
Zurmiati
Department of Animal Feed and Nutrition, Faculty of Animal Husbandry, Andalas University, 25163 Padang, Indonesia
Wizna
Department of Animal Feed and Nutrition, Faculty of Animal Husbandry, Andalas University, 25163 Padang, Indonesia
M. Hafil Abbas
Department of Livestock Production, Faculty of Animal Husbandry, Andalas University, 25163 Padang, Indonesia
Maria Endo Mahata
Department of Animal Feed and Nutrition, Faculty of Animal Husbandry, Andalas University, 25163 Padang, Indonesia
LiveDNA: 62.17378
Pakistan Journal of Nutrition
Year: 2017 | Volume: 16 | Issue: 9 | Page No.: 700-707







ABSTRACT
Background and Objective: Mannanase is a decomposer of the compound mannan, which is produced by many microorganisms including bacteria. This enzyme can be utilized in various industrial processes, such as in the animal feed industry to increase the nutritional value of the feed. This study aims to optimize the production and function of the mannanase enzyme, which is produced by Bacillus amyloliquefaciens on a coconut waste substrate. Methodology: This study was designed in the form of laboratory experiments described herein and repeated 3 times. The substrate dose, incubation time, optimum pH and temperature of the mannanase enzyme were measured. Results: The results of this study showed that the best conditions were a substrate ratio of 80% coconut waste to 20% tofu waste, a 48 h incubation time, a pH of 6.5 and a temperature of 40°C yielding a mannanase enzymatic activity of 5.13 U mL–1. When locust bean gum was added to the substrate composed of coconut and tofu waste, the best conditions were a dose of 0.6%, a 48 h incubation time, a pH = 6.5 and a temperature of 40°C yielding a 5.92 U mL–1 mannanase enzymatic activity. Conclusion: It is concluded that the activity of the mannanase enzyme with the substrate composed of coconut and tofu waste is not much different from that with the substrate containing locust bean gum, with the difference being only 13.34%.
PDF Abstract XML References Citation
Received: June 21, 2017;
Accepted: July 31, 2017;
Published: August 15, 2017
Copyright: © 2017. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Zurmiati, Wizna, M. Hafil Abbas and Maria Endo Mahata, 2017. Production of Extracellular β-mannanase by Bacillus amyloliquefaciens on a Coconut Waste Substrate. Pakistan Journal of Nutrition, 16: 700-707.
DOI: 10.3923/pjn.2017.700.707
URL: https://scialert.net/abstract/?doi=pjn.2017.700.707
DOI: 10.3923/pjn.2017.700.707
URL: https://scialert.net/abstract/?doi=pjn.2017.700.707
INTRODUCTION
Hemicellulose is a complex polysaccharide that is abundant in the cell walls of higher plants. Mannan is the major hemicellulose in cell walls, endosperm and plant seeds1. Based on the sugar composition, hemicellulose can be distinguished as xylan, mannan, arabinogalactan or arabian. Mannan can be grouped into four subfamilies depending on the side chain structure: Mannan, glucomannan, galactomannan and galactoglucomannan2. Mannan and heteromannan are widespread in nature as part of the hemicellulosic fraction and are found in plant cell walls. They can decrease the digestibility and use of nutrients as well as nutrient encapsulation and they can also increase the viscosity of feed in the gut, which ultimately reduces the rate of hydrolysis and nutrient absorption in rations. Hetero-1,4-D-xilan and hetero-1,4-D-mannan (galactose and glucomannan) are often used in various industrial processes3. The mannan compound is a polymer of mannose, while galactomannan is a galactose-intensive mannose polymer4. Mannan is particularly present in endosperm, copra, palm, coffee and locust beans5. Coconut waste is known to contain a high mannan content. Coconut waste contains polysaccharide components in the form of galactomannan 61%, mannan 26% and cellulose 13%6. Coconut waste is industrial waste or household waste that has the potential to be used as poultry feed because it is easily obtained from the remnants of the production of traditional virgin coconut oil (VCO). However, the utilization of coconut waste as livestock feed is still limited because of its high mannan content.
