
Mairizal
Department of Poultry Nutrition, Faculty of Animal Science, University of Jambi, Jambi, Indonesia
Yetti Marlida
Department of Animal Nutrition and Feed Technology, Faculty of Animal Science, University of Andalas, Padang, Indonesia
Mirzah
Department of Animal Nutrition and Feed Technology, Faculty of Animal Science, University of Andalas, Padang, Indonesia
Fahmida Manin
Department of Animal Science, Faculty of Animal Science, University of Jambi, Jambi, Indonesia
Pakistan Journal of Nutrition
Year: 2018 | Volume: 17 | Issue: 3 | Page No.: 116-123







ABSTRACT
Background and Objective: Mannanase is a mannan-degrading enzyme that can be produced by microorganisms such as bacteria. Mannanase is widely used in the animal feed industry with the aim of improving the quality of feed ingredients containing high levels of mannan, such as palm kernel meal. This study aimed to isolate, characterize and produce mannanase from bacteria found in the hindgut of termites. Materials and Methods: Bacteria were isolated by culturing on solid media containing 0.5% locust bean gum for 24 h at 30°C. The mannanolytic index was then measured by the addition of 2% congo red, which was then washed with 1 M NaCl. Mannanase activity was measured by the dinitrosalicylic acid method, while the protein content was measured by the Bradford method. Results: A total of 8 isolates were obtained. The 3 isolates with the highest mannanolytic indices were selected (isolates V9, AZ and AD). Isolate V9 exhibited the highest mannanase activity of 29.50 U mL–1 after 88 h of incubation with a specific activity of 110.32 U mg–1. The activity of the mannanase was optimal at pH 8 and at an incubation temperature of 50°C. Based on 16S-DNA analysis, isolate V9 was identified as Bacillus cereus. Conclusion: Bacillus cereus, which is isolated from the hindgut of termites, has fairly high mannanase activity and has the potential to hydrolyse mannan in feed ingredients such as palm kernel meal.
PDF Abstract XML References Citation
Received: September 25, 2017;
Accepted: January 03, 2018;
Published: February 15, 2018
Copyright: © 2018. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Mairizal, Yetti Marlida, Mirzah and Fahmida Manin, 2018. Isolation and Characterization of Mannanase-producing Bacillus cereus Isolated from the Hindgut of Termites. Pakistan Journal of Nutrition, 17: 116-123.
DOI: 10.3923/pjn.2018.116.123
URL: https://scialert.net/abstract/?doi=pjn.2018.116.123
DOI: 10.3923/pjn.2018.116.123
URL: https://scialert.net/abstract/?doi=pjn.2018.116.123
INTRODUCTION
Mannan is part of the hemicellulose content of plant cell walls and is widely distributed in nature as part of the hemicellulose fraction of hardwoods and softwoods as palm kernel meal (PKM). PKM contains a moderate level of crude protein (15.5-19.6%), a high level of fibre (13-20%) and a poor amino acid profile (PKM is deficient in lysine, methionine and tryptophan) and thus, PKM is considered to be a moderate quality feed ingredient for ruminants but is not suitable for monogastric animals1. The use of PKM for monogastric animals, such as poultry is limited because these animals lack the appropriate enzymes to hydrolyse the fibre. PKM contains high levels of fibre with non-starch polysaccharides (NSP) and β-mannan as the main fibre components, 78% of the total NSP in PKM is linear mannan2,3. Therefore, mannanase supplementation into PKM will result in the degradation of the mannan component, thereby increasing its suitability for use in poultry rations.
Mannanase is an enzyme that degrades mannan, glucomannan and galactomannan to mannose, glucose and galactose, respectively. Mannanase participates in the degradation of hemicellulose and similar polysaccharides by hydrolysing the β-1,4-glycosidic linkages within the main chain4. This enzyme can be produced from microorganisms, animals and plants. The main source of β-mannanase is microbial and it has been reported that β-mannanase is present in bacteria such as Bacillus subtilis TJ-1025, Bacillus circulans NT6.76, Bacillus sp.7, Bacillus pumilus (M27)8, Bacillus amyloliquefaciens CS479 and Bacillus licheniformis10. One microorganism that has the ability of producing this enzyme is a bacterium found in the hindgut of termites.
