
Amira M. Amr
Department of Public Health Nutrition, Al-Quds University, Jerusalem, Palestine
LiveDNA: 962.19398
Hamed R. Takruri
Department of Nutrition and Food Technology, The University of Jordan, Amman, Jordan
LiveDNA: 962.15132
Maha S. Shomaf
Department of Pathology, The University of Jordan, Amman, Jordan
Omar A. Alhaj
Department of Nutrition, Petra University, Amman, Jordan
Mo`ez Al-Islam E. Faris
Department of Clinical Nutrition and Dietetics, University of Sharjah, Sharjah, United Arab Emirates
LiveDNA: 962.14121
Wael M. Abdel-Rahman
Department of Medical Laboratory Sciences, University of Sharjah, Sharjah, United Arab Emirates
Pakistan Journal of Nutrition
Year: 2018 | Volume: 17 | Issue: 4 | Page No.: 179-189







ABSTRACT
Background and Objective: Camel milk is a folk remedy that includes valuable nutrients and bioactive zoochemicals. However, the chemopreventive potential of camel milk against colon carcinogenesis is poorly understood. This study was conducted to investigate the chemopreventive potential of camel (Camelus dromedarius) and bovine milk as well as the impact of fermenting these milks with Lactobacillus acidophilus and Streptococcus thermophilus against early colon carcinogenesis as measured by the reduction of aberrant crypt foci (ACF) in azoxymethane (AOM)-treated Fischer 344 rats. Methodology: Each of 60 weanling male rats was assigned to one of 6 experimental diet groups: Fermented and unfermented camel milk with AOM, fermented and unfermented bovine milk with AOM and positive (PC, AOM only) and negative (NC, saline vehicle only) control groups. The animals were fed the corresponding diets for 3 weeks and then received two subcutaneous injections of AOM or vehicle for 2 consecutive weeks and they were then placed on the corresponding diets for 11 weeks. At termination, all rats were euthanized, colons were harvested and the ACF counts were determined for all tested groups. Immunohistochemical testing was then performed to examine cell proliferation and apoptosis in the camel milk groups. Results: Significant reductions (p<0.05) (48.4-62.1%) in the total ACF count were observed in the colons of the rats fed all milk diets compared with rats fed on PC. However, significant differences were not observed in the total ACF between the camel and bovine milk diets or between the fermented and unfermented milk diets. In addition, significant changes were not observed in the apoptotic index for the camel milk diet compared with the index values for PC and β-catenin was generally localized to the membrane in all examined specimens. Conclusion: By virtue of its bioactive components, camel milk exhibited a chemopreventive potential against early colon carcinogenesis, however, fermentation did not improve its chemopreventive potential.
PDF Abstract XML References Citation
Received: June 15, 2017;
Accepted: December 26, 2017;
Published: March 15, 2018
Copyright: © 2018. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Amira M. Amr, Hamed R. Takruri, Maha S. Shomaf, Omar A. Alhaj, Mo`ez Al-Islam E. Faris and Wael M. Abdel-Rahman, 2018. Fermented Camel (Camelus dromedarius) and Bovine Milk Attenuate Azoxymethane-induced Colonic Aberrant Crypt Foci in Fischer 344 Rats. Pakistan Journal of Nutrition, 17: 179-189.
DOI: 10.3923/pjn.2018.179.189
URL: https://scialert.net/abstract/?doi=pjn.2018.179.189
DOI: 10.3923/pjn.2018.179.189
URL: https://scialert.net/abstract/?doi=pjn.2018.179.189
INTRODUCTION
Colorectal cancer (CRC) is a major cause of morbidity and mortality worldwide and it accounts for over 9% of all cancer cases and represents the third most common cancer worldwide1. Epidemiological evidence has shown that the majority of CRC cases of the sporadic (nonfamilial) type could be prevented by application of cancer chemoprevention and screening techniques2. Dietary factors are responsible for 70-90% of CRC cases and diet optimization can prevent most of these cases3,4. Most epidemiological and experimental studies conducted to determine the effects of milk and dairy product consumption on neoplastic development have focused on bovine milk5-8, whereas information on camel milk and other types of milk are scarce.
Natural products are important ingredients for pharmaceutical applications, particularly new therapies for preventing or treating cancer and other diseases9. Several studies have shown that camel milk is an important functional food with the potential to provide health benefits for the treatment of different ailments10. These potential health benefits are attributed to the presence of a plethora of bioactive zoochemicals, including lactoferrin, immunoglobulins and peptidoglycan recognition protein11, however, other functional components could be derived from camel milk proteins via fermentation using probiotic strains12. The reported health benefits of these bioactive components include hypotensive activity13, hypoglycemic effects14, oxidative stress reduction15, hypocholesterolemic activity and anti-carcinogenic effects16. Thus, camel milk is used in alternative medical practices to treat many diseases, including cancer16.
Azoxymethane (AOM) is an organotrophic genotoxic colon carcinogen that is activated by cytochrome P450 and then hydroxylated. Further metabolism by colonic bacteria that produce β-glucuronidase generates methyldiazonium, which can initiate cancer by inducing mutagenic lesions in target cells by the alkylation of DNA at the O6 position of guanine17. A significantly positive correlation has been found between fecal β-glucuronidase activity and precancerous aberrant crypt foci (ACF). Strains of gut bacteria that have a low ability to produce β-glucuronidase, such as probiotic bacteria (i.e., Bifidobacteria and lactic acid bacteria) are thought to provide protection against the development of colon cancer18.
