
Novirman Jamarun
Faculty of Animal Science, Andalas University, Padang, Indonesia
Mardiati Zain
Faculty of Animal Science, Andalas University, Padang, Indonesia
Arief
Faculty of Animal Science, Andalas University, Padang, Indonesia
Roni Pazla
Faculty of Animal Science, Andalas University, Padang, Indonesia
Pakistan Journal of Nutrition
Year: 2017 | Volume: 16 | Issue: 6 | Page No.: 393-399







ABSTRACT
Objective: The aim of this study was to evaluate the effects of calcium (Ca), phosphorus (P) and manganese (Mn) supplementation during Oil Palm Frond (OPF) fermentation by Phanerochaete chrysosporium on rumen fluid characteristics and Microbial Protein Synthesis (MPS). Materials and Methods: This study was conducted using a randomized block design with 3 treatments (the addition of Ca, P and Mn) and 5 replications. The following treatments were performed: T1 = 2000 ppm Ca+1000 ppm P+150 ppm Mn, T2 = 2000 ppm Ca+1500 ppm P+150 ppm Mn and T3 = 2000 ppm Ca+2000 ppm P+150 ppm Mn. The data were analyzed using one-way analysis of variance (ANOVA) followed by Duncan’s multiple range test for comparisons among the treatment means. The parameters measured were as follows: pH, Volatile Fatty Acids (VFAs) (mM), NH3 (mM), total protozoa (cells mL–1) (log 10), total bacteria (cells mL–1) (log 10) and MPS (mg mL–1). Results: The results revealed that VFA levels were significantly increased by the addition of P in T3. However, T1, T2 and T3 non-significantly affected (p>0.05) pH, NH3, total protozoa, total bacterial and MPS. Conclusion: The supplementation of OPFs with 2000 ppm Ca, 2000 ppm P and 150 ppm Mn resulted in the best rumen fluid characteristics and MPS.
PDF Abstract XML References Citation
Received: January 31, 2017;
Accepted: March 30, 2017;
Published: May 15, 2017
Copyright: © 2017. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Novirman Jamarun, Mardiati Zain, Arief and Roni Pazla, 2017. Effects of Calcium (Ca), Phosphorus (P) and Manganese (Mn) Supplementation during Oil Palm Frond Fermentation by Phanerochaete chrysosporium on Rumen Fluid Characteristics and Microbial Protein Synthesis. Pakistan Journal of Nutrition, 16: 393-399.
DOI: 10.3923/pjn.2017.393.399
URL: https://scialert.net/abstract/?doi=pjn.2017.393.399
DOI: 10.3923/pjn.2017.393.399
URL: https://scialert.net/abstract/?doi=pjn.2017.393.399
INTRODUCTION
The oil palm plant is valuable crop in many parts of the world. Indonesia is one of the largest palm oil producers worldwide, with a production capacity of 33.5 million tons from a total area of 11,672,861 ha1. This number will continue to increase with the increase in the world demand for palm oil. The oil palm plant produces three main types of waste that can be used as animal feed: Oil palm fronds, sludge palm oil and palm kernel cake. This waste is highly abundant throughout the year but its use as livestock feed is not maximized, especially on farms. Oil palm plantations can produce waste palm fronds at approximately2 8,880 kg month–1 ha–1.
Oil palm fronds have a high lignin content (30.18%), which limits their digestibility in the rumen of cattle3,4. Lignin is the fraction of the non-carbohydrate complex that is difficult to overhaul and that binds to cellulose and hemicellulose. The bonds between lignin and cellulose (lignocellulose) and between lignin and hemicellulose (lignohemicellulose) should be broken in advance so that cellulose and hemicellulose can be optimally utilized by rumen microbes.
Fermentation technology using the mold Phanerochaete chrysosporium has been proven to reduce the lignin content in palm fronds. Febrina et al.3 reported that 25.77% decrease in the lignin content with fermentation by Phanerochaete chrysosporium and supplementation with the minerals Ca and Mn. Phanerochaete chrysosporium is a model in the development and understanding of the ligninolytic enzyme production system because this fungus produces higher quantities of ligninolytic enzymes than do other strains5.
The growth and ligninolytic enzyme activity of Phanerochaete chrysosporium is influenced by the availability of minerals, such as Ca and Mn6, in the substrate. Additions of the minerals Ca at 2000 ppm and Mn at 150 ppm in the fermentation of OPFs by Phanerochaete chrysosporium were reported to maximize the growth of molds and enzyme activity, thereby increasing the digestible fiber fraction7. In addition to Ca and Mn, P is another mineral that is required for the growth of all microbial cells and for cell metabolic processes. The supplementation of P in low-quality feed for ruminant livestock can improve the fermentation process in the rumen8-10.