Termites are insects that use wood or timber as their main source of food. They attack trees and plants and are therefore, categorized as pests that are potentially harmful to humans. Termites play an important role in the recycling of plant nutrients through a process of disintegration and decomposition of organic material from wood and plant biomass into organic material. This process is caused by the presence of microbes in the hindgut of termites that can produce cellulase and lignin11. Termites have a dense and diverse population of symbiotic microflora in their gut that can degrade cellulose in wood, this cellulose is first partially degraded in the foregut of the termite by its own enzyme12. The digestive tract of termites consists of several parts such as the foregut, midgut and hindgut. Wood-feeding termites live solely on dead plant materials and their xylophagy depends on digestive symbiosis in the hindgut, which houses both eukaryotes (protozoa and fungi) and prokaryotes (bacteria and archaea)13.
Cellulolytic bacteria such as Acinetobacter, Pseudomonas, Staphylococcus and various species of the Enterobacteriaceae and Bacillaceae families have been reported to be present in the termite gut14. Several Bacillus and Paenibacillus species have been detected in the termite gut, of which the genera Bacillus are predominant with titres of up to 107 mL–1 gut contents15-17. Analysis of the extract from the termite Glyptotermes montanus shows the complex enzymatic activity of cellulases and that the activities of avicelase, β-D-1,4-mannanase, β-D-1,4-xylanase and β-D-1,4-glucosidase were very low18. The production of enzymes is easier and faster by using microbial isolates found in the hindgut of termites rather than by producing large numbers of termites for enzyme extraction. Based on this observation, the present study was conducted to isolate, characterize and produce mannanase from the bacteria found in the hindgut of termites.
MATERIALS AND METHODSM
Isolation of mannanolytic bacteria: The bacteria were isolated by the plate scratch method. Isolation of bacteria began with the sampling of the digestive tracts of termites. As many as 20 species of termite workers Microtermes sp., were anaesthetized using ice and then the mouths and anuses of the termites were smeared with paraffin and surface-sterilized using 80% ethanol to avoid contamination. The intestines were separated using a sterile surgical blade (Suzhou Texnet Co., Ltd., China) and then homogenized in 0.1% peptone by vortexing for 4 min in a sterile manner.
Isolation of bacteria was carried out on a modified spread plate19 prepared using media containing 0.5% locust bean gum (LBG), 0.075% peptone, 0.05% yeast extract and mineral mix (0.14% (NH4)2SO4, 0.2% KH2PO4, 0.03% MgSO4.7H2O, 0.03% CO (NH4)2, 0.03% CaCl2, 0.0005% FeSO4.7H2O, 0.00016% MnCl2.7H2O, 0.00014% ZnSO4.7H2O and 0.0002% COCl2). A 1 mL sample was enriched by preparing serial dilutions from 10–1 to 10–5 in 0.1% peptone. As much as 1 mL of each dilution from 10–2 to 10–5 was taken and used to inoculate 10 mL of 0.5% (LBG) liquid medium. Subsequently, the suspension was shaken for 24 h in a shaker at room temperature then streaked with a repeated scratch-quadrant method to obtain an individual colony on isolation medium consisting of 0.5% locust bean gum, mineral mix, 0.075% peptone, 0.05% yeast extract and 1.5% agar at pH 6. Isolates that can produce mannanase establish a clear zone around the colony. Observation of the clear zone was made easier by the addition of 2% congo red that was then washed with 2 M NaCl and left to stand for 15 min. This was repeated 3 times and then the formation of a clear zone was observed, the clear zone was measured. The mannanolytic index (IM) is the ratio between the diameter of the clear zone and colony diameter measured using callipers.
