
Akmal
Department of Feed and Technology, Faculty of Animal Science, Jambi University, Jambi, Indonesia
Novirman Jamarun
Department of Ruminant Nutrition, Faculty of Animal Science, Andalas University, Padang, Indonesia
Mardiati Zain
Department of Ruminant Nutrition, Faculty of Animal Science, Andalas University, Padang, Indonesia
LiveDNA: 62.19662
Pakistan Journal of Nutrition
Year: 2018 | Volume: 17 | Issue: 6 | Page No.: 306-310







ABSTRACT
Background and Objective: Manure accumulation in open areas for long periods of time promotes the synthesis of ammonia (NH3) by microbes and its emission to the atmosphere. Forage plants are used as ammonia trapsin poultry farms. This study aims to evaluate the potential of planting dwarf elephant grass and Gliricidia sepium around poultry farms to trap ammonia. Methodology: This study used a complete randomized design with a 2×3 factorial pattern with 6 replications as follows: Factor A: Two species (Dwarf elephant grass and Gliricidia sepium) and Factor B: The distance of the cages from the laying hen (1.5, 3 and 100 m). The parameters measured were the dry matter forage yield, plant height, protein content, ammonia concentration and in vitro dry matter digestibility. Results: The dry matter production of dwarf elephant grass was higher than that of Gliricidia sepium (0.5 vs 0.24 kg pot–1), but for the other parameters, such as the plant height, protein content and dry matter digestibility, the values for Gliricidia sepium were higher than those for dwarf elephant grass. Regarding the plant’s distance from the cage, a distance of 1.5 m yielded the highest levels of dry matter, forage yield, plant height, protein content and in vitro dry matter digestibility. The ammonia concentration at the cage of the laying hen was 7.7 ppm and at a distance of 3 m was 0.88 ppm. Conclusion: Gliricidia sepium was effective at absorbing air borne NH3. Plants 1.5 m from the laying hen cage showed the best results.
PDF Abstract XML References Citation
Received: September 22, 2017;
Accepted: April 18, 2018;
Published: May 15, 2018
Copyright: © 2018. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Akmal, Novirman Jamarun and Mardiati Zain, 2018. Ability of Dwarf Elephant Grass (Pennisetum purpureum Cv. Mott) and Gliricidia sepium to Capture Ammonia (NH3) Around Chicken Cages: An in vitro Evaluation. Pakistan Journal of Nutrition, 17: 306-310.
DOI: 10.3923/pjn.2018.306.310
URL: https://scialert.net/abstract/?doi=pjn.2018.306.310
DOI: 10.3923/pjn.2018.306.310
URL: https://scialert.net/abstract/?doi=pjn.2018.306.310
INTRODUCTION
Forage plants, especially natural grass, are not readily available in Indonesia for the development of ruminants. This is because most of the land has been used for housing, plantation and industry. Therefore, it is necessary to develop forage plants, including superior grasses and leguminous plants that can be planted on limited land with high production. Forage plants include all types of green plants that can be consumed by ruminant livestock, do not poison the animals and provide sufficient nutrients that can meet the needs of livestock. Forage feed is divided into two major groups:Grass (gramineae) and legumes (leguminosae). Both are providers of forage feed1 and represent up to 60% of the forage consumption in cattle2.
The production of cultivated grass is optimal with proper handling, including seed selection, land clearing and fertilizer application. The use of fertilizer on elephant grass planted in ultisol soil with 10 t haSUP>–1/year manure fertilization, 200 kg haSUP>–1/year TSP and 200 kg haSUP>–1/year urea can produce 27.3 t haSUP>–1/year of elephant grass or 4.86 t haSUP>–1/year of dry matter, whereas with the same level of fertilization on king grass yields 82.2 or 14.63 t haSUP>–1/year of dry matter3.
Feed consumed by livestock is partially excreted in the form of manure, which consists of nitrogen (N ammonia). Up to 40% of nitrogen from laying hens travels into the air in the form of NH34 and 25% originates from manure that decomposes in the form of ammonium (NH4+) and nitrate (NO3‾)5. The accumulation of manure in open areas for long periods of time promotes the synthesis of ammonia (NH3) by microbes. This results in atmospheric gas emissions6, which degrade overall air quality and can cause respiratory problems in cage workers7. According to Bittman and Mikkelsen8, NH3 is a trans boundary gas because it can be transferred as far as 10-100 km from its source to contaminate areas with no potential to produce ammonia (NH3).
