
Dina Tri Marya
Department of Animal Science and Industry, Faculty of Animal Science, Universitas Gadjah Mada, Yogyakarta, Indonesia
Nurliyani
Department of Animal Product Technology, Faculty of Animal Science, Universitas Gadjah Mada, Yogyakarta, Indonesia
Widodo
Department of Animal Product Technology, Faculty of Animal Science, Universitas Gadjah Mada, Yogyakarta, Indonesia
Sunarti
Department of Biochemistry, Faculty of Medicine, Universitas Gadjah Mada, Yogyakarta, Indonesia
Pakistan Journal of Nutrition
Year: 2017 | Volume: 16 | Issue: 6 | Page No.: 451-456







ABSTRACT
Objective: This study aimed to evaluate the microbiological and chemical characteristics and antioxidant activity of fermented milk produced by different starter combinations. Materials and Methods: Nine combinations of starter produced fermented dairy products using single-starter Lactic Acid Bacteria (LAB), a combination of starter LAB and a combination of starter LAB and yeast. The starter combinations were as follows: Lactobacillus plantarum Dad 13, L. plantarum Dad 13+Lactococcus lactis, L. plantarum Dad 13+Saccharomyces cerevisiae, L. plantarum Dad 13+Kluyveromyces marxianus, L. plantarum Dad 13+L. lactis+S. cerevisiae, L. plantarum Dad 13+L. lactis+K. marxianus, L. lactis, L. lactis+S. cerevisiae and L. lactis+K. marxianus. Total LAB and yeast were analyzed using the Total Plate Count (TPC) method. Chemical characteristics were identified by the values of pH, titratable acidity and ethanol contents. Antioxidant activities were determined by calculating the percentage of 2,2-diphenyl-1-picrylhydrazyl (DPPH) radical scavenging. Results: Present study showed that the combination of starters did not differ significantly across total LAB and yeast. However, there were significantly differences in pH, acidity, alcohol, β-carotene and antioxidant activity. The fermented milk obtained from L. plantarum Dad 13+L. lactis+K. marxianus had the lowest pH and the highest acidity and alcohol content. The combination L. plantarum Dad 13+L. lactis+S. cerevisiae produced the maximum β-carotene content and the combination of L. plantarum Dad 13+ L. lactis+S. cerevisiae had the best antioxidant activity. Conclusion: It is concluded that a combination of starter L. plantarum Dad 13, L. lactis and S. cerevisiae can be used to improve the chemical quality and antioxidant activity of fermented milk.
PDF Abstract XML References Citation
Received: February 09, 2017;
Accepted: April 18, 2017;
Published: May 15, 2017
Copyright: © 2017. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Dina Tri Marya, Nurliyani, Widodo and Sunarti, 2017. Characterization and Antioxidant Activity of Fermented Milk Produced with a Starter Combination. Pakistan Journal of Nutrition, 16: 451-456.
DOI: 10.3923/pjn.2017.451.456
URL: https://scialert.net/abstract/?doi=pjn.2017.451.456
DOI: 10.3923/pjn.2017.451.456
URL: https://scialert.net/abstract/?doi=pjn.2017.451.456
INTRODUCTION
Lactobacillus plantarum Dad 13 is a local isolate from dadih, a traditional Indonesian fermented milk from West Sumatra. Generally, dadih is produced from buffalo milk and fermented in bamboo tubes. Pato et al.1 and Nurliyani et al.2 reported that L. plantarum Dad 13 and L. lactis subsp., lactis isolated from dadih can live in the gastric juices and are resistant to bile salt, indicating probiotic activity. According to Rahayu et al.3, L. plantarum Dad 13 can inhibit the colonization of pathogenic bacteria, such as Shigella dysenteriae dky-4, Escherichia coli dky-1 and dky-2 and Salmonella typhimurium dky-3.
Milk naturally contains components that have antioxidant capacities, such as casein, lactoferrin, vitamins and some flavonoids. Milk casein (α-casein, β-casein and κ-casein) and lactoferrin are capable of inhibiting lipid peroxidation4,5. Vitamin E, vitamin C and carotenoids are non-enzymatic radical scavengers6. Some flavonoids act as antioxidants that play a role in radical scavenging and metal ion binding7. Milk processing into fermented products provides beneficial nutrients8,9, increases antioxidant activity10, inhibits oxidation and decreases lipid peroxidation, in addition to lowering the glutathione redox ratio11. During fermentation, milk may produce bioactive peptides that possess antioxidant activity12. Farvin et al.13 found that yogurt is more oxidatively-stable than milk due to its proteolytic activity.