Mannanase production and mannanase assay: The liquid medium was composed of 0.5% locust bean gum, 0.075% peptone, 0.05% yeast extract and mineral mix. The media was divided into two tubes, one for pre-cultivation and the other for cultivation. Thirty millilitres of pre-cultivation media was taken in a 300 mL Erlenmeyer flask, while as much as 300 mL of cultivation media was taken in a 1000 mL Erlenmeyer flask. Once sterilized and cooled, as much as 3 dose of the selected bacteria was inserted into the medium aseptically and then, the flasks containing the pre-cultivation isolates were grown overnight in an incubator shaker at a speed of 150 rpm and a temperature of 30°C. The pre-cultivation phase was followed by isolate cultivation for 6 days in an incubator shaker at 150 rpm at room temperature. Five millilitre samples were extracted every 8 h for 5 days. The growth curve was prepared by taking 1 mL of the samples and measuring its absorbance at λ = 660 nm in a spectrophotometer.
Crude extract of mannanase was obtained from the cell culture by centrifugation at a speed of 12,000 rpm for 15 min at 4°C. Then, the supernatant was used to test for mannanase activity and protein content. Mannanase activity was evaluated by measuring the amount of reducing sugars produced by the 3,5-Dinitrosalicylic acid (DNS) method20, mannose was used as a standard. The mannanase activity was assayed by incubating the enzyme solution for 30 min in a substrate of 0.5% locust bean gum prepared in 50 mM of phosphate buffer at pH 6. The reaction was stopped by immersing the test tube in boiling water for 20 min. The reducing sugar obtained was assessed by spectrophotometry at a wavelength of 540 nm. One unit of mannanase activity was defined as the amount of enzyme that produces 1 μmol of mannose per minute under the assay conditions.
The protein content of the crude enzyme was measured to determine the specific activity of the enzyme using the Bradford method21, i.e., by adding 5 mL of Bradford reagent into 0.1 mL of crude enzyme, vortexing well to mix thoroughly and incubating at room temperature for 10 min. Then, the absorbance was measured at λ = 595 nm, for the blank, the enzyme was replaced with distilled water. The standard used was bovine serum albumin (BSA).
Effect of pH on enzyme activity: Determination of the optimum pH was carried out by testing the enzyme activity at pH 3-9 at room temperature. The reaction mixtures were composed of 0.5% LBG in 0.5 mL of buffer (at each of the tested pH values) and 0.5 mL of crude enzyme and incubated at room temperature for 30 min. The buffer used was 50 mM of citrate buffer at pH 3-5 (citric acid+sodium citrate), 50 mM of phosphate buffer at pH 6-8 (sodium phosphate monobasic+sodium phosphate dibasic) and 50 mM of glycine-NaOH at pH 9-10.
Effect of temperature on enzyme activity: The optimum temperature was determined by testing the enzyme activity at 30-100°C at temperature intervals of 10°C. The reaction mixtures were composed of 0.5% LBG in 0.5 mL of buffer at the optimum pH and 0.5 mL of crude enzyme and incubated for 30 min in a water bath at the various temperatures. The effects of pH and temperature on mannanase activity were measured by using the DNS method17.
16S-rRNA gene analysis: Genomic DNA was used for PCR amplification by the following 16S-rRNA-specific primers: 63f (5’CAG GCC TAA CAC ATG CAA GTC-3’) and 1387r (5’-GGG CGG WTG GTA CAA GGC-3’)22. The PCR product was used for DNA electrophoresis on a 1% agarose gel and then visualized by using EtBr staining and UV illumination. The DNA amplicon was then sequenced. Homology analysis was performed using the basic local alignment tools (BLASTN) method on the GenBank database.
RESULTS AND DISCUSSION
Isolates of mannanolytic bacteria: Isolation of bacteria from the digestive tracts of Microtermes sp., worker termites grown on 0.5% locust bean gum media is an early stage in the production of mannanase enzyme. The LBG was used as a carbon source for bacterial metabolism because it is enriched with minerals that induce mannanase enzymes. The ability of bacteria to degrade mannan can be qualitatively observed by the formation of a clear zone around the bacterial colony, which was visualized by congo red staining. It was observed that out of 8 isolates, only 3 isolates had high mannanolytic indices, i.e., isolates V9, AZ and AD, which had mannanolytic indices of 2.87, 2.58 and 1.97, respectively. Isolate V9 had the largest mannanolytic index, as shown by the clear zone formed around the colonies of this isolate, Gram-staining showed that the isolated V9 is a rod-shaped Gram-positive bacterium and has spores.