The ACF represent the earliest identifiable preneoplastic lesions in colon carcinogenesis and are observed on the surface of the colon and they appear as large, thick crypts in methylene blue-stained colon specimens after treatment with chemical carcinogens, such as AOM19, 20. ACF assays have been widely used as an intermediate or endpoint to test the efficacy of nutritional factors for the prevention of CRC21,22. β-catenin is a transcriptional activator that plays an important role in epithelial tumorigenesis and belongs to the β-catenin/Tcf oncogenic pathway. β-catenin mutations are frequent (66%) in dysplastic ACF and alterations of the cellular localization of β-catenin have been observed in all dysplastic ACF23. These studies indicate a genetic alteration associated with Wnt/β-catenin signaling and these changes may play an important role in AOM-induced colon carcinogenesis. The disruption of Wnt/β-catenin signaling represents a mechanisms underlying carcinogenesis in AOM-induced colon cancer models24. Apoptosis or programmed cell death, plays a fundamental role in the maintenance of tissues and organ systems by providing a controlled cell deletion to balanced cell proliferation. Many dietary chemopreventive agents can preferentially inhibit the growth of tumor cells by targeting one or more signaling intermediates leading to induction of apoptosis. Therefore, induction of apoptosis could be considered as one of the proposed mechanisms of chemoprevention against cancer25.
Because camel milk contains a number of bioactive components that display anticarcinogenic activity and oxidative-stress-reducing potential, it is hypothesized that this milk might display significant chemopreventive potential against chemically induced colon carcinogenesis in the Fischer 344 rats model. Furthermore, because milk fermentation has been reported to exaggerate and enhance the health-promoting effects of the milk, it was also hypothesized that fermenting camel milk would improve its chemopreventive potential. To the best of our knowledge, experimental studies have not been previously conducted to investigate the in vivo chemopreventive potential of camel milk against chemically induced colon carcinogenesis.
Reports from the Middle East indicate that camel milk may also have superior chemopreventive properties over bovine milk, thus, cancer patients frequently consume camel milk16. Several studies have revealed the chemopreventive potential of bovine milk5-8, thus, bovine milk was used for comparison purposes in this study. The aim of the current study was to examine the chemopreventive potential of fresh/unfermented and fermented camel milk and compare the chemopreventive potential between camel and bovine milk during the initiation stage of CRC induced by AOM in the Fischer 344 rats model using ACF as a principal histological biomarker. Furthermore, this study elaborated on the molecular and cytological changes associated with the chemopreventive effect of camel milk against chemically induced colon cancer in rodents.
MATERIALS AND METHODS
Preparation of unfermented and fermented lactose-hydrolyzed milk: Both camel (Camelus dromedarius, sub-species Hamra) and bovine (Bos taurus) milk was purchased from local farmers. The milk samples were tested to identify the presence of antibiotics (β-lactam and tetracycline) using the Twinsensor test kit (Unisensor, Antwerp, Belgium). Fresh whole camel and bovine milk was pasteurized separately at 85°C for 3 min and then cooled to 37°C in a water bath. Lactose hydrolysis was performed for both milks to avoid the diarrhea-inducing effect of lactose on rats because rats have been reported to be lactose intolerant4. Lactose-free milk was prepared using the lactase enzyme as described by Jelen and Tossavainen26. Lactase enzyme (β-D-galactosidase, E.C. 3.2.1.23, Christen Hansen, Danisco, ATCC Microbiology, Manassas, Virginia, USA) was added to the unfermented milk at a concentration of 0.1% (1.0 g L–1) for 24 h at 4°C. The fermented milk was prepared by inoculation with lactase enzyme at a concentration of 0.05% (0.5 g L–1) along with 2% Lactobacillus acidophilus (ATCC 0885S, 314) and Streptococcus thermophilus (ATCC 0136S, 19258), which were purchased from Microbiologics, Inc. (Minnesota, USA). The milk was then incubated for 18 h at 37°C until pH 4.5 was attained.
Preparation of experimental diets: A modified AIN-93G diet formula that replaces casein with egg white solids (AIN-93G-EGG) as the protein source was used as a standard control diet27. In addition, four experimental lactose-hydrolyzed milk diets were prepared: Unfermented camel milk (UCM), fermented camel milk (FCM), unfermented bovine milk (UBM) and fermented bovine milk (FBM). Modifications to the dietary components for the camel and bovine milk diets were made based on the nutrient composition of both milk sources28 and adjustments were made to generate isocaloric and isonitrogenic diets. Both milk sources contributed to 10% of the energy content of their corresponding experimental diets. Diets were dried in oven at 50°C until moisture content reached <5%. The composition of AIN-93G-EGG and other experimental diets are shown in Table 1.
Animal experimentation: This study was approved by the ethical committee for animal experimentation at the University of Jordan. Each of sixty 5-6-weeks-old male weanling specific-pathogen-free Fischer 344 (F344)/NHsd inbred rats were randomly allocated into one of six experimental groups (10 rats/group): Negative control (NC) and positive control (PC) groups, fermented and unfermented camel milk groups and fermented and unfermented bovine milk groups.