Plants can absorb surrounding ammonia (NH3) through buds, whereas NH4+ uptake occurs through buds and roots9,10. Uptake of NH3 or NH4+ in the air through the leaf stomata occurs via cellular assimilation through the glutamine synthetase and glutamate synthase (GS-GOGAT) pathways11. NH4+ is converted into glutamine by GS and further converted from 2-Oxoglutarate (2-OG) to glutamate by GOGAT with the help of glutamate dehydrogenase. The N of NH3 manure can be used as a free source of nitrogen in forage fodder, reducing the cost of purchasing N fertilizer. Legumes are able to absorb N-NH3 released from poultry farms to the air12. The content of nitrogen in leguminous plant leaves increases in this environment13,14 and can be used as high-quality ruminant livestock feed. Other forage crops that potentially capture NH3 air include dwarf elephant grass (Pennisetum purpureum cv. Mott) and Gliricidia sepium.
Dwarf elephant grass (Pennisetum purpureum cv.Mott) is a plant that can grow in low-intensity sunlight (30-40%) and adapt to different types of soil, but requires a greater number of tillers and a longer harvest time. Gliricidia sepium is a leguminous tree with leaves that can be used as animal feed. Gliricidia sepium leaves work well as livestock feed because they have a high protein content and grow efficiently, even in the dry season15.
The quality of forage ruminant feed can be determined from its digestibility value. The digestibility value is related to the quantity of food substances that can be absorbed and utilized by livestock. Digestibility is often measured by in vitro systems that mimic the true digestive system. In vitro systems are quite accurate as long as microorganisms and enzymes are sensitive to factors that affect speed and digestion16. The aim of this study was to test the ability of dwarf elephant grass (Pennisetum purpureum cv. Mott) and Gliricidia sepium to utilize ammonia nitrogen (N-NH3) from chicken manure and its effect on production, protein content and digestibility in vitro.
MATERIALS AND METHODS
The design used was a complete randomized design with a 2×3 factorial pattern with 6 replications. Factor A represented the two species [Dwarf elephant grass (A1) and Gliricidia sepium (A2)] and Factor B represented the distance to the chicken cage [1.5 m (B1), 3 m (B2) and 100 m (B3)]. This study used 240 sixty-weeks-old ISA brown hens assigned to 240 individual cages (1 hen/cage). The cage size was 20 m×10 m. Individual metabolic cages were 40 cm× 40 cm×60 cm. All experimental diets were based on corn, soybean meal and palm kernel meal and formulated to be isocaloric (2,700 ME kcal kgSUP>–1) and 17% crude protein. Plants were cultivated in a pot system (Fig. 1). Pots containing the experimental plants (Dwarf elephant grass and Gliricidia sepium) and growth medium were placed inside the socket pots. Plants were watered twice per day and given slow-release standard fertilizer once. Plants were grown for 5 months and trimmed 4 times.
The ammonia concentration (ppm) was measured using BW Technologies GAXT-A-DL Gas Alert Extreme NH3 Single Gas Detector at the cage. The plant height (cm/plant), dry matter forage yield (kg/pot) and protein content (%) according to proximate analysis17 and in vitro dry matter digestibility (IVDMD) (%) were measured among plants18.
 | |
| Fig. 1: | Cage layout on the farm with greenery planted next to the coop |
| Table 1: | Dry matter (DM) forage yield of dwarf elephant grass and Gliricidia sepium (kg/pot) |
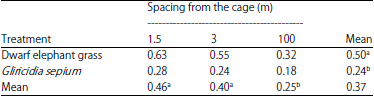 | |
Different superscript letters in rows and columns show significantly different results (p<0.05) | |
| Table 2: | Height of dwarf elephant grass and Gliricidia sepium (cm) |
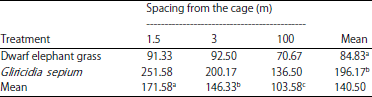 | |
Different superscript letters in rows and columns show significantly different results (p<0.05) | |
Statistical analysis: The data were analyzed by two-way analysis of variance and when significant (p<0.05), they were tested with the Tukey’s test19.
RESULTS AND DISCUSSION
Ammonia content at the study site: The highest concentration of ammonia was found in the laying hen coop (7.7 ppm), while at a distance of 1.5 and 3 m, the concentrations were 2.33 and 0.88 ppm, respectively. The concentration of ammonia was undetectable at a distance of 100 m. The ammonia concentration was strongly influenced by the distance from the exhaust fans13.
Production of dry matter: Dry matter production (DM) of dwarf and gamal elephant grasses is shown in Table 1. The types and spacing of plants showed a very significant effect (p<0.01) on dry matter production. The result of Tukey’s test (p<0.05) showed that the dry matter production of dwarf elephant grass (0.5 kg/pot) was higher than that of gamal (0.24 g/pot), while the spacing showed that B1 (0.46 kg/pot) was not significantly different from that of B2 (0.39 kg/pot), but was significantly different from that of B3 (0.25 kg/pot). The higher production on B1 and B2 shows that N derived from chicken manure can be utilized by mesophyll tissue in plant leaves. Dwarf elephant grass is a fast-growing plant and can be harvested at 80 days, after which it is harvested every 40 days. Each planted grass trimming can produce up to 60 tillers, increasing production. Dwarf elephant grasses grow in clumps with compact fibres and continue to produce saplings when trimmed regularly. Dense dwarf elephant grass can reach heights greater than 1 m20. The production of dry matter by dwarf elephant grass in this study was higher than 0.205 kg/stem21 and approached the dry matter production of Gliricidia sepium (0.31-0.80 kg/stem22).