The clear zone of isolate V9 is large, but isolate V9 is not necessarily the isolate that was selected to be used as a mannanase. It is important to note that the plate screening method is not quantitative and the correlation between enzyme activity and colony-to-clear-zone ratio is considered to be weak23. Therefore, the growth curve and mannanase activity should be noted so that the isolates with the highest enzyme activity will be selected.
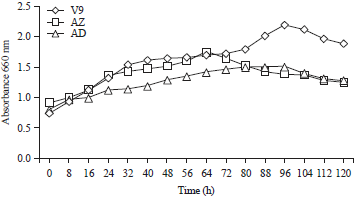 | |
| Fig. 1: | Growth curves of bacterial isolates V9, AZ and AD in liquid medium containing 0.5% LBG |
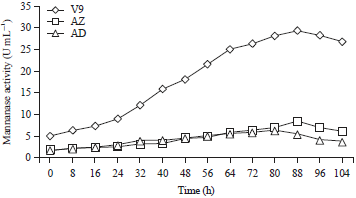 | |
| Fig. 2: | Mannanase activities of isolates V9, AZ and AD grown in specific media containing 0.5% LBG |
Growth curve: Isolates V9, AZ and AD were grown in liquid media containing 0.5% locust bean gum as a carbon source for growth, which will spur bacteria to secrete cellular metabolites such as mannanase. The growth of bacteria in liquid media containing LBG can be seen from the colour change of the media from translucent white to yellowish-white and turbid. The LBG is a galactomannan with a mannose to galactose ratio of 4:124 and can be used as a carbon source by the bacteria for growth of biomass, cell maintenance and generation of cellular products.
The growth curves of bacterial isolates V9, AZ and AD show the different growth rates, isolates V9 and AD reached a growth peak after 96 h of incubation, but the growth peak of isolate AD was lower than that of isolate V9, isolate AZ reached a growth peak after 64 h of incubation (Fig. 1). The height of the growth peak in isolate V9 indicated that the bacteria were able to regenerate well by utilizing LBG as a carbon source for growth, hence, the bacteria will also secrete mannanase to degrade mannan in high quantities.
Figure 1 shows that the adaptation phase (lag phase) in the environment is less visible than the growth curve shown above. This is because isolates V9, AZ and AD adapted to the environment during pre-cultivation. Isolate V9 is in the multiplication phase (exponential phase) of growth for 40 h from the beginning of the incubation, whereas, it is in the stationary phase from hour 48-80 post-incubation, going back to the propagation phase until 96 h post-incubation and then entering a death phase at hour 104 until hour 120. Isolates AZ and AD seem to have longer static phases than isolate V9.
Mannanase production: The mannanase activity of isolate V9 was higher than that of isolates AZ and AD (Fig. 2), hence, isolate V9 was selected as the isolate to be used for the optimal production of mannanase. Mannanase is the primary metabolite needed by the bacteria to degrade the carbon source, i.e., the mannan contained in the LBG substrate for bacterial growth.
The adaptation phase of isolate AZ lasted from 0-24 h. Then, the cells entered the propagation phase until an incubation time of 80 h and reached their optimum point at 88 h, the enzyme activity at this stage was 8.43 U mL–1. Similarly, the isolate AD reached the optimum point of mannanase production after 80 h of incubation, the enzyme activity at this stage was 6.45 U mL–1.
The mannanase activity of isolate V9 increased after the first 8 h of incubation. The mannanase activity at the beginning of the incubation was 5.12 U mL–1 at 0 h and reached 29.50 U mL–1 at 88 h, which was the optimum point for this isolate. After 96 h, the mannanase activity decreased in all isolates, which is consistent with the observation that the bacteria entered the death phase at this time. The production time of the highest mannanase activity attained was then used as a standard to determine the next highest mannanase activity.