Each animal was housed in a single plastic cage (North Kent Plastic Cages, Ltd, Dartford, UK) and food and water were available ad libitum. Artificial light was supplied from neon tubes (12:12 h light:dark cycles). The relative humidity and temperature were maintained at 50±5% and 22±2°C, respectively.
| Table 1: | Dietary composition of experimental and control diets |
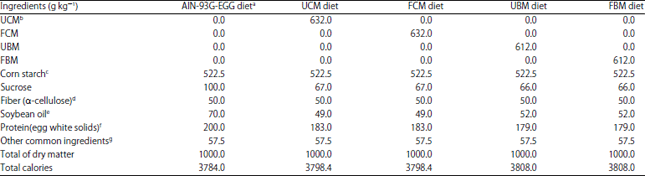 | |
FBM: Fermented bovine milk, FCM: Fermented camel milk, UBM: Unfermented bovine milk, UCM: Unfermented camel milk. aControl diet is based on the American Institute of Nutrition diet for experimental animals for growth (AIN-93G) with the needed modifications when casein is replaced with egg white solids as the protein source (AIN-93G-EGG) as described by Reeves27, bModifications in dietary components for camel milk and bovine milk diets were made according to Food Composition Tables for Arab Gulf Countries24. Both milk types contribute to 10% of the energy content of their corresponding experimental diets, cCornstarch amount includes: 390.5 g recommended in AIN-93G-EGG diet and extra 132 g used as substitution for the dextrinized cornstarch because it is usually added to facilitate pelleting while no pelleting was done here, dα-Cellulose from Sigma Chemicals Co. (St. Louis, Missouri, USA), eCrude refined soybean oil without added vitamins, fEgg white solids (spray dried) from (Bouwhuis Enthoven, Raalte, Holland), assuming that it contains 83% protein (200 g egg white solids provides approximately 166 g protein), gCommon ingredients include: Mineral mixture (AIN-93G-EGG-MX), 35 g; vitamin mixture (AIN-93G-VX), 10 g; Biotin premix, 10 g; Choline bitartrate, 2.5 g; Tert-butylhydroquinone (TBHQ), 14 mg | |
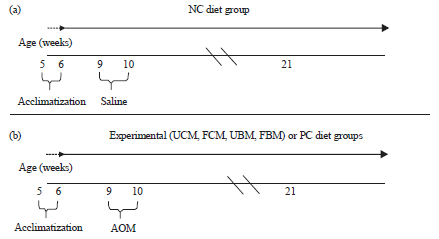 | |
| Fig. 1(a-b): | Schematic representation of the study protocol, (a) Negative control diet group (NC) and (b) Experimental diet groups include AOM carcinogen-injected groups fed with lactose-hydrolyzed milk diets: Unfermented camel milk (UCM) diet, fermented camel milk (FCM) diet, unfermented bovine milk (UBM) diet, fermented bovine milk (FBM) diet and positive control (PC) diet |
Study protocol: The initiation protocol for cancer chemoprevention was as follows: Rats in both the PC and NC groups were fed the standard AIN diet, whereas the experimental groups were fed the AIN diet supplemented with the milk preparations. All groups were fed their diets 3 weeks prior to AOM carcinogen administration, throughout the week of carcinogen or saline injections and 11 weeks after the last injection. All rats in the experimental groups and PC group received the AOM carcinogen (Sigma Chemicals Co., St Louis, Missouri, USA), which was dissolved in normal saline solution and administered at a concentration of 15 mg kg–1 body weight subcutaneously in the neck region29, whereas, the rats in the NC group were administered normal saline without the AOM carcinogen. AOM was administered to rats in two consecutive doses at 9 and 10 weeks of age. The schematic representation of the study protocol is shown in Fig. 1.
Body weight and food consumption were monitored once and twice a week, respectively, until the termination of the experiment at 11 weeks after the second AOM injection. At termination, the rats were euthanized and a necropsy was carefully performed. The colons were harvested and preserved in a 10% buffered formalin solution (Sigma Chemicals Co., St Louis, Missouri, USA). The colons were then used for the ACF counting tests according to the method described by Bird19.
The formalin-fixed colonic tissues were cut into 4 cm segments and stained in 0.2% methylene blue solution for 3-5 min. The total number of ACF and the aberrant crypts (ACs) in each focus were counted under a microscope (40-100X). ACF were identified by the following morphological characteristics: (i) Enlarged and elevated crypts compared with the normal mucosa and (ii) An increased pericryptal space and irregular lumens (as described by McLellan and Bird30 and McLellan et al.31). Crypt multiplicity (number of ACs per focus) was calculated after ACF counting. Most of the ACF were harvested for the histological analyses. Methylene blue-stained colonic tissues were examined under a microscope and large ACF containing four or more ACs were marked. Atleast 10-12 pieces of colonic tissues containing large ACF were dissected from the mid-distal part of each colon, where most of the ACF occurred. These tissues were then processed and appropriately oriented in paraffin blocks for longitudinal sectioning.
Immunostaining for β-catenin: Colon tissue samples were fixed in 10% phosphate-buffered formalin for 10 h at 4°C, dehydrated in ascending concentrations of ethanol, cleared with xylene and embedded in PolyFin (Triangle Biomedical Sciences, Durham, NC). Paraffin-embedded tissue blocks were cut with a rotary microtome into 4 μm sections mounted on positively charged slides. Standard and published procedures were used in the current study for immunostaining for β-catenin32,33.
The sections were incubated with antibodies against β-catenin clone 14/β-catenin (BD Transduction laboratories) at a 1:100 dilution in antibody diluent (Dako, Glostrup, Denmark) in a humidity chamber overnight at 4°C. The specific immunoreactivity was visualized using 3, 3-Diaminobenzidine (DAB) for 10 min at room temperature. The sections were counter stained with Mayer’s hematoxylin, dehydrated and mounted. The immunohistochemical quantification for nuclear or non-nuclear stains for β-catenin positive cells was performed in a blinded manner and the respective positivity was determined for the brown-stained cells beyond the background-level immunoreactivity in each case. For each sample, five random fields involving crypts were analyzed to quantify the staining as the percent positive cells by counting the brown-stained cells among the total number of cells. The percent positivity was determined as the number of brown positive cells×100/total number of cells. The threshold for considering a case as nuclear was 10%34.
