
Muhammad Irfan
Laboratory of Physiology and Cell Signaling, College of Veterinary Medicine, Kyungpook National University, 41566 Daegu, Republic of Korea
Man Hee Rhee
Laboratory of Physiology and Cell Signaling, College of Veterinary Medicine, Kyungpook National University, 41566 Daegu, Republic of Korea
Pakistan Journal of Nutrition
Year: 2018 | Volume: 17 | Issue: 1 | Page No.: 25-33







ABSTRACT
Background and Objective: Ethnomedicine and natural products have revitalized the interest in traditional remedies that are being used in prophylactic regimes for cardiovascular diseases. Consumption of tomato and tomato products reduces the hyper-activation of platelets and the process of atherosclerosis and hence reduces the incidence of cardiovascular diseases. The objective of this study was to investigate the antiplatelet activity of water-soluble tomato concentrate (WSTC) and dry water-soluble tomato concentrate (DWSTC) in a detailed manner to identify the underlying mechanism using ex vivo and in vivo experiments with rat platelets for demonstrating the benefits of the dietary supplementation of tomato concentrates which are equivalent to human doses. Methodology: Aggregation was induced by adenosine diphosphate (ADP) and the extent of aggregation was determined using light transmission aggregometry. Granule-secretion, [Ca2+]i and thromboxane B2 were examined. Phosphorylation of signaling molecules, including mitogen-activated protein kinase (MAPK), was investigated by immunoblotting. Prothrombin time and activated partial thromboplastin time were also examined. Moreover, in vivo thrombus wet weight was tested by arteriovenous bypass thrombosis. Results: Both concentrates significantly inhibited agonist-induced platelet aggregation, serotonin release, [Ca2+]i and thromboxane B2 concentration induced by ADP. Moreover, phosphorylation of MAPK pathway factors, i.e., ERK, JNK and P38, was reduced. The concentrates inhibited in vivo thrombus formation without affecting prothrombin time and activated partial thromboplastin time. Conclusion: Both concentrates possess ethnomedicinal properties to prevent cardiovascular ailments and can be used as a functional food or dietary supplement.
PDF Abstract XML References Citation
Received: August 08, 2017;
Accepted: November 29, 2017;
Published: December 15, 2017
Copyright: © 2018. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Muhammad Irfan and Man Hee Rhee, 2018. Tomato Concentrates Inhibit Platelet Function and Thrombus Formation in Rats. Pakistan Journal of Nutrition, 17: 25-33.
DOI: 10.3923/pjn.2018.25.33
URL: https://scialert.net/abstract/?doi=pjn.2018.25.33
DOI: 10.3923/pjn.2018.25.33
URL: https://scialert.net/abstract/?doi=pjn.2018.25.33
INTRODUCTION
Platelet activation underlies thrombotic cardiovascular diseases such as atherosclerosis, thrombosis, myocardial infarction and coronary artery disease (CAD), which are responsible for significant numbers of deaths and disabilities and their prevalence in developed countries is quickly increasing1. Activation of platelets plays crucial role in hemostasis and their hyperactivation is the causal factor for the growth and advancement of atherosclerosis2. Dietary compounds are being given focused attention in efforts to slow down the progression of these diseases3. The popularity of ethnomedicine and natural products has increased and strengthened the interest in traditional remedies for cardiovascular disease (CVD)4. During the last few decades, the consumption of tomatoes has increased and previous studies provide evidence for the overall health benefits of tomatoes, leading them to be considered cardiovascular-protective foods5,6. Nutrients available in the tomato are associated with theoretical or proven effects on reducing the risk of degenerative and cardiovascular system diseases6, such as the cardioprotective effects of lycopene. Previous studies have proposed a link between a lower incidence of CVD and the consumption of tomatoes in Mediterranean countries. A previous study7 reported the antiplatelet activity of water-soluble tomato concentrate (WSTC, Fruitflow I) at 900 mg kg–1 and that of dry water-soluble tomato concentrate (DWSTC, Fruitflow II) at 45 mg kg–1. In the present study, low doses of WSTC (300 mg kg–1) and DWSTC (15 mg kg–1) were investigated, which are equivalent to human doses, to explore the benefits of dietary supplementation as a prophylactic regime for cardiovascular disorders.