The mannanase activity of isolate V9 was higher than the activity of the mannanase produced by Bacillus pumilus S5 (which is 0.1989 U mL–1), Bacillus megaterium25, Bacillus sp., GA2(1)26, Brevibacillus borstelensis27 and Bacillus amyloliquefaciens28.
The specific mannanase activities of the 3 isolates were consistent with the rates of mannanase activity (Fig. 3). The specific mannanase activities of the 3 isolates were up to three times higher than the measured enzyme activities. This finding was the result of low levels of total protein, leading to a high effective purity of the enzyme. The increased specific activity of the mannanase produced by isolate V9 peaked at 88 h with a value of 110.32 U mL–1, while the specific mannanase activity of isolate AZ at h 88 is 29.92 U mL–1 and that of isolate AD at h 80 is 20.84 U mL–1. The specific activity of this enzyme is higher than that of the enzymes from Saccharopolyspora29 and Streptomyces costaricaus30.
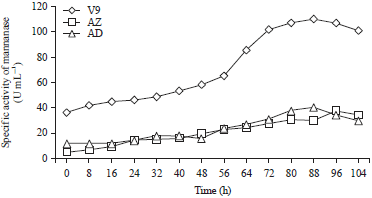 | |
| Fig. 3: | Specific activities of mannanase isolates V9, AZ and AD |
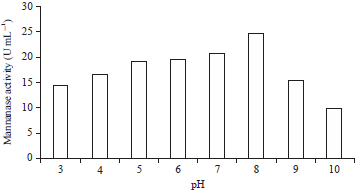 | |
| Fig. 4: | Effect of pH on the activity of mannanase from isolate V9 |
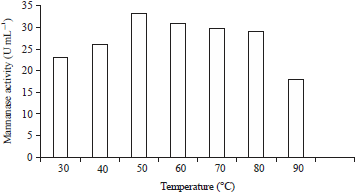 | |
| Fig. 5: | Effect of temperature on the activity of the mannanase from isolate V9 incubated at the optimal pH |
Effect of pH on enzyme activity: Isolate V9 was selected for the production of mannanase because isolate V9 was capable of producing mannanase at high quantities compared with isolates AZ and AD. Selected isolates were characterized further to determine the optimal pH and temperature for the enzyme activity and to determine the stability of mannanase at the optimal pH and temperature.
Mannanase-producing isolate V9 was affected by changes in the pH, as reflected in the value of enzyme activity, which fluctuated with changes in pH in the range 3-10. As seen in Fig. 4, the isolated V9 had the highest mannanase activity, 24.60 U mL–1, at pH 8. If the pH was too acidic or alkaline, there would be an obvious decrease in enzyme activity. When pH conditions are optimal for the bacteria, the enzymatic activity is the highest. This finding is due to changes in the ionization states of ionic groups in active sites of enzymes, which cause conformational changes in the active site to facilitate more effective in binding of the enzyme to substrates to convert them into products. The induction of morphological changes of the bacteria and secretion of the enzyme was affected by the pH conditions of bacterial growth media31. The results of this study are consistent with those reported for Bacillus nealsonii PN-1132 and Bacillus sp., HDYM-0533, which exhibit optimal mannanase production at pH 8.
Effect of temperature on enzyme activity: The optimum temperature for mannanase production was investigated by conducting an activity assay in LBG in phosphate buffer at the optimum pH (pH 8.0) at various temperatures from 30-90°C. The influence of temperature on the enzymatic activity of mannanase isolate V9 at optimum pH conditions can be seen in Fig. 5. The mannanase activity of isolate V9 was lower at a temperature of 30°C (23.23 U mL–1) and increased with an increase in temperature. The enzyme activity also increased until the optimum temperature was attained, i.e., a temperature of 50°C with an enzymatic activity of 33.24 U mL–1. Then, at temperatures from 60-90°C the activity of the enzyme decreased again. Inactivation by temperature occurs due to weak intermolecular forces affecting the stability of the three-dimensional structure of the enzyme, leading to a decrease in the catalytic capabilities of the enzyme34. The same optimum temperature for mannanase activity was also reported in Bacillus subtilis WY3435, Bacillus circulans M-2136 and Bacillus sp., GA2(1)26.