TUNEL staining for in situ apoptotic cells: Apoptotic cells were detected using the Dead End Colorimetric TUNEL system (Promega, Madison, WI, USA) following the manufacturer’s protocol. The sections were observed under 400X magnification for TUNEL-positive cells (brown color). The apoptotic index was calculated as the number of apoptotic cells×100 /total number of cells34.
Statistical analysis: The statistical analysis was conducted using the Statistical Analysis System (SAS, version 9.1, SAS Institute Inc., Cary, NC, USA). Variables were analyzed by a one-way analysis of variance (ANOVA). Fisher’s (F-test) protected least significant difference (LSD) test was used to analyze the means and detect significant differences between the treatment means. Differences were considered significant at p<0.05. The data were presented as the least square means±standard error of the mean.
RESULTS
During the experimental period, fluctuations occurred in the food intake of the rat groups, however, the average weight gain throughout the study did not differ significantly among the experimental diet groups (Table 2). Figure 2 shows the different patterns of weight gain among the different experimental and control groups.
ACF were barely observed in the colons of rats of the NC group, whereas ACF with different counts and multiplicities were observed in the carcinogen (AOM)-treated groups. Table 3 shows the ACF types in the colons of rats fed different experimental diets. The number of small or low multiplicity foci with one, two or three ACs (1 AC, 2 AC, 3 AC, respectively) in the colons of the rats fed the different milk diets (UCM, FCM, UBM and FBM) were significantly lower than those of the PC group (p<0.05) and significantly higher than those of the NC group (p<0.05).
| Table 2: | Average weight gain and food intakes for experimental animals during the study period |
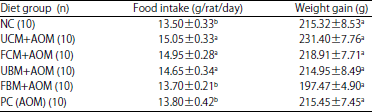 | |
FBM: Fermented bovine milk, FCM: Fermented camel milk, NC: Negative control, PC: Positive control, UBM: Unfermented bovine milk, UCM: Unfermented camel milk, n: Number of rats in each group, Values are least square means±standard error of mean. a-dMeans with different superscripts within each column are significantly different (p<0.05) by Fisher’s protected least significant differences test (PLSD) | |
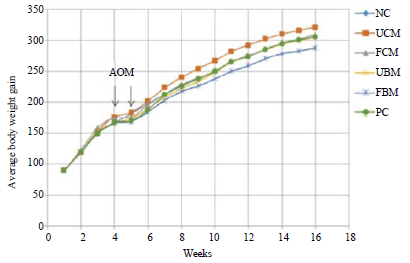 | |
| Fig. 2: | Effect of different control and experimental diets on average body weight change for rats over the 16 weeks. Experimental diet groups include azoxymethane (AOM) carcinogen-injected groups fed with lactose-hydrolyzed milk diets: FBM, Fermented bovine milk, FCM: Fermented camel milk, NC: Negative control, PC: Positive control, UBM: Unfermented bovine milk, UCM: Unfermented camel milk |
| Table 3: | Average counts of different ACF types, total AC, total ACF and CM in colons of rats fed different control and experimental diets |
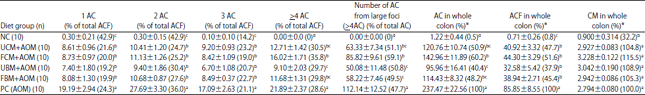 | |
AC: Aberrant crypts, ACF: Aberrant crypt foci, AOM: Azoxymethane, CM: Crypt multiplicity, FBM: Fermented bovine milk, FCM: Fermented camel milk, NC: Negative control, PC: Positive control, UBM: Unfermented bovine milk, UCM: Unfermented camel milk. n: Number of rats involved in the study group. Values are least square Mean±Standard error of mean. a-dMeans with different superscripts within each column are significantly different (p<0.05) by Fisher’s protected least significant differences test (PLSD).*Percentages were calculated relative to positive control group | |
However, the four milk diet groups exhibited insignificantly different behavior regarding ACF formation, with unfermented bovine milk exhibiting the lowest values for different AC and ACF markers. ACF with high crypt multiplicity (number of ACs/focus, >4 AC/focus) behaved similarly in all groups except the FCM and UBM groups, which were significantly different. Moreover, the number of AC from large foci in the rats fed the different milk diets was significantly lower than that of the rats of the PC group (p<0.05).
The AC counts in the whole colons of the experimental and control groups are shown in Table 3. The four milk diet groups were not significantly different from each other. Furthermore, the number of ACs in whole colons from the UBM group was (p<0.05) lower than that in the FCM group. However, the counts in the whole colons in the UBM and FCM groups were not significantly different from the other milk diet groups (UCM and FBM).
The total ACF in the whole colons of the rats fed the different experimental diets is shown in Table 3. The total ACF number in the whole colons of the four milk diet treatments were lower than that in the PC group (p<0.05). However, the number of ACF was not significantly different among the four milk diet groups in the whole colons (Table 3).
The crypt multiplicity (CM) in the colons of the four milk diet groups was not significantly different from that of the PC group (Table 3). All of the examined specimens except one showed membranous β-catenin because the percentage of nuclear β-catenin was 2% below the cutoff threshold. The specimen of PC (without chemopreventive material) that showed nuclear β-catenin was an adenocarcinoma with 20% nuclear positivity (Fig. 3).
For the apoptosis index (TUNEL), camel milk did not show any significant difference compared with the PC.