The role of yeast in dairy fermentation continues to show favorable progress. Yeast growth in fermented products provides flavor components14, as result of the utilization of lactose or metabolites of Lactic Acid Bacteria (LAB)15,16. The interaction between LAB and yeast results in a decline in pH and maintains the stability of viable LAB17. Presence of yeast can also increase the antioxidant activity of milk peptides related to 2,2-diphenyl-1-picrylhydrazyl (DPPH) and hydroxyl radical scavenging18. In addition, β-carotene, produced as a result of yeast metabolism, can also act as an antioxidant. Yeast has been used as a co-culture in traditional fermented products in some regions, such as kefir, koumiss, amasi and dadih. However, the composition of natural starters is unknown and it is difficult to obtain the stability of fermentation products. Efforts are underway to utilize yeast as co-starter in the manufacturing of fermented products to obtain more stable product quality19-21.
The ability of L. plantarum Dad 13 to produce fermented milk with the best microbiological, chemical and antioxidant characteristics have not yet been evaluated. Therefore, it is necessary to investigate the ability of L. plantarum Dad 13 individually or in combination with other lactic acid bacteria or a combination thereof with yeast to produce the most favorable fermented milk characteristics.
MATERIALS AND METHODS
Lactic acid bacteria and yeast: The starter culture of L. plantarum Dad 13, L. lactis, S. cerevisiae and K. marxianus was obtained from the Food and Nutrition Culture Collection (FNCC), Centre for Food and Nutrition Studies, at the Universitas Gadjah Mada in Yogyakarta, Indonesia. Skim milk (powder) was purchased from a departmental store in Yogyakarta and raw milk was obtained from the Dairy Processing Unit at the Universitas Gadjah Mada.
Preparation of culture stocks: The culture stock was made from 0.1 mL of a frozen culture that had been thawed. Culture stock was cultivated in 10 mL medium (MRS for LAB and YPG for yeast culture) and incubated at 37°C for 24 h. The culture stock was then reproduced using a similar procedure. The stocks were centrifuged at 3500 rpm for 10 min to obtain cell pellets. Approximately 2% (v/v) of glycerol and 10% (w/v) of skimmed milk was added to cell pellets to obtain a solution (1:1:1 ratio). Culture stock was stored in microtubes at -20°C.
Preparation of starter: As much as one inoculating loop from LAB and yeast culture was transferred into each tube containing 10 mL of medium (MRS for LAB, YPG for yeast), which had been sterilized by autoclaving at 121°C, 15 psi for 15 min. The LAB and yeast were incubated at 37°C for 24 h. Starter was prepared by inoculating 10% (v/v) of inoculum in 18% (w/v) skimmed milk, which had been sterilized at 110°C, 13 psi for 10 min, then incubated for 12-18 h to form the curd; these results are called the mother starter. As much as 10% (v/v) of mother starter was inoculated in 100 mL of 18% (w/v) sterilized skimmed milk followed by incubation for 12-18 h to produce a bulk starter. The starter was then inoculated into the milk to be fermented.
Preparation of fermented milk: Raw cow’s milk was pasteurized at 85°C for 30 min, then cooled to 40°C. As much as 3% (v/v) of starter was inoculated followed by incubation at 37°C for 18 h. Starter inoculated in the milk represent the single starter of LAB, the combination of LAB starter (1:1 ratio), the combination of LAB and yeast starter (2:1 ratio) or the combination of two starters of LAB and yeast (1:1:1 ratio). Nine groups of fermented milk produced the following combinations: L. plantarum Dad 13 (K1), L. plantarum Dad 13+L. lactis (K2), L. plantarum Dad 13+S. cerevisiae (K3), L. plantarum Dad 13+K. marxianus (K4), L. plantarum Dad 13+L. lactis+S. cerevisiae (K5), L. plantarum Dad 13+L. lactis+K. marxianus (K6), L. lactis (K7), L. lactis+ S. cerevisiae (K8), L. lactis+K. marxianus (K9).