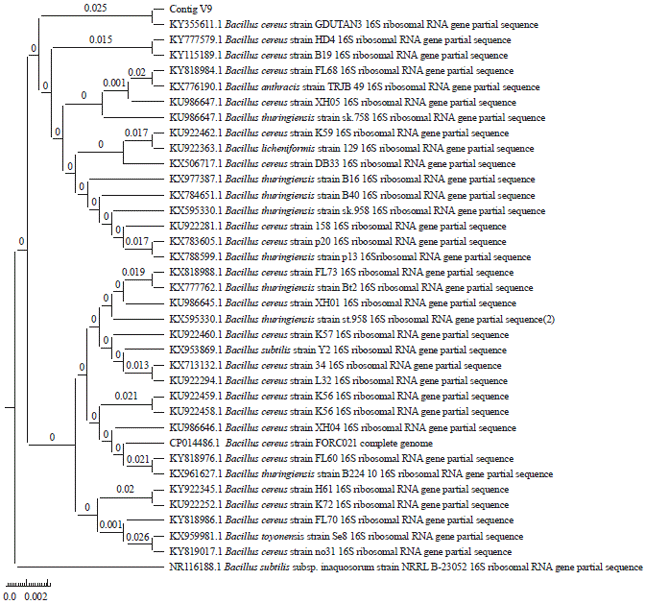 | |
| Fig. 6: | Phylogenetic tree for isolates V9 constructed using Mega 6.0 software |
Identification of isolates V9: Based on the results of partial sequencing of the 16S-rRNA gene and BLAST-N analysis, isolate V9 has proximity to Bacillus cereus at a similarity index of 99% with accession number KU 510086.1. The phylogenetic tree constructed also identifies the position of isolate V9 to be adjacent to Bacillus cereus (Fig. 6). Bacillus cereus has also been identified with similar results in Odontotermes and Heterotermes sp.,37, Mastotermes darwiniensis15, Coptotermes curvignathus38 and Amitermes evitifer39.
Bacillus cereus isolated from hindgut of termites exhibits sufficiently high mannanase activity and can be considered for the production of enzymes. Mannanase from Bacillus cereus can be used to hydrolyse feed containing high mannan, such as palm kernel meal and coconut meal, to increase the quality of the feed. Therefore, in the future, a study needs to be conducted to test the quality of feed containing high mannan after the addition of mannanase produced by Bacillus cereus.
CONCLUSION
Bacillus cereus, which is isolated from the hindgut of termites, has fairly high mannanase activity and has the potential to hydrolyse mannan in feed ingredients.
ACKNOWLEDGMENTS
This study was funded by the DIPA Directorate of Research and Community Service Ministry of Research Technology and Higher Education Republic of Indonesia for the fiscal year 2016, number 042.06.0/2016, dated 7 December, 2015, with a letter of contact research agreement with number 37/UN21.6/LT/2016. The authors are very grateful to the Rector of Jambi University and the Directorate of Higher Education who have allowed the implementation of this study.
REFERENCES
- Abdollahi, M.R., B.J. Hosking, D. Ning and V. Ravindran, 2016. Influence of palm kernel meal inclusion and exogenous enzyme supplementation on growth performance, energy utilization and nutrient digestibility in young broilers. Asian-Aust. J. Anim. Sci., 29: 539-548.
Direct Link - Dusterhoft, E.M., A.G.J. Voragen and F.M. Engels, 1991. Non-starch polysaccharides from sunflower (Helianthus annuus) meal and palm kernel (Elaeis guineenis) meal-preparation of cell wall material and extraction of polysaccharide fractions. J. Sci. Food Agric., 55: 411-422.
CrossRefDirect Link - El-Sharounya, E.E., N.M.K. El-Toukhyb, N.A. El-Sersyc and A.A. El-Aal El-Gayara, 2015. Optimization and purification of mannanase produced by an alkaliphilic-thermotolerant Bacillus cereus N1 isolated from Bani Salama Lake in Wadi El-Natron. Biotechnol. Biotechnol. Equip., 29: 315-323.