MATERIALS AND METHODS
Chemical preparation: WSTC and DWSTC were obtained from DSM Nutritional Products Ltd. (Basel, Switzerland). The DWSTC was diluted to the appropriate concentration immediately before all experiments were performed. Adenosine diphosphate (ADP) was purchased from Chrono-log (Havertown, PA, USA) and Fura-2/AM was obtained from Sigma Chemical Co. (St. Louis, MO, USA). TXB2 EIA kits were purchased from Enzo Life Sciences (Plymouth Meeting, PA, USA) and serotonin EIA kits were purchased from Labor Diagnostika Nord GmbH and Co. KG (Nordhorn, Germany). Antibodies against phospho-p44/42 (phospho-ERK), p44/42 (ERK), phospho-p38, p38, phospho-SAPK/JNK and SAPK/JNK were purchased from Cell Signaling (Beverly, MA, USA). All chemicals were of reagent grade.
Animals and dosage: Male sprague Dawley rats weighing from 240-250 g (6 weeks old) were obtained from Orient Co. (Seoul, Korea). Rats were acclimatized for 1 week before the experiments and maintained in an air-conditioned animal room with a 12/12 h light/dark cycle at a temperature and humidity of 23±2°C and 50±10%, respectively. Rats were randomly divided into three groups, a normal group, WSTC-treated group (300 mg kg–1) and DWSTC-treated group (15 mg kg–1) (n = 10 in each group and n = 4 in each group for the AV shunt model). The normal dose for human consumption is 3 g/day, which is a dose of tomato extract equivalent to 2.5 tomatoes. Dose normalization for rats performed to obtain 300 and 15 mg kg–1 for WSTC and DWSTC, respectively as previously described by Reagan-Shaw et al.8. Rats were orally administered both concentrates once a day for 4 weeks. At the end of the 4th week, blood was collected from the rats for analysis. All experiments were carried out according to IACUC guidelines and approved by the Ethics Committee of the College of Veterinary Medicine, Kyungpook National University, Korea (Permit number: 2015-0094).
Platelet preparation and aggregation assay: Whole blood was obtained by heart puncture 2 h after the last oral administration and platelets were prepared as previously described by Park et al.9. Platelet aggregation was performed as previously described by Jeon et al.10 using light transmission aggregometry. Briefly, the washed platelets (3×108 mL–1) were pre-incubated at 37°C for 2 min in the presence of 1 mM CaCl2 and stimulated with doses of ADP. The mixture was further incubated for 5 min with stirring and measured using an aggregometer (Chrono-log, Havertown, PA).
[Ca2+]i measurement: The intracellular calcium ion concentration ([Ca2+]i) was measured with Fura-2/AM as previously described Kim et al.11, Fura-2 fluorescence in the cytosol was measured with a spectrofluorometer (F-2500, Hitachi, Japan) and [Ca2+]i was calculated as previously described by Schaeffer and Blaustein12 using following formula:
![]()
where, 224 nM is the dissociation constant of the Fura-2-Ca2+ complex and Fmin and Fmax represent the fluorescence intensity levels at very low and very high Ca2+ concentrations, respectively.
Serotonin release assay: The platelet aggregation assay was carried out as described by Jeong et al.7. After terminating the aggregation reaction, the mixture was immediately centrifuged at 12,000 rpm for 5 min at 4°C. The supernatant was collected and serotonin release was measured with a serotonin ELISA kit (Labor Diagnostika Nord GmbH and Co. Nordhorn, Germany) according to the manufacturer’s instructions.
Evaluation of TXA2 generation: The platelet aggregation assay was performed and the reaction was terminated by adding ice-cold 2.5 mM EDTA and 100 μM indomethacin. The mixture was centrifuged at 12,000 rpm for 5 min at 4°C, the supernatant was collected and the concentration of TXB2 (stable metabolite of TXA2) was measured using a TXB2 EIA kit according to the manufacturer’s protocol.