Mannan can be degraded by the mannanase enzyme. The mannanase enzyme is the enzyme that can decompose heteromannan into mannose, glucose and galactose. Mannan degradation requires a variety of enzymes such as β-mannanase (EC 3.2.178), which can hydrolyze β-1,4-D mannopyranosyl in the main skeleton of mannan polymers such as galactomannan and glucomannan to produce short chain mannooligosaccharides. Furthermore, the compound is hydrolyzed by the action of the β-mannosidase enzyme (EC 3.2.1.25) and α-galactosidase (EC 3.2.1.22), yielding 7 mannose and galactose molecules7. The mannanase enzyme can be produced by microorganisms commonly found in the ground, compost, or rumen animals3. Several Bacillus species that produce the mannanase enzyme are as follows: B. licheniformis8, Bacillus sp. strain JAMB-7509, B. subtilis WY 3410, B. subtilis B3611, B. subtilis WL-312, B. circulans CGMCC155413, Bacillus sp. MSJ-514, B. licheniformis15, B. subtilis BCC4105116, B. subtilis Bs517, B. subtilis G118, B. subtilis MAFICS 1119, B. subtilis TJ 10220, B. nealsonii PN 1121, B. subtilis YH1222, B. circulans NT 6.723, B. amyloliquefaciens 10A124 and B. amyloliquefaciens CS 4725.
Several studies regarding β-mannanase characterization have been published. Bacteria of B. circulans NT 6.7 produce the mannanase enzyme at an optimum pH and temperature of 6.0 and 60°C, respectively23. B. subtilis MAFICS 11 produce the mannanase enzyme at an optimum pH and temperature of 6.0 and 5.0°C, respectively19. Bacillus pumilus (M27) produce the mannanase enzyme at an optimum pH and temperature of 8 and 60°C, respectively26. Bacillus sp., produce the mannanase enzyme at an optimum pH and temperature of 7.0 and 60°C27. Bacillus amyloliquefaciens bacteria isolated from Lunang Kab, Pesisir Selatan, West Sumatra, Indonesia, produce several types of enzymes and give a good effect on the ration efficiency of ducks and improve feed quality. The administration of B. amyloliquefaciens to the drinking water of starter Pitalah ducks for 6 weeks decreased feed consumption and improved the efficiency of the ration, with a>15% decrease in feed conversion due to an increase in the small intestinal CFU and acidity. This, in turn, resulted in an increase in the income over feed cost (IOFC) but has no influence on weight gain28. The fermentation process of rice bran with B. amyloliquefaciens caused a change of nutrient content, the fermentation to increase digestibility of crude protein, calcium, phosphor, but does not decrease crude fiber29. However, characterization of the mannanase enzyme in B. amyloliquefaciens bacteria has never been performed. From the description above, study is conducted to discover the optimum conditions for the production of the mannanase enzyme by B. amyloliquefaciens.
MATERIALS AND METHODS
Organisms: The B. amyloliquefaciens bacteria were isolated from Lunang. Kab, Pesisir Selatan, West Sumatra, Indonesia.
Determination of the best substrate ratio of coconut waste to the tofu waste, amount of locust bean gum and incubation time for β-mannanase production: To obtain the maximum production of β-mannanase, a production curve was made. The first step to make a starter is to take 1 g of B. amyloliquefaciens cultures and inoculate it on 25 mL of nutrient broth medium. The culture was incubated at 40°C and 120 rpm for 48 h and then, 4% starter is inoculated in the basic production medium [0.35% yeast extract, 0.35% tripton, 0.035% MgSO4, 0.245% KH2PO4, 0.175% (NH4)2SO4 and 0.2% NaCl ratio of the coconut waste to tofu waste according to the treatment, i.e., 90:10, 80:20, 70:30 and 60:40%]. The dosage of locust bean gum as a substitute into the substrate of coconut waste and tofu waste varied according to the treatment (0.4, 0.5 and 0.6%). The bacteria were incubated at 40°C and 120 rpm. Sampling is performed every 12 h, starting from the 12th h until the 120th h and each treatment is repeated 3 times. Samples as much as 3 mL were taken to be tested for β-mannanase activity. The β-mannanase activity test utilized the Nelson method30.