CrossRefDirect Link - Wang, M., S. You, S. Zhang, W. Qi and Z. Liu et al., 2013. Purification, characterization and production of β-mannanase from Bacillus subtilis TJ-102 and its application in gluco-mannooligosaccharides preparation. Eur. Food Res. Technol., 237: 399-408.
CrossRefDirect Link - Pangsri, P., Y. Piwpankaew, A. Ingkakul, S. Nitisinprasert and S. Keawsompong, 2015. Characterization of mannanase from Bacillus circulans NT6.7 and its application in mannooligosaccharides preparation as prebiotic. SpringerPlus, Vol. 4.
CrossRefDirect Link - Harnentis, Y. Marlida, Y. Rizal and M.E. Mahata, 2013. Isolation, characterization and production of mannanase from thermophilic bacteria to increase the feed quality. Pak. J. Nutr., 12: 360-364.
CrossRefDirect Link - Adiguzel, A., H. Nadaroglu and G. Adiguzel, 2015. Purification and characterization of β-mannanase from Bacillus pumilus (M27) and its applications in some fruit juices. J. Food Sci. Technol., 52: 5292-5298.
CrossRefDirect Link - Cho, S.J., 2009. Isolation and characterization of mannanase producing Bacillus amyloliquefaciens CS47 from horse feces. J. Live Sci., 19: 1724-1730.
CrossRefDirect Link - Ge, J., R. Du, D. Zhao, G. Song, M. Jin and W. Ping, 2016. Kinetic study of a β-mannanase from the Bacillus licheniformis HDYM-04 and its decolorization ability of twenty-two structurally different dyes. Springer Plus, Vol. 5, No. 1.
CrossRefDirect Link - Itakura, S., H. Tanaka and A. Inoki, 1997. Distribution of cellulases, glucose and related substance in the body of Coptotermes formosanus. Mat. Organisme., 31: 17-29.
Direct Link - Gupta, P., K. Samant and A. Sahu, 2012. Isolation of cellulose-degrading bacteria and determination of their cellulolytic potential. Int. J. Microbiol., Vol. 2012.
CrossRef - Hinzato, S., M. Muramatsu, T. Matsui and Y. Watanabe, 2005. Molecular phylogenetic diversity of the Bacterial community in the gut of the termite Coptotermes formosanus. Biosci. Biotechnol. Biochem., 69: 1145-1155.
CrossRefDirect Link - Pourramezan, Z., G.R. Ghezelbash, B. Romani, S. Ziaei and A. Hedayatkhah, 2012. Screening and identification of newly isolated cellulose-degrading bacteria from the gut of xylophagous termite Microcerotermes diversus (Silvestri). Microbiology, 81: 736-742.
CrossRefDirect Link - Kuhnigk, T., E.M. Borst, A. Ritter, P. Kampfer, A. Graf, H. Hertel and H. Konig, 1994. Degradation of lignin monomers by the hindgut flora of xylophagous termites. J. Syst. Applied Microbiol., 17: 76-85.
CrossRefDirect Link - Kuhnigk, T. and H. Konig, 1997. Degradation of dimeric lignin model compounds by aerobic bacteria isolated from the hindgut of xylophagous termites. J. Basic Microbiol., 37: 205-211.
CrossRefDirect Link - Wenzel, M., I. Schonig, M. Berchtold, P. Kampfer and H. Konig, 2002. Aerobic and facultatively anaerobic cellulolytic bacteria from the gut of the termite Zootermopsis angusticollis. J. Applied Microbiol., 92: 32-40.
CrossRefDirect Link - Purwadaria, T., P.P. Ketaren, A.P. Sinurat and I. Sutikno, 2003. Identification and evaluation of fiber hydrolytic enzymes in the extract of termites (Glyptotermes montanus) for poultry feed application. Indon. J. Agric. Sci., 4: 40-47.
Direct Link - Sanders, E.R., 2012. Aseptic laboratory techniques: Plating methods. J. Vis. Exp., Vol. 63.