Viable microorganisms: Lactobacillus plantarum Dad 13 and L. lactis were enumerated (pour plate) on MRS agar and incubated at 30°C for 48 h. Yeast was enumerated (pour plate) on chloramphenicol (100 ppm) YPG agar and incubated at 30°C for 72 h.
Chemical analysis: Titratable acidity was determined by titration of a 10 g sample (3 drops of phenolphthalein were added) with 0.1 N of NaOH. Then, its value was calculated as equivalent percent (w/w) of lactic acid and expressed as a percent of lactic acid. The pH value of the samples was measured using a pH meter (Hana-HI 98103). Ethanol content was determined by gas chromatography. β-carotene was measured according to Cagampang and Rodriguez22 with some modifications. Antioxidant activity was measured as the percentage of DPPH (2,2-diphenyl-1-picrylhydrazyl) radical scavenging activity using the method performed by Son and Lewis23.
Statistical analysis: The experimental results were expressed as the Mean±Standard Deviation. The statistical analysis was performed using IBM SPSS 22. The significance of differences between the samples were analyzed by the one-way ANOVA. Further analysis was conducted using the Duncan Multiple Range Test (DMRT) to determine significant differences between means at 5% significance level.
RESULTS
Effect on total LAB and yeast: The starter and starter combinations did not affect the LAB and yeast populations in the production of fermented milk (p>0.05) (Table 1).
Effect on pH: The starter and starter combinations significantly affected the pH value of fermented milk produced (p<0.05) (Table 2).
Effect on titratable acidity: The starter and starter combinations significantly affected the titratable acidity of fermented milk (p<0.05) (Table 2).
Ethanol content: The starter and starter combinations significantly affected the ethanol content of fermented milk (p<0.05) (Table 2).
β-carotene content: The starter and starter combinations significantly affected the content of β-carotene (p<0.05) (Table 2).
| Table 1: | Average lactic acid bacteria and yeast in fermented milk (log CFU mL–1) |
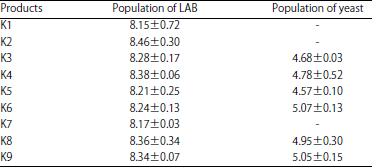 | |
K1: L. plantarum Dad 13, K2: L. plantarum Dad 13+L. lactis, K3: L. plantarum Dad 13+S. cerevisiae, K4: L. plantarum Dad 13+K. marxianus, K5: L. plantarum Dad 13+L. lactis+S. cerevisiae, K6: L. plantarum Dad 13+L. lactis +K. marxianus, K7: L. lactis, K8: L. lactis+S. cerevisiae, K9: L. lactis+K. marxianus | |
| Table 2: | pH, acidity, ethanol and β-carotene values of fermented milk based on the different combinations of starter |
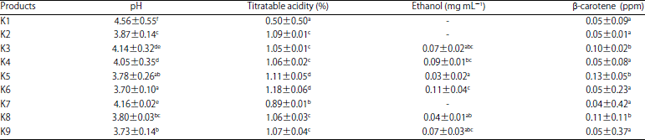 | |
K1: L. plantarum Dad 13, K2: L. plantarum Dad 13+L. lactis, K3: L. plantarum Dad 13+S. cerevisiae, K4: L. plantarum Dad 13+K. marxianus, K5: L. plantarum Dad 13+L. lactis+S. cerevisiae, K6: L. plantarum Dad 13+L. lactis+K. marxianus, K7: L. lactis, K8: L. lactis+S. cerevisiae, K9: L. lactis+K. marxianus, a-fValues with different superscripts are significantly different in a column (p<0.05) | |
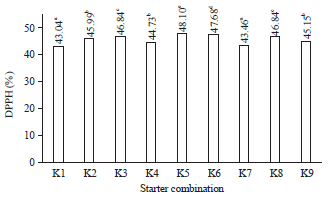 | |
| Fig. 1: | DPPH radical scavenging activity (%) of fermented milk from different starter combinations |
K1: L. plantarum Dad 13, K2: L. plantarum Dad 13+L. lactis, K3: L. plantarum Dad 13+S. cerevisiae, K4: L. plantarum Dad 13+ K. marxianus, K5: L. plantarum Dad 13+L. lactis+S. cerevisiae, K6: L. plantarum Dad 13+L. lactis+K. marxianus, K7: L. lactis, K8: L. lactis+S. cerevisiae, K9: L. lactis+K. marxianus, a-fSignificant difference between combinations (p<0.05) | |
DPPH radical scavenging activity: The starter and starter combinations affected the DPPH radical scavenging activity of fermented milk (Fig. 1).