PT/aPTT measurement: The activated partial thromboplastin time (aPTT) and prothrombin time (PT) were measured using an Automated Coagulation Laboratory 100 Instrument (Instrumentation Laboratory Co., Milan, Italy). Briefly, the PPP from rats treated with WSTC and DWSTC was incubated at 37°C for 7 min. Then, 100 L of the incubated PPP was mixed with 50 mL of cephalin in the process plate and coagulation was started with the addition of 1 mM CaCl2 and 100 mL thromboplastin to the PPP for the aPTT and PT assays, respectively.
A-V shunt model: The in vivo antithrombotic activities of WSTC and DWSTC were evaluated in a rat extra corporeal shunt model using the method of Umetsu and Sanai13, with slight modifications. Briefly, the arteriovenous shunt model was used and blood circulation in the cannula was carried out for 15 min, after which thrombus weight was determined immediately.
Immunoblotting: Immunoblots were performed as described by Endale et al.14. Briefly, after terminating the aggregation reaction, lysates were prepared by solubilizing and centrifuging the platelets in sample lysis buffer. Protein concentration was determined using a bicinchoninic acid assay. Total cell proteins (35 μg) from the platelet lysate were separated using 10% SDS-PAGE and transferred to polyvinylidene difluoride membranes in transfer buffer. The membranes were blocked in TBS-T containing 5% dry skim milk and incubated with primary antibody diluted in 5% bovine serum albumin solution. The blots were then incubated with horseradish peroxidase-conjugated secondary antibody that was visualized by binding an enhanced chemiluminescence (Advansta, USA).
Statistical analysis: Data were analyzed with one-way analysis of variance followed by a post hoc Dunnett's test in order to measure the statistical significance of the differences observed (SAS Institute Inc., Cary, NC, USA). All data are presented as the mean±standard error of the mean (SEM). p-values of 0.05 or less were considered to be statistically significant.
RESULTS
Inhibitory effects of tomato concentrate on agonist-induced platelet aggregation and intracellular calcium elevation: The effect of the concentrates on agonist-induced platelet aggregation was evaluated. Firstly, the effects of various agonists (collagen, ADP and thrombin) was investigated and found concentrates to be more effective against the ADP-induced ex-vivo platelet aggregation. Results showed that WSTC (300 mg kg–1) inhibited ADP-induced platelet aggregation. The results with DWSCT (15 mg kg–1) were not significant (Fig. 1). Moreover, intracellular calcium ([Ca2+]i) release plays key role in platelet degranulation and aggregation15. Therefore, it was examined whether concentrates affected [Ca2+]i . As shown in Fig. 2, both concentrates significantly decreased [Ca2+]i mobilization.
Inhibitory effect on granule secretion (serotonin release) and TXB2 production: Activated platelets release the contents of granules such as alpha-granules and dense granules and the secretion of the contents of these granules enhances platelet activation, including intracellular signaling pathways. The effect of serotonin release from dense granules was evaluated. The results indicated that both concentrates inhibited serotonin release (Fig. 3a).
Thromboxane A2 amplifies platelet activation during hemostasis as a mediator. Because it is physiologically unstable, the effect on the production of its metabolite, TXB2 was investigated.
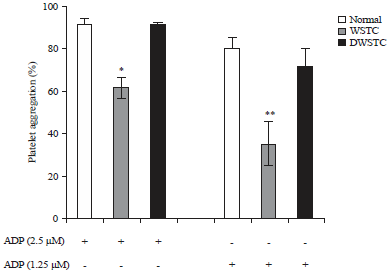 | |
| Fig. 1: | Effect of tomato concentrates (WSTC: 300 mg kg–1 and DWSTC: 15 mg kg–1) on ADP (2.5 μM, 1.25 μM)-induced platelet aggregation ex vivo. Aggregation was quantified and expressed as a percentage |
Each graph shows the Mean±SEM of at least four independent experiments. *p<0.05 and **p<0.01 compared to the agonist control | |
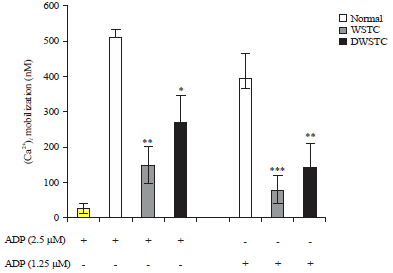 | |
| Fig. 2: | Tomato concentrates (WSTC: 300 mg kg–1 and DWSTC: 15 mg kg–1) inhibited intracellular calcium mobilization. Washed platelets (3×108 mL–1) were incubated with a calcium fluorophore (5 μM, Fura-2/AM) and stimulated with ADP (2.5 μM or 1.25 μM) |
The results are presented as the Mean±SEM of at least four independent experiments. *p<0.05, **p<0.01 and ***p<0.001 compared to the agonist control | |
The results showed that WSTC and DWSTC decreased ADP-induced TXB2 production (Fig. 3b).