Determination of the best pH for the medium for the production of β-mannanase: The determination of the pH of the medium for the optimum production of β-mannanase by B. amyloliquefaciens was performed by growing B. amyloliquefaciens in production medium with pH values of 2, 2.5, 3, 3.5, 4,4.5, 5, 5.5, 6, 6.5, 7, 7.5, 8 , 8.5, 9, 9.5 and 10 after incubation at 40°C for 48 h on a substrate with a ratio of coconut pulp to tofu waste of 80:20% and the addition of 0.6% locust bean gum. The supernatant was then removed to test for its β-mannanase activity.
Determination of the best temperature of the medium for the production of β-mannanase: The determination of the medium temperature for the optimum production of β-mannanase by B. amyloliquefaciens was performed by growing B. amyloliquefaciens in production medium with temperatures of 20, 25, 30, 35, 40, 45, 50, 55 and 60°C. After incubation at a pH of 6.5 for 48 h on a substrate with a ratio of coconut waste to tofu waste of 80:20% and the addition of 0.6% locust bean gum, the supernatant was then removed to test for its β-mannanase activity.
Mannanase: To measure the mannanase activity using the method of Nelson30, the test for the activity of mannanase was carried out by taking 1 mL of crude enzyme and adding 1 mL of substrate (0.5% locust bean gum in 0.01 M phosphate buffer at a pH of 7) and then mixing them using a vortex mixer. They were then reacted in a shaker water bath (MEMMERT, Waterbath, Model WNB 14) for 30 min at 60°C. Furthermore, the reaction is stopped by placing it in boiling water for 5 min. Then, the hydrolyzed products were centrifuged (Hitachi himac CR21GIII) with a speed 5000 rpm for 30 min. Approximately 1 mL of hydrolysate was combined with Nelson AB reagent (1 mL) and cooked in boiling water for 5 min. Then, phosphomolybdate reagent (1 mL) and distilled water (7 mL) were added, followed by the release of glucose, which is read using a spectrophotometer (Shimadzu UV-1800 UV-vis spectrophotometer) at a wavelength of 575 nm.
Experimental design: This study was designed in the form of the described laboratory experiments repeated 3 times. To obtain the optimum results and activity of the mannanase enzyme, the coconut waste is combined with the tofu waste in the following ratios: A. 90:10, B. 80:20, C. 70:30 and D. 60:40%. Then, locust bean gum in dosages of 0.6, 0.5 and 0.4% was added to the substrate.
RESULTS AND DISCUSSION
Influence of the incubation time, ratio of coconut waste to tofu waste and addition of locust bean gum as a growth substrate on the production of β-mannanase by B. amyloliquefaciens: The experiments were performed on substrates with the following ratios of coconut to tofu waste: 90:10%, 80:20%, 70:30% and 60:40%. Locust bean gum was added at concentrations of 0.4, 0.5 and 0.6%. To determine the optimum conditions for β-mannanase production by B. amyloliquefaciens, incubation times of 12, 24, 36, 48, 60, 72, 84, 96, 108 and 120 h were evaluated (Fig. 1, 2). The pH and temperature of the medium were 7.0 and 40°C, respectively. The production of β-mannanase by B. amyloliquefaciens is expressed in units of enzyme activity because the concentration of the enzyme is directly proportional to the enzyme activity. At an incubation temperature of 40°C, mannanase was produced by B. amyloliquefaciens starting at 12th h, the activity continued to rise and the highest value was at 48 h with an activity of 2.24 U mL–1 on a substrate with a ratio of coconut pulp to tofu waste of 80:20%. The activity slowly decreased after 48 h until the 120th h, which had an activity of 1.62 U mL–1 (Fig. 1). Likewise with locust bean gum added into the substrate, the enzyme activity continue to increase and was the highest at 48 h, with the activity of 2.28 U mL–1 at a locust bean gum concentration of 0.6%. Then, the activity slowly decreased until 120 h, at which a value of 1.65 U mL–1 (Fig. 2) was reached.