DISCUSSION
The total number of ACF (85.85±8.55) in the PC group (Table 3) was consistent with that observed in other studies that counted ACF within 10-12 weeks after the final AOM injection. The results of the current study are within the range of other reported studies, however, Sengupta et al.35 reported a lower ACF number (29.6±4.03) and Rao et al.36 reported a higher number (114±10). Such variations might be attributed to different rat ages, control diet compositions and experimental protocols.
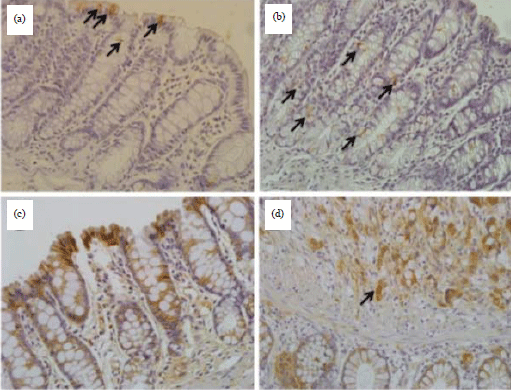 | |
| Fig. 3: | Detection of apoptosis and β-catenin subcellular localization, (a, b) Two representative examples of the average level of apoptotic figures (arrows) observed in this study using TUNEL assay for apoptosis detection, (c) Immunohistochemical staining for β-catenin showing membranous localization in the colon mucosal epithelial cells and (d) β-catenin immunohistochemistry showing nuclear localization (arrow) in a case adenocarcinoma in PC group. The overall percentage was estimated to be 20% in this case. DAB chromogen, Mayer’s hematoxylin counterstaining, original magnification 400X |
Another important criterion for understanding the early stages of colon carcinogenesis is the differential growth dynamics of ACF in different regions of the colon37. The colons of rats fed different milk diets had a significantly (p<0.05) lower average number of AC subtypes and lower total AC and ACF in the whole colons (Table 3) and the proximal and distal colon sections (data not shown) compared with the PC group, indicating that all of the milk treatments may provide chemopreventive activity against early carcinogenesis.
Although discrepancies were observed between the ACF assay and colon tumor development in the animal assays, the overall consistency was quite high. The number of "large" ACF (four or more crypts/focus) or those with high CM demonstrated a better association with tumor development than the total number of ACF31,38. Hence, these dysplastic or hyperproliferative ACF are considered more accurate predictors of subsequent tumorigenicity than smaller ones when testing the efficacy of chemopreventive agents29.
In the current study, rats fed different milk diets had 26.8-58.4% fewer large ACF (>4 AC) than the rats in the PC group (Table 3), indicating an intense chemopreventive potential. For the unfermented milk diets, the lactose-hydrolyzed bovine milk diet showed a striking 58.4% reduction in large ACF (>4 AC) compared with the PC, whereas the rats fed the unfermented camel milk diet only showed a 42% reduction. Moreover, the fermented bovine milk diets generated a 46.7% reduction in large ACF, whereas the fermented camel milk diet only generated a 26.8% reduction, indicating a higher chemoprevention effectiveness for unfermented milk compared with fermented milk.
Large ACF have been shown to accurately indicate the preneoplastic potential, thus, the results suggest that milk, particularly bovine milk, may act by retarding the progression of early ACs. This preferred effect is elucidated by the substantial reduction in multicrypt foci (>4 AC) by the different milk treatments and the ability of these diets to considerably reduce the number of ACs derived from large foci (Table 3). These crypts contributed to 47.7-59.1% of the total AC count for the different experimental groups, thus, the statistical analysis of their counts followed a similar trend to that described for the number of large ACF.
At this stage of carcinogenesis, the percentage of ACF from large crypts in the AOM-treated groups ranged between 28.6 and 35.8%, which was comparable to that reported by other researchers Rao et al.39, Reddy et al.40, Rowland et al.41 and Verghese et al.42. Such a narrow range of variations in the large ACF contribution to the total ACF count led to the lack of significant differences in crypt multiplicity (number of ACs/focus) among the different experimental groups (Table 3).
The chemopreventive effect of fermented dairy products that contain live lactic acid bacteria along with their metabolites have been shown to modulate the immune response in animals, suppress carcinogenesis in rodents, inhibit the activity of enzymes related to carcinogenesis and bind carcinogenic and mutagenic chemical compounds43,44. Further, conjugated linoleic acid found in ruminant milk inhibits the growth of several human cancer cell lines and has been found to suppress chemically induced tumor development at several sites in animal models6. Moreover, investigations have demonstrated a protective role for bovine milk proteins against tumor development and showed that both whey and casein proteins were more protective against the development of intestinal cancers in rats than other dietary proteins5. Studies on whey protein demonstrated that the major component bovine lactoferrin inhibits colon carcinogenesis in rats treated with AOM8.
According to the available literature, studies have not previously compared the anticarcinogenic effects of fermented camel and bovine milk. However, Abu-Tarboush45 investigated the growth and proteolytic activities of these milks, which were pasteurized and incubated with yogurt starter cultures as single and mixed cultures. Reddy et al.40 and Abu-Tarboush45 demonstrated the growth of four strains of Streptococcus thermophilus and three strains of Lactobacillus delbrueckii sp. bulgaricus. Proteolysis of milk proteins is known to release bioactive peptides that might have antihypertensive, antithrombotic, immunomodulating and anticarcinogenic-effects46,41. Consequently, differences in the proteolytic activities of different types of milk might have contributed to the variations in anticarcinogenic effects observed in the current study, although the differences were not significant.