WSTC and DWSTC inhibit in vivo thrombus formation: The AV shunt thrombosis model is known to be useful for evaluating in vivo antithrombotic effects13. Therefore, the effect of tomato concentrates on an extracorporeal shunt model was investigated to determine their inhibitory effects on thrombus formation. As shown in Fig. 4, WSTC and DWSTC significantly reduced thrombus formation by 35 and 46%, respectively, compared to the negative control.
Coagulation parameters (PT/aPTT) remain unaltered: The effects of both concentrates on coagulation parameters were determined by aPTT and PT assays using rat PPP. The results indicated that PT and aPTT were not affected by the WSTC and DWSTC treatments (Fig. 5).
WSTC and DWSTC attenuate agonist-induced MAPK phosphorylation: In order to identify the underlying pathway of the effects of WSTC and DWSTC on platelet activation, it is evaluated their effects on protein phosphorylation in an intracellular signaling pathway. It is well known that the phosphorylation of MAPK (ERK, JNK and P38-MAPK) mediates the platelet activation pathway.
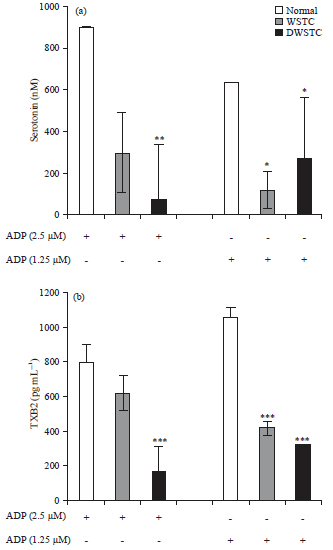 | |
| Fig. 3: | Inhibitory effects of tomato concentrates (WSTC: 300 and DWSTC: 15 mg kg–1) on serotonin, (a) thromboxane and (b) generation. After termination of the aggregation reaction, serotonin release and TXB2 production were assessed using a serotonin ELISA kit and TXB2 EIA kit according to the manufacturer’s protocols |
Bar graph shows the Mean±SEM of at least four independent experiments. *p<0.05, **p<0.01 and ***p<0.001 compared to the agonist control | |
Therefore, it is determined whether both concentrates attenuated MAPK phosphorylation. WSTC and DWSTC inhibited the ADP-induced phosphorylation of ERK and JNK but not P38-MAPK (Fig. 6).
DISCUSSION
CVD is considered a major lethal disease in the Western world and atherosclerosis (i.e., atherothrombosis) is its major underlying cause, which leads to stroke and death (www.oms.org).
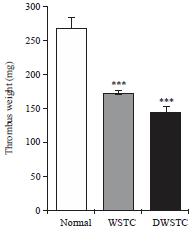 | |
| Fig. 4: | In vivo effects of tomato concentrates (WSTC: 300 and DWSTC: 15 mg kg–1) on thrombus formation. Briefly, the arteriovenous shunt model was used, blood circulation in the cannula was carried out for 15 min and thrombus weight was determined immediately |
Bar graph shows the Mean±SEM of at least four independent experiments. ***p<0.001 versus control | |
The prophylactic and pharmacological suppression of platelet activation is thought to be one of the best prevention methods for CVD16 and they act by slowing down the development of atherothrombosis and hence decreasing the risk of stroke17. The results from primary prevention trials are available, which recommend that the side effects of prophylactic regimes outweigh their benefits18.