DISCUSSION
The results of this study showed a positive interaction between L. plantarum and L. lactis, as well as between LAB and yeast (S. cerevisiae or K. marxianus) in the production of fermented milk. The LAB population tended to be higher in combination compared to the single starter LAB (L. plantarum or L. lactis only), although there was no significant difference (p>0.05). The presence of yeast as a co-starter increased LAB population as result of the capability of yeast to stimulate the growth of LAB, perhaps due to the utilization of yeast metabolites in the form of vitamins, amino acids and peptides14,24,25. The yeast population tended to be higher if K. marxianus was used as a co-starter compared to S. cerevisiae (K4>K3, K6>K5, K8>K9), possibly due to the ability of K. marxianus to utilize lactose as a carbon source15.
The pH of fermented milk depends on properties of the organic acids produced26 and is also influenced by the ability of LAB to produce lactic acid, in addition to LAB population and the type of LAB used27. Lactic acid, as the main product of fermentation, can be dissociated to H+ and CH3CHOHCOO‾ ions. The accumulation of lactic acid increased the pH of the fermented medium due to an increase in the concentration of
H+ released28. The results showed that the combination of LAB (L. plantarum+L. lactis) or the combination between LAB and yeast significantly lowered the pH of fermented milk compared to a single starter (L. plantarum or L. lactis only). Lactobacillus lactis is homofermentative bacteria which can rapidly metabolize lactose into 90% of lactic acid29. The positive synergy between L. plantarum and L. lactis in dairy fermentation30 could lead to an increased rate and amount of lactic acid24, resulting in a decline in the pH value of the medium. The contribution of yeast as a co-starter in lowering pH due to the production of acid compounds is a result of its lipolytic and proteolytic activity. Titratable acidity and pH values were inversely related. The highest titratable acidity was obtained from the starter combination of L. plantarum Dad 13+L. lactis+K. marxianus (1.18±0.06 %); however, it did not differ significantly from the combination of L. plantarum Dad 13+L. lactis+S. cerevisiae (p>0.05).
The concentration of alcohol obtained from these experiments demonstrated the effect of yeast as a co-starter. During milk fermentation, ethanol, CO2 and organic acids were produced as the result of yeast carbohydrate metabolism. Amino acids are also converted by yeast into alcohol through a series of decarboxylation and reduction31. The fermented milk using K. marxianus tended to produce higher alcohol content than the S. cerevisiae milk (p<0.05). The ability of K. marxianus to utilize lactose that was not produced by S. cerevisiae could explain this result16.
The β-carotene is a component of milk that acts as an antioxidant and can be obtained from S. cerevisiae32. Cow's milk contains β-carotene entirely derived from the diet. The antioxidant capacity of fermented milk is enhanced by the utilization of S. cerevisiae as a co-starter through increasing levels of β-carotene. The results showed a significant increase in levels of carotenoids when using S. cerevisiae as a co-starter. The highest carotenoid content was obtained from the combination of L. plantarum starter Dad 13+L. lactis S. cerevisiae (0.13±0.05 ppm) but the content was not significantly different from the combination of L. plantarum Dad 13+S. cerevisiae or L. lactis+S. cerevisiae. The other combinations were not significantly different compared to β-carotene content in raw milk (0.05 ppm).