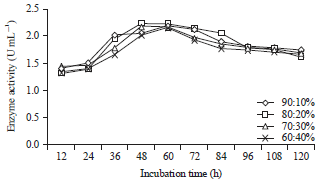 | |
| Fig. 1: | Activity of the mannanase enzyme with various incubation times and ratios of coconut and tofu waste in the substrate |
This is caused by the amount of component on the substrate as the incubation time increases along with the growth of bacterial cells, which then decreases when the required carbon source is low. Figure 1 and 2 show that once the enzyme activity reaches its optimum point at 48 h, there is a decrease in the enzyme activity.
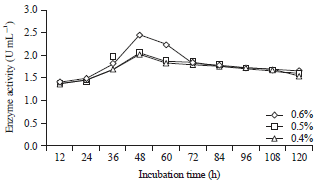 | |
| Fig. 2: | Activity of the mannanase enzyme for various incubation times and with various amounts of locust bean gum added to the substrate |
The results of this study are not much different from those reported by Sumardi31, in which the best conditions for the thermophilic aerobic bacteria Geobacillus stearothermophilus L-07 to produce β-1,4-mannanase (3.1 U mg–1) were found to be with a carbon source of 0.65% (w/v) locust bean gum, at a pH of 7.0 and with 36 h of incubation at 60°C. Additionally, the same treatment but with the temperature increased to 70°C led to the best production with incubation for 12 h (3.2 U mg–1).
Production of β-mannanase in medium with various pH values: The effect of the pH on mannanase production is presented in Fig. 3 and 4. Treatment to change the medium pH to produce β-mannanase B. amyloliquefaciens was performed on the substrate of coconut waste and tofu waste with a ratio of 80:20%, with the addition of 0.6% (w/v) locust bean gum and after 48 h of incubation time. From the experimental results, it is known that β-mannanase is produced by B. amyloliquefaciens at pH values from 2-10. At a pH of 2.0 and 10 with a substrate having a ratio of coconut waste to tofu waste of 80:20%, the β-mannanase production is very small (0.91 and 0.86 U mL–1) (Fig. 3).
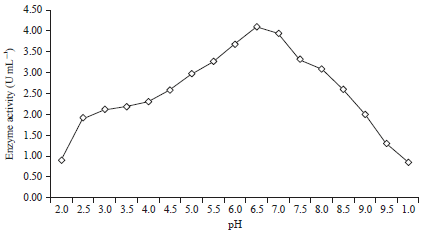 | |
| Fig. 3: | Activity of the mannanase enzyme at various pH values with a substrate of coconut and tofu waste (80:20%) |
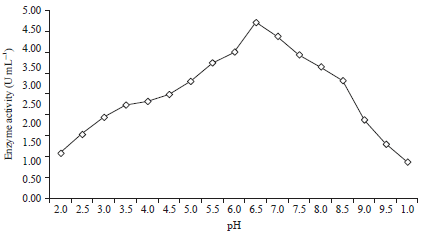 | |
| Fig. 4: | Activity of the mannanase enzyme at various pH values with 0.6% locust beam gum added to the substrate |
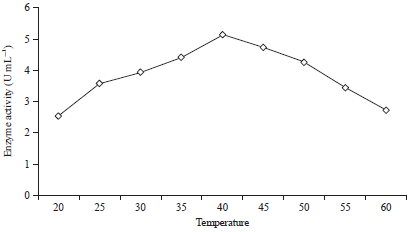 | |
| Fig. 5: | Activity of the mannanase enzyme for various temperatures with a substrate of coconut and tofu waste (80:20%) |
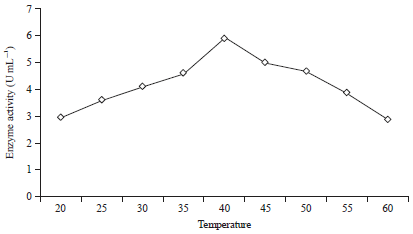 | |
| Fig. 6: | Activity of the mannanase enzyme for various temperatures with 0.6% locust beam gum added to the substrate |
Likewise with 0.6% (w/v) locust bean gum added, the enzyme activities were 1.22 and 0.99 U mL–1, respectively (Fig. 4). From the treatment to evaluate the effect of the pH of the medium with a substrate having a ratio of coconut waste to tofu waste of 80:20%, it is known that the optimum pH for growth is obtained at 6.5 (4.09 U mL–1) (Fig. 3). Likewise, with a substrate having 0.6% (w/v) locust bean gum added, it is known that the optimum pH for growth is 6.5 (4.71 U mL–1) (Fig. 4). At pH values lower or higher than the optimum pH, mannanase production decreases. This is closely related to bacterial growth. At acidic or alkaline pH, bacterial growth will be inhibited due to the elucidation of H+ and H- ions, decreased cellular stability, cell membrane enzyme inhibition and protein transport32.
