was produced along with the cell growth before reaching the stationary phase. The value of this protease activity was higher than that of Putranto19. The Lactobacillus acidophilus bacteria were at the highest activity when the incubation time was 18 hours (0.752 U mg–1). Because isolate BAF-715 showed the highest proteolytic activity, this isolate was selected for further research.
Characterization of the crude extract protease of the selected LAB isolate (BAF-715) Effect of the casein substrate concentration: The effect of the casein substrate concentration on the protease activity is shown in Fig. 3. The highest protease activity was obtained at a concentration of 2.5% at 88.4 U mL–1. Meanwhile, the protease activity decreased when the concentration of the casein substrate was 3%. Comparing the results of Oke and Onilude11 study using P. acidilactici and the results of Akinkugbe and Onilude20 study using Lactobacillus acidophilus, the highest protease activity was achieved at a casein concentration of 2%.
Effect of pH: The pH activity profile of an enzyme describes the pH at the time that the proton should be donated or received on the catalytic side of the enzyme to be at the desired ionization level. Figure 4 shows that the isolate protease activity of LAB has an optimum pH of 7 with an enzyme activity of 19.89 U mL–1. Changes in alkaline or acidic pH conditions result in a decrease in enzyme activity. It is estimated that the enzyme activation change is caused by ionization of the enzyme’s ionic group, on the active side or on the other side, thus indirectly affecting the active side. The ionic group plays a role in maintaining the active side conformation for binding the substrate and converting it into the product. These enzyme activity results were higher than that reported in the research of Putranto19, in which Lactobacillus acidophilus bacteria had an optimum pH of 6 with an enzyme activity of 13.5 units mg–1; the protease product of P. acidilactici had an optimum pH of 4.0. The research results by Oke and Onilude11 using the protease from Lactobacillus plantarum had an optimum pH of 7, which was in agreement with the research results by Sulthoniyah et al.12. Ionization can also be experienced by the substrate or substrate-enzyme complex, which also affects enzyme activity21.
Effect of temperature: Temperature is a vital factor affecting protease activity. According to Baehaki and Budiman22, in general, each enzyme has a maximum activity at a certain temperature, meaning that the enzyme activity will increase as the temperature increases until the optimum temperature is reached. The increase in temperature affects the substrate conformation change so that the substrate’s active side encounters barriers while entering the enzyme's active site, causing a decrease in enzyme activity. In Fig. 5, the optimum activity at 40°C was 29.19 U mL–1 and the enzyme activity decreased as the temperature increased. At a temperature of 30°C, there was an increase in protease activity. At 50-90°C, a decrease in protease activation was observed. The stability of an enzyme depends on several things, namely, hydrogen bonding, hydrophobic bonding, ionic interactions and disulfide bridges. The stability of the enzyme in response to temperature can be maintained when the protein’s amino acids form a certain conformation that is resistant to the effects of denaturation; in general, proteins are denatured at high temperatures23. Compared with the Lactobacillus plantarum protease, the optimum temperature is 47°C with an activity of 9.76 U mL–1; the research results by Sulthoniyah et al.12 on the optimum activity of the proteases of Streptococcus lactis and Lactococcus lactis revealed an optimum temperature of 28°C, in agreement with the research results by Akinkugbe and Onilude20.
Effect of metal ions: The results showed that the metal ions that act as activators are Mg2+ and Mn2+ ions at a concentration of 5 mM with an activity of 23.16 and 39.39 U mL–1, respectively.
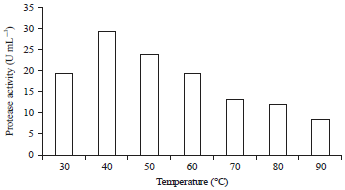 | |
| Fig. 5: | Effect of temperature on the protease activity of lactic acid bacteria BAF-715 |
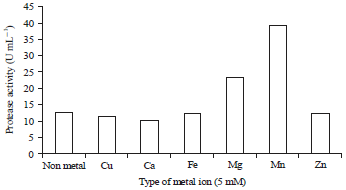 | |
| Fig. 6: | Metal ion effect on the protease activity of lactic acid bacteria BAF-715 |
While with Cu2+, Ca2+, Fe2+ and Zn2+, the protease activity values were 11.54, 10.03, 12.19 and 12.21 U mL–1, respectively, showing a reduction compared with the activity of the treatment without adding metal ions in which the activity was only 12.67 U mL–1, as shown in Fig. 6. Cofactor bonds or enzyme inhibitors may alter the enzyme's ability to bind to the substrate and alter the enzyme's catalytic capacity because the enzyme structure in the presence of an inhibitor or cofactor undergoes physical and chemical changes such that the activity changes24. The research results by Xu et al.10 demonstrated that P. pentosaceus had 220 proteases, the metal activator was the Ca2+ ion, the inhibitors were Zn2+, Mg2+ and Fe3+ and the activity was inhibited by the metal-chelating EDTA and sodium dodecyl sulfate. Regarding the protease of Lactobacillus plantarum, the metal activator was Mg2+ with an enzyme activity of 7.5 UI mg–1, as shown in the research results by Sulthoniyah et al.12. According to Suhartono24, the presence of cofactors or inhibitors may alter the enzyme's ability to bind the substrate and may alter the enzyme's catalytic capacity because the enzyme structure undergoes physical and chemical changes such that the activity changes.
 | |
| Fig. 7: | Phylogenetic trees of isolate BAF-175 based on the 16S rRNA gene with maximum likelihood (ML), neighbor joining (NJ) and maximum parsimony (MP) analysis |
The 0.005 scale shows the evolutionary distance at the branch length, whereas the number on the branch shows the bootstrap value 1000 (ML/NJ/MP) | |
Molecular identification of isolate BAF-715: Genotypic characterization of the LAB isolate was performed based on DNA sequencing of the gene encoding 16S rRNA to determine the genus and species. The DNA encoding the 16S rRNA can be used as a molecular marker for the definition of a species because these molecules exist in every organism and they perform identical functions in all organisms. DNA from isolate BAF-715 was amplified using 63F and 1387R primers. These primers do not form a duplex structure with an enzyme-recognizable 5 edge 5'-3exonuclease and no nucleotides are cut off at the 5' end to affect the primary annealing temperature25.