The antioxidative properties of fermented milk in this study were measured by 2,2-diphenyl-1-picrylhydrazyl (DPPH) radical scavenging (%). Raw milk had 22.17% of DPPH radical scavenging which increased to 23.11% after pasteurization. This result is likely due to the formation of Maillard Reaction Products (MRP), which have antioxidant activity during pasteurization33. The highest rate of DPPH radical scavenging in this study was obtained from the combination of L. plantarum Dad 13+L. lactis, S. cerevisiae (48.10%) (Fig. 1). This value is relatively higher than the value obtained by Fitrotin et al.21 in the fermentation of sesame milk using L. plantarum Dad 13 (45.2% from the initial conditions, 19.3%). Other studies have reported 27% in the fermentation of whey from cow’s milk using L. lactis subsp., lactis as the starter34 and 22.2% in the manufacturing of Kunnu (traditional fermented milk from Nigeria) using L. plantarum35. Antioxidant activity of fermented milk increases due to proteolysis activity, which produces peptides13,36 that act as an electron donor and react with free radicals to form more stable products37. Lactic acid as a metabolite may also serve as a chelating agent that plays a role in inhibiting free radicals38.
Lactobacillus plantarum Dad 13 combined with L. lactis and S. cerevisiae can be used to produce the best product characteristics, particularly antioxidant activity. Saccharomyces cerevisiae in fermentation is essential to promote β-carotene and the use of L. lactis in fermentation increases the production of lactic acid, which can act as a scavenger of free radicals. The presence of L. lactis also plays a role in breaking down milk protein into bioactive peptides during fermentation to increase the antioxidant activity of the fermented milk.
CONCLUSION
It is concluded that the combination of LAB and yeast had no effect on microbiological characteristics, whereas the combination of starter L. plantarum Dad 13, L. lactis and S. cerevisiae improved the chemical quality and antioxidant activity of fermented milk.
REFERENCES
- Pato, U., I.S. Surono, Koesnandar and A. Hosono, 2004. Hypocholesterolemic effect of indigenous dadih lactic acid bacteria by deconjugation of bile salts. Asian-Aust. J. Anim. Sci., 17: 1741-1745.
Direct Link - Cervato, G., R. Cazzola and B. Cestaro, 1999. Studies on the antioxidant activity of milk caseins. Int. J. Food Sci. Nutr., 50: 291-296.
CrossRefDirect Link - Gutteridge, J.M.C., S.K. Paterson, A.W. Segal and B. Halliwell, 1981. Inhibition of lipid peroxidation by the iron-binding protein lactoferrin. Biochem. J., 199: 259-261.
CrossRefDirect Link - Zulueta, A., A. Maurizi, A. Frigola, M.J. Esteve, R. Coli and G. Burini, 2009. Antioxidant capacity of cow milk, whey and deproteinized milk. Int. Dairy J., 19: 380-385.
CrossRefDirect Link - Lindmark-Mansson, H. and B. Akesson, 2000. Antioxidative factors in milk. Br. J. Nutr., 84: 103-110.
CrossRefDirect Link - Satir, G. and Z.B. Guzel-Seydim, 2016. How kefir fermentation can affect product composition? Small Rumin. Res., 134: 1-7.
CrossRefDirect Link - Adolfson, O., S.N. Meydani and R.M. Robert, 2004. Yogurt and gut function. Am. J. Clin. Nutr., 80: 245-256.
PubMedDirect Link - Balakrishnan, G. and R. Agrawal, 2014. Antioxidant activity and fatty acid profile of fermented milk prepared by Pediococcus pentosaceus. J. Food Sci. Technol., 51: 4138-4142.
CrossRefDirect Link - Ebringer, L., M. Ferencik and J. Krajcovic, 2008. Beneficial health effects of milk and fermented dairy products-review. Folia Microbiol., 53: 378-394.
CrossRefDirect Link - Fitzgerald, R.J. and B.A. Murray, 2006. Bioactive peptides and lactic fermentations. Int. J. Dairy Technol., 59: 118-125.
CrossRefDirect Link - Farvin, K.S., C.P. Baron, N.S. Nielsen and C. Jacobsen, 2010. Antioxidant activity of yoghurt peptides: Part 1-in vitro assays and evaluation in ω-3 enriched milk. Food Chem., 123: 1081-1089.
CrossRefDirect Link - Fleet, G.H. and M.A. Main, 1987. The occurrence and growth of yeasts in dairy products. Int. J. Food Microbiol., 4: 145-155.
CrossRefDirect Link - Roostita, R. and G.H. Fleet, 1996. Growth of yeasts in milk and associated changes to milk composition. Int. J. Food Microbiol., 31: 205-219.