The optimal pH value obtained in this study for mannanase production by B. amyloliquefaciens was similar to that for mannanase production by Bacillus subtilis YH12, with optimum conditions of a pH of 6.5 and 55°C22. Bacillus amyloliquefaciens CS 47 produces the mannanase enzyme at a pH of 6.0 and 50°C25. The optimum of pH and temperature for the activity of the mannanase enzyme produced by Bacillus sp., SWU 60 are 6.0 and 60°C, respectively33. In general, the optimum pH of mannanase is between 7.1 and 7.7, except for the Bacillus originating mannanase, whose optimum pH range is 3-6.57. Each enzyme has an optimum pH and is active in a relatively narrow pH range34 because each enzyme has an optimum pH typical for its enzyme application and thus, it will also be efficient at that pH4.
Production of β-mannanase at various temperatures: Treatment to evaluate the effect of temperature on the production of β-mannanase by B. amyloliquefaciens was performed on the substrate with a ratio of coconut waste to tofu waste of 80:20% with the addition of 0.6% (w/v) locust bean gum, with an incubation time of 48 h and at a pH of 6.5. The effect of temperature on the production of the mannanase enzyme by B. amyloliquefaciens can be seen in (Fig. 5, 6). From the experiment results, it is known that B. amyloliquefaciens produced β-mannanase in a temperature range of 20-60°C. In the temperature range of 20-60°C, the production of β-mannanase on the substrate with a ratio of coconut waste to tofu waste of 80:20%, was very small (2.53 and 2.68 U mL–1) (Fig. 5). The production of β-mannanase with the addition of 0.6% (w/v) locust bean gum to the substrate is also very small (2.96 and 2.86 U mL–1) (Fig. 6). The enzyme activity will increase with an increase of the temperature until the optimum temperature is reached. However, after passing the optimum temperature, the enzyme activity will decrease. Increased temperatures will increase the kinetic energy of the enzymes so that the vibrational and rotational movement of the enzymes and substrates will be increase, which will cause greater opportunities to collide. There is an increase in temperature up to a certain limit that will cause increased catalytic activity of the enzyme and increased the process of enzyme destruction34. From the effect of the media temperature on a substrate with a ratio of coconut to tofu waste of 80:20%, it is known that the optimum medium growth temperature is 40°C (5.13 U mL–1) (Fig. 5). With 0.6% (w/v) locust bean gum added to the substrate, it is known that the optimum temperature of growth is obtained at a medium temperature of 40°C (5.92 U mL–1) (Fig. 6).