Plant height: The crop type and spacing of the chicken coop showed very significant differences (p<0.01) relative to plant height (Table 2). Further results from Tukey’s test (p<0.05) on plant type showed that treatments A1 and A2 were significantly different between high-grade dwarf elephant grass (84.83 cm) and gamal (196.17 cm). The difference was caused by the characteristics of each plant. According to Muslihat23, the difference in the height of each treatment is determined by the growth and growth of the cell, where the faster the cell divides and extends or grows, the faster the plant increases in height. Yasin et al.24 stated that the average height of dwarf elephant grass is 117.2 cm and the average height of Gliricidia sepium is 196.17 cm, which is higher than the value reported by Winata et al.25 (170.08 cm). Plant height significantly contributes to forage production.
Further Tukey’s test (p<0.05) of plant spacing from the cage showed that the plant height at B1 (171.58 cm) was significantly different from those at B2 (146.33 cm) and B3 (103.58 cm) and that the plant height at B2 was significantly different from that at B3. The height of the plants decreased in line with an increase in the distance of the plant from the cage because the availability of N-ammonia decreases at a distance of 100 m from the cage, resulting in a decrease in the rate of photosynthesis. The rate of development of plant height after reaching the peak point will decrease with increasing age as the plant gradually decreases the rate of photosynthesis. The photosynthesis products are transported to the growing tissue. The fewer photosynthetic products are transported, the slower the growth. Eventually, the plant will stop growing26.
| Table 3: | Protein content of dwarf elephant Grass and Gliricidia sepium (%) |
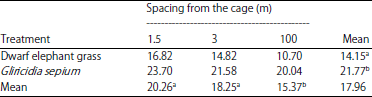 | |
Different superscript letters in rows and columns show significantly different results (p<0.05) | |
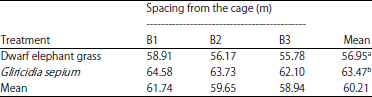
| Table 4: | In vitro dry matter digestibility (%) |
 | |
Different superscript letters in rows and columns show significantly different results (p<0.05) | |
Protein content: The mean protein contents of dwarf and gamal elephant grass are shown in Table 3. Variety analysis of plant species and plant spacing showed that the protein content of both plants differed significantly (p<0.01) from the crude protein content and that there was no interaction between the two factors. Further tests showed a higher crude protein content in Gliricidia sepium (p<0.05) than dwarf elephant grass. The leguminous plant protein content was higher than that of grass because legumes are included in the protein source feed, while the dwarf elephant grass is included in the fibre source feed. The content of dwarf elephant grass protein in this study (14.25%) was higher than that reported by Yasin et al.24 (11.50%), while the protein content of Gliricidia sepium (21.77%) was higher than those of Aderinola and Binuomote27(17.79%).
Further Tukey’s test (p<0.05) showed that the plant protein content in B1 was significantly different from B2 and B3. The decrease in protein content with the increase in distance from the chicken coop was in accordance with a previous study conducted by Adrizal et al.13, who stated that the N content of leaves is higher in plants that grow close to the ammonia gas drain than those that grow further away.
Dry matter digestibility in vitro : The results of plant variety analyses showed significantly different (p<0.01) levels of dry matter digestibility. Further BNJ assay results showed increased digestibility of Gliricidia sepium than dwarf elephant grass because the dwarf elephant grass has higher crude fibre content than Gliricidia sepium. A high content of crude fibre can cause a decrease in feed digestibility. The digestibility of Gliricidia sepium in this study was higher than that of Sukanten et al.28 (52.37%), but lower than that of Ahn et al.29 (79.1%). The in vitro digestibility of elephant grass dry matter in this research was 63.47% (Table 4), close to the value reported by Budiman et al.30 (66.63%). Zailan et al.31 reported that the dry matter digestibilities at 8 and 12 weeks of age were 66.63 and 60.53%, respectively. Yasin et al.24 stated that the digestibility of dry matter leaves in vitro ranged from 71.08-72.93%.
N ammonia from manure has the potential to increase the production and quality of forage feed. Utilization of N ammonia from manure for plants can reduce contamination to the environment. Charles and Hariono32 argued that ammonia gas can pollute the environment and decrease the appearance of livestock, increase the sensitivity of cattle to disease and decrease the work efficiency of the cage workers. Therefore, planting forage around the cage can reduce environmental pollution from ammonia.