In the current study, only 10% of different milk preparations were added to the experimental diets and unfermented bovine and camel milk showed a 62.1 and 52.3% reduction in the total ACF count, respectively, compared with the PC group. This reduction was attributed to the endogenous enzymes of the milk itself and the production of bioactive peptides by the enzymatic activities of the contaminating bacteria in the raw milk. These results are consistent with those of Tavan et al.47 for unfermented bovine milk (66% reduction). However, Narushima et al.48 reported that the incorporation of 10% lyophilized unfermented bovine milk into the diet of F344 rats led to a 19.2% reduction in the total number of ACF induced by a heterocyclic amine.
Compared with the PC group, the rat groups fed FBM and FCM showed a 54.6 and 48.4% reduction in the total ACF count, respectively, which could have been caused by a further breakdown of active peptides responsible for the anticarcinogenic effect. A higher reduction (93-96%) in the ACF count was reported by Tavan et al.47 using a fermented bovine milk diet. The discrepancies in these results may have been caused by the following factors: first, a lower number of ACF (7.3±4.2) were induced by the heterocyclic aromatic amines in the PC group of Tavan’s study compared with that induced by the AOM in the current study (85.85±8.55), second, fermented milk preparations contributed to only 10% of the energy content of the experimental diets in the current study compared with 30% in Tavan’s study, third, different bacterial cultures were used in both studies and fourth, 3% of the bacterial culture was inoculated into the pasteurized milk in Tavan’s study, whereas only 2% of the culture was added in the current study47.
The reduction in the total number of ACF reported in the current study by both types of fermented milk (54.6% and 48.4%) were similar to those reported by Narushima et al.48, who incorporated lyophilized yogurt powder at a concentration of 10% into the diet of F344 rats and observed a 45.7% reduction in the total number of ACF compared with that in the positive control. Narushima et al.48 used an equal ratio of both Lactobacillus delbrueckii sp. bulgaricus and Streptococcus thermophilus and inoculated them in milk at a concentration of 2%. This protocol was closer to the current study protocol compared with that followed Tavan et al.47 However, both studies used different types of heterocyclic amines as carcinogens, which may have caused variations from the current study.
In the current study, the early stage of carcinogenesis was not sufficient to exhibit the molecular changes in β-catenin and the apoptotic index because these markers are exhibited at more advanced stages of carcinogenesis. In another study Xiao et al.49, increased apoptosis and decreased nuclear expression levels of β-catenin were reported in experimental animals at 34 weeks after the last injection. In the current study, the presence of membranous β-catenin rather than nuclear or cytoplasmic β-catenin in all examined colon samples along with the lack of significant differences in apoptosis between the camel milk and PC groups were consistent with the lack of significant differences in CM between the milk treatments and PC group (Table 3). β-catenin has been observed at the cell membrane in ACF with hyperplasia, which is similar to that in normal colon epithelium, however, in ACF with dysplasia as well as in adenoma and carcinoma, β-catenin is localized less in the membrane and more in the nuclei and cytoplasm50. Furthermore, the observed results were consistent with a lack of severe dysplasia, adenoma and adenocarcinoma in all milk-treated groups (data not shown). All these results are consistent with early carcinogenesis (as expressed by suppressed ACF) and not encompassing dramatic cytological and molecular changes, which indicates that such changes are shown at later stages of carcinogenesis.
This is the first in vivo study to examine the chemopreventive effects of camel milk against early colon carcinogenesis induced in an animal model that mimics human sporadic CRC, thus, the findings are only preliminary. Moreover, a number of limitations were observed, thus, further investigations are required. The chemopreventive ability of camel milk was studied in the early stages of carcinogenesis using ACF precancerous lesions, which may have been insufficient for demonstrating the chemopreventive effects of colon cancer or fully elucidating the underlying mechanisms. Further investigations of the chemopreventive ability of camel milk against later stages of carcinogenesis may clarify the chemopreventive effects of milk, although a study period of more than 30 weeks will be required.
CONCLUSION
The results of the current research showed that pasteurized camel and bovine milk exhibit chemopreventive potential against preneoplastic lesions as expressed by the ACF counts in the early stages of colon carcinogenesis. Moreover, fermenting both types of milk did not significantly improve the chemopreventive potential. The current study failed to elucidate the molecular mechanisms underlying the chemopreventive potential of the tested milk at the early stages of carcinogenesis.
ACKNOWLEDGMENTS
The authors would like to thank "The Deanship of Academic Research" at the University of Jordan and the University of Sharjah for funding this research through Grant No. 22/2/2015. This study was also supported by the Environment and Cancer Research Group, Sharjah Institute for Medical Research (SIMR), University of Sharjah. The authors would also like to thank Vidhya Anish Nair and Harini Sampith Kumar for providing technical assistance.
REFERENCES
- Schoen, R.E., P.F. Pinsky, J.L. Weissfeld, L.A. Yokochi and T. Church et al., 2012. Colorectal-cancer incidence and mortality with screening flexible sigmoidoscopy. New Engl. J. Med., 366: 2345-2357.
Direct Link - Ahmed, F.E., 2004. Effect of diet, life style and other environmental/chemopreventive factors on colorectal cancer development and assessment of the risks. J. Environ. Sci. Health, Part C, 22: 91-148.
PubMedDirect Link - De Angelis, R.C., I.C.M. Terra and J.V.M. Campos, 1988. The effect of maturation and source of dietary protein on the capacity of the small intestine to hydrolize lactose in rats. Nutr. Res., 8: 1413-1420.
CrossRefDirect Link - Gill, H.S. and M. Cross, 2000. Anticancer properties of bovine milk. Br. J. Nutr., 84: 161-164.
Direct Link - Parodi, P.W., 1999. Conjugated linoleic acid and other anticarcinogenic agents of bovine milk fat. J. Dairy Sci., 82: 1339-1349.