The production of mannanase decreases at a temperature higher than the optimum temperature. This is due to the increase of the melting point of the fat layer and the depletion of the cell membrane (leak) as well as the occurrence of protein denaturation that is sensitive to heat, thus, the growth of bacteria is inhibited and the production of the enzyme decreases. The cell membrane adjusts the composition of saturated fatty acids according to changes in the ambient temperature35. The results of this study are not much different from those reported by Kanjanavas et al.36, in which B. licheniformis THCM 3.1 produces the mannanase enzyme at an optimum temperature and pH of 45°C and 9.0, respectively. Bacillus amyloliquefaciens 10A1 with an ammonium nitrate nitrogen source produced a mannanase enzyme activity of 61.5 U mg–1 at a temperature of 30°C and pH of 724. The best conditions for producing β-1,4-mannanase (3.1 U mg–1) by the bacteria of the thermophilic aerobic Geobacillus stearothermophilus L 07 were with a carbon source of 0.65% (w/v) locust bean gum, a pH of 7.0 and 36 h of incubation time at 60°C. The same treatment but with the temperature increased to 70°C were the best conditions for incubation for 12 h (3.2 U mg–1)31. Based on the results of this experiment, it can be seen that a substrate with a composition of 80% coconut waste and 20% tofu waste with the addition of 0.6% locust bean gum is better than the other treatments because it produces the highest mannanase activity. The mannanase enzyme produced by Bacillus amyloliquefaciens can be applied well at a pH of 6.5 and a temperature of 40°C, but at lower and higher pH values or higher temperatures, the ability to produce the mannanase enzyme will not be optimal.
CONCLUSION
Bacteria of B. amyloliquefaciens produce the mannanase enzyme and the activity of the mannanase enzyme with a substrate that is a combination of coconut and tofu waste is not much different compared to with a substrate using locust bean gum, with only a 13.34% difference and the optimum pH and temperature are the same.
SIGNIFICANCE STATEMENTS
This study discovers the optimum conditions for the production of the mannanase enzyme by Bacillus amyloliquefaciens bacteria, which can be beneficial for the livestock industry to improve the nutritional value of feed ingredients. This study will help researchers to uncover the ability of Bacillus amyloliquefaciens to produce a mannanase enzyme that can be explored by other researchers. Thus, a new theory on the optimum conditions for Bacillus amyloliquefaciens to produce the mannanase enzyme may be arrived at.
ACKNOWLEDGMENTS
This study was supported by a National Strategic Research Grant from the Directorate General of Higher Education, Department of National Education, Republic of Indonesia grant program (124/SP2H/LT/DRPM/III/-2015). Authors are very grateful to the Minister of Research Technology and Higher Education who allowed us to conduct this study.
REFERENCES
- Petkowicz, C.L.D.O., F. Reicher, H. Chanzy, F.R. Taravel and R. Vuong, 2001. Linear mannan in the endosperm of Schizolobium amazonicum. Carbohydr. Polym., 44: 107-112.
CrossRefDirect Link - Moreira, L.R. and E.X. Filho, 2008. An overview of mannan structure and mannan-degrading enzyme systems. Applied Microbiol. Biotechnol., 79: 165-178.
CrossRefPubMedDirect Link - Hilge, M., S.M. Gloor, W. Rypniewski, O. Sauer and T.D. Heightman et al., 1998. High-resolution native and complex structures of thermostable β-mannanase from Thermomonospora fusca-substrate specificity in glycosyl hydrolase family 5. Structure, 6: 1433-1444.
CrossRefDirect Link - Ooi, T. and D. Kikuchi, 1995. Purification and some properties of β-mannanase from Bacillus sp. World J. Microbiol. Biotechnol., 11: 310-314.
CrossRefDirect Link - Purawisastra, S., 2001. Effect of coconut galactomannan isolate on decreased serum cholesterol level of rabbit. Warta Litbang Kesehatan, Vol. 5, No. 3-4.