CONCLUSION
Gliricidia sepium is effective at absorbing N-NH3 from grass plants. The best distance is 1.5 m from the chicken coop.
REFERENCES
- Patterson, P.H. and E.S. Lorenz, 1996. Manure nutrient production from commercial White Leghorn hens. J. Applied Poult. Res., 5: 260-268.
CrossRefDirect Link - Carlile, F.S., 1984. Ammonia in poultry houses: A literature review. World's Poult. Sci. J., 40: 99-113.
CrossRefDirect Link - Donham, K.J., D. Cumro and S. Reynolds, 2002. Synergistic effects of dust and ammonia on the occupational health effects of poultry production workers. J. Agromedicine, 8: 57-76.
PubMedDirect Link - Bittman, S. and R. Mikkelsen, 2009. Ammonia emissions from agricultural operations: Livestock. Better Crops, 93: 28-31.
Direct Link - Inokuchi, R. and M. Okada, 2001. Physiological adaptations of glutamate dehydrogenase isozyme activities and other nitrogen-assimilating enzymes in the macroalga Bryopsis maxima. Plant Sci., 161: 35-43.
CrossRefDirect Link - Cruz, J.L., P.R. Mosquim, C.R. Pelcani, W.L. Araujo and F.M. DaMatta, 2004. Effects of nitrate nutrition on nitrogen metabolism in cassava. Biol. Planta., 48: 67-72.
CrossRefDirect Link - Yin, Z.H., W. Kaiser, U. Heber and J.A. Raven, 1998. Effects of gaseous ammonia on intracellular pH values in leaves of C3-and C4-plants. Atmos. Environ., 32: 539-544.
CrossRefDirect Link - Adrizal, P.H. Patterson, R.M. Hulet and R.M. Bates, 2006. Growth and foliar nitrogen status of four plant species exposed to atmospheric ammonia. J. Environ. Sci. Health Part B: Pestic. Food Contamin. Agric. Wastes, 41: 1001-1018.
CrossRefDirect Link - Adrizal, A., P.H. Patterson, R.M. Hulet, R.M., Bates and C.A.B. Myers et al., 2008. Vegetative buffers for fan emissions from poultry farms: 2. ammonia, dust and foliar nitrogen. J. Environ. Sci. Health Part B: Pestic. Food Contamin. Agric. Wastes, 43: 96-103.
CrossRefDirect Link - Tilley, J.M.A. and R.A. Terry, 1963. A two-stage technique for the in vitro digestion of forage crops. Grass Forage Sci., 18: 104-111.
CrossRefDirect Link - Liyanage, M.D.S., 1998. Nitrogen fixation and effects of pruning on Gliricidia sepwm and Leucaena leucocephala. INIS Respirat. Res., 29: 41-60.
Direct Link - Muslihat, L., 2003. Teknik percobaan takaran pupuk kandang pada pembibitan abaca. Buletin Teknik Pertanian, 8: 37-39.
Direct Link - Yasin, M., M.A. Malik and M.S. Nazir, 2003. Effect of different spatial arrangements on forage yield, yield components and quality of mott elephantgrass. J. Agron., 2: 52-58.
CrossRefDirect Link - Winata, N.A.S.H., Karno and Sutarno, 2012. Pertumbuhan dan produksi hijauan gamal (Gliricidia sepium) dengan berbagai dosis pupuk organik cair. Anim. Agric. J., 1: 797-807.
Direct Link - Ogunbosoye, D.O. and F.K. Otukoya, 2014. Evaluation of preference and intake of browse species by West African dwarf goats in Nigeria. Int. J. Innov. Res. Dev., 3: 168-176.
Direct Link - Sukanten, I.W., S. Uchida, I.M. Nittis, K. Lana and S. Putra, 1995. Chemical composition and nutritive value of the Gliricidia sepium provenances in dryland farming area in Bali, Indonesia. Asian-Aust. J. Anim. Sci., 8: 231-239.
CrossRefDirect Link - Ahn, J.H., B.M. Robertson, R. Elliott, R.C. Gutteridge and C.W. Ford, 1989. Quality assessment of tropical browse legumes: Tannin content and protein degradation. Anim. Feed Sci. Technol., 27: 147-156.
CrossRefDirect Link - Budiman, B., R.D. Soetrisno, S.P.S. Budhi and A. Indrianto, 2012. Morphological characteristics, productivity and quality of three napier grass (Pennisetum purpureum schum) cultivars harvested at different age. J. Indonesian Trop. Anim. Agric., 37: 294-301.
CrossRefDirect Link - Zailan, M.Z., H. Yaakub and S. Jusoh, 2016. Yield and nutritive value of four Napier (Pennisetum purpureum) cultivars at different harvesting ages. Agric. Biol. J. North Am., 7: 213-219.
Direct Link