CrossRefDirect Link - Viljoen, B.C., A. Lourens-Hattingh, B. Ikalafeng and G. Peter, 2003. Temperature abuse initiating yeast growth in yoghurt. Food Res. Int., 36: 193-197.
CrossRefDirect Link - Li, Y., T.J. Liu and G.Q. He, 2015. Antioxidant activity of peptides from fermented milk with mix culture of lactic acid bacteria and yeast. Adv. J. Food Sci. Technol., 7: 422-427.
Direct Link - Jakobsen, M. and J. Narvhus, 1996. Yeasts and their possible beneficial and negative effects on the quality of dairy products. Int. Dairy J., 60: 755-768.
CrossRefDirect Link - Gadaga, T.H., A.N. Mutukumira and J.A. Narvhus, 2000. Enumeration and identification of yeasts isolated from Zimbabwean traditional fermented milk. Int. Dairy J., 10: 459-466.
CrossRefDirect Link - Fitrotin, U., T. Utami, P. Hastuti and U. Santoso, 2015. Antioxidant properties of fermented sesame milk using Lactobacillus plantarum Dad 13. Int. Res. J. Biol. Sci., 4: 56-61.
Direct Link - Son, S. and B.A. Lewis, 2002. Free radical scavenging and antioxidative activity of caffeic acid amide and ester analogues: Structure-activity relationship. J. Agric. Food Chem., 50: 468-472.
CrossRefDirect Link - Tamime, A.Y. and R.K. Robinson, 2007. Tamime and Robinson's Yoghurt Science and Technology. 3rd Edn., Woodhead Publishing Limited, Cambridge, UK., Pages: 791.
Direct Link - Gobbetti, M., 1998. The sourdough microflora: Interactions of lactic acid bacteria and yeasts. Trends Food Sci. Technol., 9: 267-274.
CrossRefDirect Link - Lindgren, S.E. and W.J. Dobrogosz, 1990. Antagonistic activities of lactic acid bacteria in food and feed fermentations. FEMS Microbiol. Rev., 7: 149-163.
PubMedDirect Link - Charalampopoulos, D., R. Wang, S.S. Pandiella and C. Webb, 2002. Application of cereals and cereal components in functional foods: A review. Int. J. Food Microbiol., 79: 131-141.
CrossRefDirect Link - Papagianni, M., 2012. Metabolic engineering of lactic acid bacteria for the production of industrially important compounds. Comput. Struct. Biotechnol. J., Vol. 3.
CrossRefDirect Link - Ortigosa, M., A. Arizcun, A. Irigoyen, M. Oneca and P. Torre, 2006. Effect of Lactobacillus adjunct cultures on the microbiological and physicochemical characteristics of Roncal-type ewe's-milk cheese. Food Microbiol., 23: 591-598.
CrossRefDirect Link - Wilkowska, A., D. Kregiel, O. Guneser and Y.K. Yuceer, 2015. Growth and by‐product profiles of Kluyveromyces marxianus cells immobilized in foamed alginate. Yeast, 32: 217-225.
CrossRefDirect Link - Lange, N. and A. Steinbuchel, 2011. β-carotene production by Saccharomyces cerevisiae with regard to plasmid stability and culture media. Applied Microbiol. Biotechnol., 91: 1611-1622.
CrossRefDirect Link - Virtanen, T., A. Pihlanto, S. Akkanen and H. Korhonen, 2007. Development of antioxidant activity in milk whey during fermentation with lactic acid bacteria. J. Applied Microbiol., 102: 106-115.
CrossRefPubMedDirect Link - Osuntoki, A. and I. Korie, 2010. Antioxidant activity of whey from milk fermented with Lactobacillus species isolated from Nigerian fermented foods. Food Technol. Biotechnol., 48: 505-511.
Direct Link - Chen, J., H. Lindmark-Mansson, L. Gorton and B. Akesson, 2003. Antioxidant capacity of bovine milk as assayed by spectrophotometric and amperometric methods. Int. Dairy J., 13: 927-935.
CrossRefDirect Link - Unal, G., S.N. El, A.S. Akalin and N. Dinkci, 2013. Antioxidant activity of probiotic yoghurt fortified with milk protein based ingredients. Ital. J. Food Sci., 25: 63-69.
Direct Link