CrossRefDirect Link - Miller, G.L., 1959. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem., 31: 426-428.
CrossRefDirect Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Marchesi, J.R., T. Sato, A.J. Weightman, T.A. Martin, J.C. Fry, S.J. Hiom and W.G. Wade, 1998. Design and evaluation of useful bacterium-specific PCR primers that amplify genes coding for bacterial 16S rRNA. Applied Environ. Microbiol., 64: 795-799.
Direct Link - Maki, M., K.T. Leung and W. Qin, 2009. The prospects of cellulase-producing bacteria for the bioconversion of lignocellulosic biomass. Int. J. Biol. Sci., 5: 500-516.
CrossRefDirect Link - Sachlehner, A. and D. Haltrich, 1999. Purification and some properties of a thermostable acidic endo-β-1,4-D-mannanase from Sclerotium (Athelia) rolfsii. FEMS Microbiol. Lett., 177: 47-55.
CrossRefDirect Link - Adebayo-Tayo, B., T. Elelu, G. Akinola and I. Oyinloye, 2013. Screening and production of mannanase by Bacillus strains isolated from fermented food condiments. Innov. Roman. Food Biotechnol., 13: 53-62.
Direct Link - Sudathip, C., N. Suda, P. Kulwadee and A. Jirawan, 2015. Identification of suitable condition for mannanase production by Bacillus sp. GA2(1). Br. Biotechnol. J., 5: 92-97.
Direct Link - Utami, W., A. Meryandini and K.G. Wiryawan, 2013. Characterization of bacterial mannanase for hydrolyzing palm kernel cake to produce manno-oligosaccharides prebiotics. Media Peternakan, 36: 192-196.
CrossRefDirect Link - Zurmiati, Wizna, M.H. Abbas and M.E. Mahata, 2017. Production of extracellular β-mannanase by Bacillus amyloliquefaciens on a coconut waste substrate. Pak. J. Nutr., 16: 700-707.
CrossRefDirect Link - Gupta, R., P. Gigras, H. Mohapatra, V.K. Goswami and B. Chauhan, 2003. Microbial α-amylases: A biotechnological perspective. Process Biochem., 38: 1599-1616.
CrossRefDirect Link - Chauhan, P.S., A. Bharadwaj, N. Puri and N. Gupta, 2014. Optimization of medium composition for alkali-thermostable mannanase production by Bacillus nealsonii PN-11 in submerged fermentation. Int. J. Curr. Microbiol. Applied Sci., 3: 1033-1045.
Direct Link - Dan, Z., P. Wenxiang, S. Gang, L. Hongzhi, L. Xue and G. Jingping, 2012. Optimization of mannanase production by Bacillus sp. HDYM-05 through factorial method and response surface methodology. Afr. J. Microbiol. Res., 6: 176-183.
Direct Link - Bommarius, A.S. and J.M. Broering, 2005. Established and novel tools to investigate biocatalyst stability. Biocatal. Biotransform., 23: 125-139.
CrossRefDirect Link - Jiang, Z.Q., Y. Wei, D. Li, L. Li, P. Chai and I. Kusakake, 2006. High-level production, purification and characterization of a thermostable β-mannanase from the newly isolated Bacillus subtilis WY34. Carbohydr. Polym., 66: 88-96.
CrossRefDirect Link - Mou, H., F. Zhou, X. Jiang and Z. Liu, 2011. Production, purification and properties of β-mannanase from soil bacterium Bacillus circulans M-21. J. Food Biochem., 35: 1451-1460.
CrossRefDirect Link - Sreena, C.P., N.K. Resna and D. Sebastian, 2015. Isolation and characterization of cellulase producing bacteria from the gut of termites (Odontotermes and Heterotermes species). Br. Biotechnol. J., 9: 1-10.
Direct Link - Ramin, M., A.R. Alimon, N. Abdullah, J.M. Panandam and K. Sijam, 2008. Isolation and identification of three species of bacteria from the termite Coptotermes curvignathus (Holmgren) present in the Vicinity of University Putra Malaysia. Res. J. Microbiol., 3: 288-292.
CrossRefDirect Link