Previous studies have reported the antiplatelet activity of tomato extracts, however, the present study explored the mechanistic aspects of the antiplatelet activity of tomato concentrates at a concentration equivalent to the human dose19,20. According to a previous study7, antiplatelet activity of water-soluble tomato concentrate at 900 mg kg–1 and that of dry water-soluble tomato concentrate at 45 mg kg–1. In the present study, low doses of WSTC and DWSTC, which are equivalent to human doses, were investigated to explore the benefits of dietary supplementation as a prophylactic regime for cardiovascular disorders. Firstly, effects of both concentrates on agonist-induced platelet aggregation were investigated. As expected, the current study showed that WSTC inhibited ADP-induced platelet aggregation in a dose-dependent manner, whereas, DWSCT only slightly reduced ADP-induced platelet aggregation, the data were not statistically significant but indicated an inhibitory tendency. The consequences of ADP-induced signaling through its P2Y12 receptor include platelet aggregation and granule secretion21.
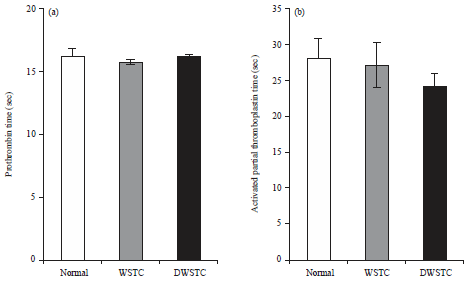 | |
| Fig. 5: | Effects of tomato concentrates (WSTC: 300 mg kg–1 and DWSTC: 15 mg kg–1) on coagulation parameters. Briefly, 100 μL of the incubated PPP was mixed with 50 mL of cephalin in the process plate and coagulation was started with the addition of 1 mM CaCl2 and 100 mL thromboplastin to the PPP for the aPTT and PT assays, respectively |
Bar graph shows the Mean±SEM of at least three independent experiments. Differences were not statistically significant | |
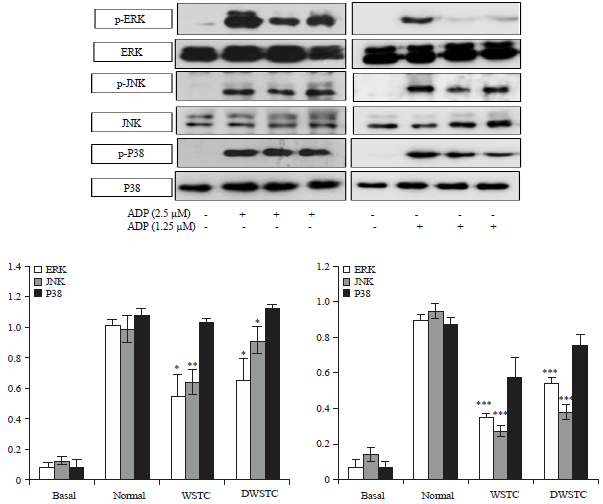 | |
| Fig. 6: | Tomato concentrates (WSTC: 300 mg kg–1 and DWSTC: 15 mg kg–1) attenuated the phosphorylation of MAPKs (i.e., ERK1/2, JNK and p38). Briefly, after terminating the aggregation reaction, lysates were then prepared by solubilizing and centrifuging the platelets in sample lysis buffer. Proteins were then extracted, separated, transferred to PVDF membranes and probed with respective antibodies |
All immunoblots were carried out in at least four independent experiments. *p<0.05, **p<0.01 and ***p<0.001 compared to the agonist-treated group | |
Therefore, in order to clarify the inhibitory mechanism of the tomato concentrates, downstream signaling components, including calcium mobilization and granule secretions were examined. Moreover, platelet granule molecules play key roles in hemostasis and thrombosis, including activating other cells or cellular adhesion22. The role of selected tomato concentrates in the secretion of dense granules by platelets was studied by performing a serotonin release assay. Both concentrates reduced serotonin release in ADP-induced platelet aggregation. These results demonstrated that the anti-platelet effect of these concentrates occurs through its suppressive effect on platelet dense granules secretions. It is reported that components of tomato extract inhibit different pathways of platelet aggregation and suggested that total active fraction (tAF) and its subfractions prevent P-selectin (α-granules secretion) expression and activation of integrin αIIbβ3 (i.e., GPIIb/IIIa)23. Expression of this integrin and P-selectin is specific to platelets and megakaryocytes and activation of these proteins is essential for platelet aggregation19,23.