CrossRefDirect Link - Tsuda, H. and K. Sekine, 2000. Milk components as cancer chemopreventive agents. Asian Pac. J. Cancer Prev., 1: 277-282.
PubMed - Tsuda, H., K. Sekine, Y. Ushida, T. Kuhara, N. Takasuka, M. Iigo, B.S. Han and M.A. Moore, 2000. Milk and dairy products in cancer prevention: Focus on bovine lactoferrin. Mutat. Res., 462: 227-233.
CrossRefDirect Link - Shwter, A.N., N.A. Abdullah, M.A. Alshawsh, H.R. El-Seedi and N.A. Al-Henhena et al., 2016. Chemopreventive effect of Phaleria macrocarpa on colorectal cancer aberrant crypt foci in vivo. J. Ethnopharmacol., 193: 195-206.
CrossRefDirect Link - Al Haj, O.A. and H.A. Al Kanhal, 2010. Compositional, technological and nutritional aspects of dromedary camel milk. Int. Dairy J., 20: 811-821.
CrossRefDirect Link - Khedid, K., M. Faid, A. Mokhtari, A. Soulaymani and A. Zinedine, 2009. Characterization of lactic acid bacteria isolated from the one humped camel milk produced in Morocco. Microbiol. Res., 164: 81-91.
CrossRefDirect Link - Yateem, A., M.T. Balba, T. Al-Surrayai, B. Al-Mutairi and R. Al-Daher, 2008. Isolation of lactic acid bacteria with probiotic potential from camel milk. Int. J. Dairy Sci., 3: 194-199.
CrossRefDirect Link - Salami, M., A.A. Moosavi-Movahedi, F. Moosavi-Movahedi, M.R. Ehsani and R. Yousefi et al., 2011. Biological activity of camel milk casein following enzymatic digestion. J. Dairy Res., 78: 471-478.
Direct Link - Hamad, E.M., E.A. Abdel-Rahim and E.A. Romeih, 2011. Beneficial effect of camel milk on liver and kidneys function in diabetic sprague-dawley rats. Int. J. Dairy Sci., 6: 190-197.
CrossRefDirect Link - Dallak, M., 2009. Camel's milk protects against cadmium chloride-induced hypochromic microcytic anemia and oxidative stress in red blood cells of white albino rats. Am. J. Pharmacol. Toxicol., 4: 136-143.
Direct Link - Salwa, M.Q. and A.F.K. Lina, 2010. Antigenotoxic and anticytotoxic effect of camel milk in mice treated with cisplatin. Saudi J. Biol. Sci., 17: 159-166.
CrossRefDirect Link - Sohn, O.S., F.S. Emerich, S.P. Requeijo, J.H. Weisburger and F.J. Gonzalez, 2001. Differential effects of CYP2E1 status on the metabolic activation of the colon carcinogens azoxymethane and methylazoxymethanol. Cancer Res., 61: 8435-8440.
PubMedDirect Link - Femia, A.P., C. Luceri, P. Dolara, A. Giannini and A. Biggeri et al., 2002. Antitumorigenic activity of the prebiotic inulin enriched with oligofructose in combination with the probiotics Lactobacillus rhamnosus and Bifidobacterium lactis on azoxymethane-induced colon carcinogenesis in rats. Carcinogenesis, 23: 1953-1960.
CrossRefPubMedDirect Link - Bird, R.P., 1987. Observation and quantification of aberrant crypts in the murine colon treated with a colon carcinogen: Preliminary findings. Cancer Lett., 37: 147-151.
PubMedDirect Link - Stevens, R., H. Swede and D. Rosenberg, 2007. Epidemiology of colonic aberrant crypt foci: Review and analysis of existing studies. Cancer Lett., 252: 171-183.
CrossRefPubMedDirect Link - Faris, M.A.I.E., H.R. Takruri, M.S. Shomaf and Y.K. Bustanji, 2009. Chemopreventive effect of raw and cooked lentils (Lens culinaris L.) and soybeans (Glycine max) against azoxymethane-induced aberrant crypt foci. Nutr. Res., 29: 355-362.
CrossRefDirect Link - Sekine, K., Y. Ushida, T. Kuhara, M. Iigo and H. Baba-Toriyama et al., 1997. Inhibition of initiation and early stage development of aberrant crypt foci and enhanced natural killer activity in male rats administered bovine lactoferrin concomitantly with azoxymethane. Cancer Lett., 121: 211-216.
CrossRefDirect Link - Boone, C.W., V.E. Steele and G.J. Kelloff, 1992. Screening for chemopreventive (anticarcinogenic) compounds in rodents. Mutat. Res./Fundam. Mol. Mechan. Mutag., 267: 251-255.
Direct Link - Chen, J. and X.F. Huang, 2009. The signal pathways in azoxymethane-induced colon cancer and preventive implications. Cancer Biol. Ther., 8: 1313-1317.
Direct Link - Khan, N., F. Afaq and H. Mukhtar, 2006. Apoptosis by dietary factors: The suicide solution for delaying cancer growth. Carcinogenesis, 28: 233-239.
PubMedDirect Link - Jelen, P. and O. Tossavainen, 2003. Low lactose and lactose-free milk and dairy products-prospects, technologies and applications. Aust. J. Dairy Technol., 58: 161-165.
Direct Link - Reeves, P.G., 1997. Components of the AIN-93 diets as improvements in the AIN-76A diet. J. Nutr., 127: 838S-841S.
PubMedDirect Link - Pereira, M.A., L.H. Barnes, V.L. Rassman, G. Kelloff and V. Steele, 1994. Use of azoxymethane-induced foci of aberrant crypts in rat colon to identify potential cancer chemopreventive agents. Carcinogenesis, 15: 1049-1054.