Direct Link - Duffaud, G.D., C.M. McCutchen, P. Leduc, K.N. Parker and R.M. Kelly, 1997. Purification and characterization of extremely thermostable beta-mannanase, beta-mannosidase and alpha-galactosidase from the hyperthermophilic eubacterium Thermotoga neapolitana 5068. Applied Environ. Microbiol., 63: 169-177.
Direct Link - Zhang, J., Z. He and K. Hu, 2000. Purification and characterization of β-mannanase from Bacillus licheniformis for industrial use. Biotechnol. Lett., 22: 1375-1378.
CrossRefDirect Link - Takeda, N., K. Hirasawa, K. Uchimura, Y. Nogi and Y. Hatada et al., 2004. Purification and enzymatic properties of a highly alkaline mannanase from alkaliphilic Bacillus sp. strain JAMB-750. Int. J. Biol. Macromol., 4: 67-74.
Direct Link - Jiang, Z.Q., Y. Wei, D. Li, L. Li, P. Chai and I. Kusakake, 2006. High-level production, purification and characterization of a thermostable β-mannanase from the newly isolated Bacillus subtilis WY34. Carbohydr. Polym., 66: 88-96.
CrossRefDirect Link - Li, Y.N., K. Meng, Y.R. Wang and B. Yao, 2006. A β-mannanase from Bacillus subtilis B36: Purification, properties, sequencing, gene cloning and expression in Escherichia coli. Zeitschrift fur Naturforschung C, 61: 840-846.
CrossRefDirect Link - Yoon, K.H., S. Chung and B.L. Lim, 2008. Characterization of the Bacillus subtilis WL-3 mannanase from a recombinant Escherichia coli. J. Microbiol., 46: 344-349.
CrossRefDirect Link - Yang, P., Y. Li, Y. Wang, K. Meng and H. Luo et al., 2009. A novel β-mannanase with high specific activity from Bacillus circulans CGMCC1554: Gene cloning, expression and enzymatic characterization. Applied Biochem. Biotechnol., 159: 85-94.
CrossRefDirect Link - Zhang, M., X.L. Chen, Z.H. Zhang, C.Y. Sun and L.L. Chen et al., 2009. Purification and functional characterization of endo-β-mannanase MAN5 and its application in oligosaccharide production from konjac flour. Applied Microbiol. Biotechnol., 83: 865-873.
CrossRefDirect Link - Songsiriritthigul, C., B. Buranabanyat, D. Haltrich and M. Yamabhai, 2010. Efficient recombinant expression and secretion of a thermostable GH26 mannan endo-1,4-β-mannosidase from Bacillus licheniformis in Escherichia coli. Microb. Cell Factories, Vol. 9.
CrossRefDirect Link - Summpunn, P., S. Chaijan, D. Isarangkul, S. Wiyakrutta and V. Meevootisom, 2011. Characterization, gene cloning and heterologous expression of β-mannanase from a thermophilic Bacillus subtilis. J. Microbiol., 49: 86-93.
CrossRefDirect Link - Huang, J.L., L.X. Bao, H.Y. Zou, S.G. Che and G.X. Wang, 2012. High-level production of a cold-active B-mannanase from Bacillus subtilis BS5 and its molecular cloning and expression. Mol. Gen. Microbiol. Virol., 27: 14-17.
CrossRefDirect Link - Vu, T.T., D.T. Quyen, T.T. Dao and S.T. Nguyen, 2012. Cloning, high-level expression, purification and properties of a novel endo-beta-1,4-mannanase from Bacillus subtilis G1 in Pichia pastoris. J. Microbiol. Biotechnol., 22: 331-338.
PubMedDirect Link - Lv, J., Y. Chen, H. Pei, W. Yang, Z. Li, B. Dong and Y. Cao, 2013. Cloning, expression and characterization of β-mannanase from Bacillus subtilis MAFIC-S11 in Pichia pastoris. Applied Biochem. Biotechnol., 169: 2326-2340.