The DNA sequence analysis of the 16S rRNA BAF-715 isolate obtained sequences with a length of 1200 bp. The sequencing of the 16S rRNA BAF715 isolates compared with sequences in the GenBank NCBI database using the BLAST algorithm. The BLAST result for isolate BAF-715 showed 99% homology/similarity to Pediococcus pentosaceus (KP 18922.8.1) (Fig. 7). The classification of isolate BAF-715 is as follows: Kingdom: Bacteria; Division: Firmicutes; Class: Bacilli; Order: Lactobacillales; Family: Lactobacillaceae; Genus: Pediococcus; Species: Pediococcus pentosaceus.
CONCLUSION
Isolate BAF-715 has a higher protease activity than that of isolate BAF-514 and BAF-1121 (18.48, 11.32 and 9.65 U mL–1, respectively). Isolate BAF-715 shows the highest proteolytic index and activity, with protease activity at an incubation time of 40 h, 2.5% casein substrate, 40°C, pH 7, metal ions Mg2+ and Mn2+ (5 Mm) as activators and metal ions Fe2+, Cu2+, Ca2+ and Zn2+ as inhibitors. The result of the molecular identification with 16S rRNA sequencing and phylogenetic analysis based on the Neighbor Joining method for isolate BAF-715 reveals that it is Pediococcus pentosaceus. A protease produced by Pediococcus pentosaceus from Bekasam showed a high proteolytic activity, making it the best protease for application as a beef tenderizer.
SIGNIFICANCE STATEMENTS
This study discovers the optimum conditions for the production of the protease enzyme by Pediococcus pentosaceus bacteria; the findings can be beneficial for the food industry to improve meat tenderizers. This study will help researchers uncover the ability of Pediococcus pentosaceus to produce a protease enzyme that can be explored by other researchers. Thus, a new theory on the optimum conditions for Pediococcus pentosaceus to produce the protease enzyme may be developed.
ACKNOWLEDGMENTS
This study was funded and supported by the Directorate General of Higher Education of the Ministry of Research, Technology and Higher Education through the Hibah Doktor scheme 2017, No: 48/UN21.17/PP/2017. We are very grateful to the Minister of Research, Technology and Higher Education for allowing us to conduct this study.
REFERENCES
- Rao, M.B., A.M. Tanksale, M.S. Ghatge and V.V. Deshpande, 1998. Molecular and biotechnological aspects of microbial proteases. Microbiol. Mol. Biol. Rev., 62: 597-635.
Direct Link - Takagi, H., M. Kondou, T. Hisatsuka, S. Nakamori, Y.C. Tsai and M. Yamasaki, 1992. Effects of an alkaline elastase from an alkalophilic Bacillus strain on the tenderization of beef meat. J. Agric. Food Chem., 40: 2364-2368.
CrossRefDirect Link - Oyeleke, S.B., E.C. Egwim and S.H. Auta, 2010. Screening of Aspergillus flavus and Aspergillus fumigatus strains for extracellular protease enzyme production. J. Microbiol. Antimicrob., 2: 83-87.
Direct Link - Sandhya, C., A. Sumantha, G. Szakacs and A. Pandey, 2005. Comparative evaluation of neutral protease production by Aspergillus oryzae in submerged and solid-state fermentation. Biochemistry, 40: 2689-2694.
CrossRefDirect Link - Kumar, D. and T.C. Bhalla, 2004. Purification and characterization of a small size protease from Bacillus sp. APR-4. Indian J. Exp. Biol., 42: 515-521.
PubMedDirect Link - Xu, Y., M. Dai, J. Zang, Q. Jiang and W. Xia, 2015. Purification and characterization of an extracellular acidic protease of Pediococcus pentosaceus isolated from fermented fish. Food Sci. Technol. Res., 21: 739-744.
CrossRefDirect Link - Oke, M.A. and A.A. Onilude, 2014. Partial purification and characterization of extracellular protease from Pedicoccus acidilactici. Niger. J. Basic Applied Sci., 22: 19-25.
Direct Link - Sulthoniyah, S.T.M., Hardoko and H. Nursyam, 2015. Characterization of extracellular protease lactic acid bacteria from shrimp paste. J. Life Sci. Biomed., 5: 1-5.
Direct Link - Tamura, K., M. Nei and S. Kumar, 2004. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. U.S.A., 101: 11030-11035.
CrossRefDirect Link - Tamura, K., G. Stecher, D. Peterson, A. Filipski and S. Kumar, 2013. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol., 30: 2725-2729.
CrossRefPubMedDirect Link - Akinkugbe, A.O. and A.A. Onilude, 2013. Selective comparability and physiological studies of lactic acid bacteria protease and Calotropis procera (Linn) extracts. Peer J. Pre, Vol. 1.
Direct Link - Marchesi, J.R., T. Sato, A.J. Weightman, T.A. Martin, J.C. Fry, S.J. Hiom and W.G. Wade, 1998. Design and evaluation of useful bacterium-specific PCR primers that amplify genes coding for bacterial 16S rRNA. Applied Environ. Microbiol., 64: 795-799.
Direct Link