PubMedDirect Link - McLellan, E.A. and R.P. Bird, 1988. Aberrant crypts: Potential preneoplastic lesions in the murine colon. Cancer Res., 48: 6187-6192.
PubMed - McLellan, E., A. Medline and R. Bird, 1991. Sequential analyses of the growth and morphological characteristics of aberrant crypt foci: Putative preneoplastic lesions. Cancer Res., 51: 5270-5274.
PubMed - Abdel-Rahman, W.M., 2006. Mutations of beta-catenin and KRAS in colorectal carcinomas. Digest. Dis. Sci., 51: 616-617.
PubMedDirect Link - Abdel-Rahman, W.M., M. Ollikainen, R. Kariola, H.J. Jarvinen and J.P. Mecklin et al., 2005. Comprehensive characterization of HNPCC-related colorectal cancers reveals striking molecular features in families with no germline mismatch repair gene mutations. Oncogene, 24: 1542-1551.
PubMedDirect Link - Velmurugan, B., R.P. Singh, R. Agarwal and C. Agarwal, 2010. Dietary‐feeding of grape seed extract prevents azoxymethane‐induced colonic aberrant crypt foci formation in fischer 344 rats. Mol. Carcinog., 49: 641-652.
CrossRefDirect Link - Sengupta, A., S. Ghosh and S. Das, 2003. Tea can protect against aberrant crypt foci formation during azoxymethane induced rat colon carcinogenesis. J. Exp. Clin. Cancer Res., 22: 181-191.
PubMedDirect Link - Rao, C.V., T. Kawamori, R. Hamid and B.S. Reddy, 1999. Chemoprevention of colonic aberrant crypt foci by an inducible nitric oxide synthase-selective inhibitor. Carcinogenesis, 20: 641-644.
CrossRefDirect Link - Magnuson, B.A., I. Carr and R.P. Bird, 1993. Ability of aberrant crypt foci characteristics to predict colonic tumor incidence in rats fed cholic acid. Cancer Res., 53: 4499-4504.
PubMed - Pretlow, T.P., M.A. O`Riordan, G.A. Sonich, S.B. Amini and T.G. Pretlow, 1992. Aberrant crypts correlate with tumor incidence in F344 rats treated with azoxymethane and phytate. Carcinogenesis, 13: 1509-1512.
CrossRefPubMedDirect Link - Rao, C.V., B. Simi and B.S. Reddy, 1993. Inhibition by dietary curcumin of azoxymethane-induced ornithine decarboxylase, tyrosine protein kinase, arachidonic acid metabolism and aberrant crypt foci formation in the rat colon. Carcinogenesis, 14: 2219-2225.
Direct Link - Reddy, B.S., C.V. Rao and K. Seibert, 1996. Evaluation of cyclooxygenase-2 inhibitor for potential chemopreventive properties in colon carcinogenesis. Cancer Res., 56: 4566-4569.
Direct Link - Rowland, I.R., C.J. Rumney, J.T.Coutts and L.C. Lievense, 1998. Effect of bifidobacterium longum and inulin on gut bacterial metabolism and carcinogen-induced aberrant crypt foci in rats. Carcinogenesis, 19: 281-285.
PubMedDirect Link - Verghese, M., D.R. Rao, C.B. Chawan, L.L. Williams and L. Shackelford, 2002. Dietary inulin suppresses azoxymethane-induced aberrant crypt foci and colon tumors at the promotion stage in young fisher 344 rats. J. Nutr., 132: 2809-2813.
PubMedDirect Link - Davoodi, H., S. Esmaeili and A. Mortazavian, 2013. Effects of milk and milk products consumption on cancer: A review. Comprehens. Rev. Food Sci. Food Safety, 12: 249-264.
CrossRefDirect Link - Perdigon, G., A.M. de LeBlanc, J. Valdez and M. Rachid, 2002. Role of yoghurt in the prevention of colon cancer. Eur. J. Clin. Nutr., 56: S65-S68.
PubMedDirect Link - Abu-Tarboush, H., 1996. Comparison of associative growth and proteolytic activity of yogurt starters in whole milk from camels and cows. J. Dairy Sci., 79: 366-371.
CrossRefDirect Link - Dziuba, M., B. Dziuba and A. Iwaniak, 2009. Milk proteins as precursors of bioactive peptides. Acta Sci. Pol Technol. Aliment, 8: 71-90.
Direct Link - Tavan, E., C. Cayuela, J.M. Anroine, G. Trugnan, C. Chaugier and P. Cassand, 2002. Effects of diary products on heterocyclic aromatic amine-induced rat colon carcinogenesis. Carcinogenesis, 23: 477-483.
Direct Link - Narushima, S., T. Sakata, K. Hioki, T. Itoh, T. Nomura and K. Itoh, 2010. Inhibitory effect of yogurt on aberrant crypt foci formation in the rat colon and colorectal tumorigenesis in RasH2 mice. Exp. Anim., 59: 487-494.
PubMedDirect Link - Xiao, H., X. Hao, B. Simi, J. Ju, H. Jiang, B.S. Reddy and C.S. Yang, 2008. Green tea polyphenols inhibit colorectal Aberrant Crypt Foci (ACF) formation and prevent oncogenic changes in dysplastic ACF in azoxymethane-treated F344 rats. Carcinogenesis, 29: 113-119.
CrossRefPubMedDirect Link - Hao, X.P., T.G. Pretlow, J.S. Rao and T.P. Pretlow, 2001. β-Catenin expression is altered in human colonic aberrant crypt foci. Cancer Res., 61: 8085-8088.
PubMed