CrossRefDirect Link - Wang, M., S. You, S. Zhang, W. Qi and Z. Liu et al., 2013. Purification, characterization and production of β-mannanase from Bacillus subtilis TJ-102 and its application in gluco-mannooligosaccharides preparation. Eur. Food Res. Technol., 237: 399-408.
CrossRefDirect Link - Chauhan, P.S., A. Bharadwaj, N. Puri and N. Gupta, 2014. Optimization of medium composition for alkali-thermostable mannanase production by Bacillus nealsonii PN-11 in submerged fermentation. Int. J. Curr. Microbiol. Applied Sci., 3: 1033-1045.
Direct Link - Liu, H.X., J.S. Gong, H. Li, Z.M. Lu and H. Li et al., 2015. Biochemical characterization and cloning of an endo-1,4-β-mannanase from Bacillus subtilis YH12 with unusually broad substrate profile. Process Biochem., 50: 712-721.
CrossRefDirect Link - Pangsri, P., Y. Piwpankaew, A. Ingkakul, S. Nitisinprasert and S. Keawsompong, 2015. Characterization of mannanase from Bacillus circulans NT6.7 and its application in mannooligosaccharides preparation as prebiotic. SpringerPlus, Vol. 4.
CrossRefDirect Link - Mabrouk, M.E.M. and A.M.D. El Ahwany, 2008. Production of 946-mannanase by Bacillus amylolequifaciens 10A1 cultured on potato peels. Afr. J. Biotechnol., 7: 1123-1128.
Direct Link - Cho, S.J., 2009. Isolation and characterization of mannanase producing Bacillus amyloliquefaciens CS47 from horse feces. J. Live Sci., 19: 1724-1730.
CrossRefDirect Link - Adiguzel, A., H. Nadaroglu and G. Adiguzel, 2015. Purification and characterization of β-mannanase from Bacillus pumilus (M27) and its applications in some fruit juices. J. Food Sci. Technol., 52: 5292-5298.
CrossRefDirect Link - Harnentis, Y. Marlida, Y. Rizal and M.E. Mahata, 2013. Isolation, characterization and production of mannanase from thermophilic bacteria to increase the feed quality. Pak. J. Nutr., 12: 360-364.
CrossRefDirect Link - Zurmiati, Wizna, M.H. Abbas, M.E. Mahata and R. Fauzano, 2017. Effect of Bacillus amyloliquefaciens as a probiotic on growth performance parameters of Pitalah ducks. Int. J. Poult. Sci., 16: 147-153.
CrossRefDirect Link - Wizna, H. Abbas, Y. Rizal, A. Djulardi and H. Muis, 2012. The effect of supplementation of micro nutrient on nutrient rice bran which fermented by Bacillus amyloliquefaciens. Pak. J. Nutr., 11: 439-443.
CrossRefDirect Link - Nelson, N., 1944. A photometric adaptation of the Somogyi method for the determination of glucose. J. Biol. Chem., 153: 375-380.
CrossRefDirect Link - Booth, I.R., 1985. Regulation of cytoplasmic pH in bacteria. Microbiol. Rev., 49: 359-378.
Direct Link - Seesom, W., P. Thongket, T. Yamamoto, S. Takenaka, T. Sakamoto and W. Sukhumsirichart, 2017. Purification, characterization and overexpression of an endo-1,4-β-mannanase from thermotolerant Bacillus sp. SWU60. World J. Microbiol. Biotechnol., Vol. 33.
CrossRefDirect Link - Kanjanavas, P., P. Khawsak, A. Pakpitcharoen, S. Areekit and T. Sriyaphai et al., 2009. Over-expression and characterization of the alkalophilic, organic solvent-tolerant and thermotolerant endo-1,4-β-mannanase from Bacillus licheniformis isolate THCM 3.1. ScienceAsia, 35: 14-23.
CrossRefDirect Link