It is well-known that cytosolic calcium plays a key role in platelet activation. Increasing calcium levels lead to several signaling pathways involved in the actin-myosin interaction and protein kinase C (PKC)24. The results of Ca2+ mobilization study indicated that tomato concentrates significantly decreased the intracellular calcium concentration in ADP-induced platelet aggregation. TXA2 production from arachidonic acid via the cyclo-oxygenase pathway is an important positive feedback loop mechanism for platelet activation25. The significant reduction of its production by both concentrates was observed in ADP-induced platelet aggregation. The current study indicated the suppression potential of tomato on P2Y12, thus, the antagonism of this receptor with a natural therapy may represent a novel therapeutic regime.
Many lines of evidence indicate that platelets are constantly exposed to a variety of activating factors, including collagen, fibrinogen, ADP, vWF, thrombin and thromboxane and inhibitory factors such as endothelial-derived NO, prostacyclin (PGI2) and ADPase26. Thrombotic or bleeding disorders can develop following the disruption of this equilibrium. Thus, a durable equilibrium between the two opposing processes of platelet activation and inhibition is thought to be critical for normal hemostasis. The present study suggests that the pretreatment of activated platelets with WSTC and DWSTC may contribute to the maintenance of this balance. The PT and aPTT results did not differ between the normal and treatment groups, suggesting that neither concentrate affected the integrity of the extrinsic and intrinsic cascades of the coagulation system.
Adam et al.27 established that the MAPKs ERK2, JNK1 and p38 of the MAPK are present in platelets and activated by several agonists. ERK2 amplifies collagen-induced platelet secretion28, whereas, ERK2 activation is dependent on TXA2 production and ADP release in agonist-induced platelet aggregation29. p38MAPK involves in platelet spreading and adhesion30 and their inhibitors have shown suppression of platelet activation31. Kim et al.32 also has shown the involvement of MAPKs in the regulation of platelet adhesion, spreading and aggregation. In the present study, the results indicated that WSTC and DWSTC inhibited the ADP-induced phosphorylation of MAPKs such as ERK1/2 and JNK.
The antiplatelet effects of both tomato concentrates may be attributed to their bioactive compounds (nucleoside derivatives [F1], phenolic conjugates [F2], flavonoid derivatives [F3] including quercetin derivatives) as stated by O’Kennedy et al.23. Results indicated that DWSTC inhibited ex vivo Ca2+ mobilization, TXA2 production, serotonin release and MAPK phosphorylation as well as in vivo thrombus formation but was unable to significantly inhibit ex vivo platelet aggregation, although it showed an inhibitory trend. This may because aggregation is the summation of a complex pathway involving the transduction of multiple signals within the platelets, which cause platelet activation. The effect of given compound on all other factors (such as ATP secretion, P-selectin expression and cGMP or cAMP production) that may contribute to aggregation could not be explored in this study.
CONCLUSION
In summary, tomato concentrates inhibited ex vivo platelet aggregation, [Ca2+]i mobilization and serotonin and TXB2 release and reduced in vivo thrombus formation without affecting coagulation parameters. Findings of the present study show that both concentrates inhibit ADP-induced platelet activation through the modulation of downstream signaling via the MAPK pathway.
SIGNIFICANCE STATEMENT
This study discovers the mechanistic aspects of the antiplatelet activity of tomato concentrates at a concentration equivalent to the human dose. Hence, benefits of dietary supplementation as a prophylactic regime for platelet related cardiovascular disorders could be presumed. Moreover, future studies might be intended to uncover other beneficial health aspects of tomato concentrates.
ACKNOWLEDGMENT
This study was supported by the National Research Foundation of Korea. Grant funded by the Korean government (Grant No. 2015R1D1A1A09057204).