The processes of OPF fermentation by Phanerochaete chrysosporium as a source of energy and microbial protein synthesis are difficult to separate because the growth of rumen microbes is supported by feed degradation in the rumen. If the requirements of microbial growth are met and create conditions conducive to ruminal activity, the population and the number of microbes would be optimal for accelerating feed degradation11.
This study aimed to evaluate the effects of Ca, P and Mn supplementation during OPF fermentation by Phanerochaete chrysosporium on rumen fluid characteristics and microbial protein synthesis.
MATERIALS AND METHODS
For this study, the fermentation of OPFs was conducted at the Laboratory of Technology for Feed Industries, Andalas University and the rumen fermentability analysis was conducted at the Laboratory of Dairy Cattle Nutrition, Faculty of Animal Science, Bogor Agricultural Institute, from August through October, 2016. The variables measured in this study were the characteristics of rumen fluid (pH, NH3 and VFA), total bacteria, total protozoa and microbial protein synthesis.
OPF fermentation: The palm fronds used as the raw material in this study were taken from the distal two-thirds of the OPF. The OPF substrates were cut, dried and finely milled. The Ca was obtained from CaSO4, P was obtained from KHPO4 and Mn was obtained from MnSO4.H2O. Phanerochaete chrysosporium was maintained on Potato Dextrose Agar (PDA) slants at 4°C, transferred to PDA plates at 37°C for 6 days and subsequently grown on OPFs mixed with rice bran. Equal amounts of OPF leaves and stems were used and then the mineral solution from Brook et al.12 was added. The fermentation process was initiated by adding water to the OPFs until the water level reached 70%, then, Ca, P or Mn were added, depending on the treatment. Observations were made every 120 h for 20 days.
Rumen fluid characteristics: The measured rumen fluid characteristics were as follows: Ruminal pH, NH3-N concentration and VFA concentration. Fermenter tubes were each filled with 0.5 g of sample and then 40 mL of buffer solution and 10 mL of fresh rumen fluid (ratio of 4:1) were added. After the tube received CO2, the tubes were closed with a rubber ventilator. Tubes were inserted into the fermenter shaker water bath at a temperature of 39°C and incubated for 4 h, followed by the analyses of pH, NH3 and VFA. The ruminal pH was measured using a pH meter. The NH3-N concentration was measured using the microdiffusion (Conway) method and the total VFA concentration was measured using the steam distillation method.
Rumen protozoa population: Measurements were carried out using a counting chamber. Up to 0.5 mL of rumen solution was fixed with 0.5 mL methyl green formalin saline (MFS) solution in tubes and thoroughly mixed13. Up to 0.1 mL of each sample was placed on the counting chamber (hemocytometer) using a pipette and a cover slip was placed over the sample. The protozoa were counted on the counter under a microscope at 40X magnification. Based on the number of protozoa obtained by the above counting procedure, the number per 1 mL of rumen content can be calculated using the following Eq. 1:
where, n is the number of protozoa in the counting chamber and d is the sample dilution
Aerobic bacteria population
Sample preparation: First, 1 mL samples of aseptic rumen fluid were pipetted into 9 mL of diluent (Butterfield's phosphate buffer) to obtain a 1:10 dilution and then homogenized (10–1 dilution).
Analysis of total bacteria: Analysis of total bacteria uses the Anoxomat system (MART Microbiology, B.V., Netherlands) which is equipment for cultivating microorganisms in an oxygen-depleted or CO2-enriched environment.
From the sample preparation (10–1 dilution), for subsequent dilutions (1 0–2, 10–3, 10–4 and so on as needed), 1 mL of the previous solution was added into 9 mL of diluent using a sterile pipette. Then, the solutions were homogenized via vortex. Next, 1 mL of any desired dilution was added to a sterile petri dish, this was replicated twice. Then, 15-20 mL of Nutrient Agar (NA) media was poured into a petri dish to assess the total bacteria in the given examples and samples. The medium was homogenized by twirling the cup such that the sample mixed. Next, the samples were frozen. The cups were inserted upside down into an Anoxomat jar to create aerobic conditions according to the instructions for the Anoxomat system. Then, the cup was incubated at 37°C for 48 h with the cup in the upside down position. The number of colonies on each petri dish between 25 and 250 colonies was counted. If the number of colonies were >250, it was recorded as too many to count (TMTC). Colony calculations were performed.
Microbial protein synthesis: Microbial protein synthesis was measured using a centrifuge and a spectrophotometer. The centrifuge used in this study was a Beckman J2-21 centrifuge, while the spectrophotometer used was a LW scientific spectrophotometer UV-200Rs.
To measure microbial protein synthesis, a 20 mL sample of rumen fluid was initially centrifuged at 400 rpm for 45 sec in order to separate the bacteria. The samples were then centrifuged at 408 rpm for 5 min in order to reduce the population of protozoa in the rumen fluid and to remove any remaining food particles.
Aliquots (rumen fluid that had been centrifuged at 408 rpm with a decreased number of protozoa and no food particles) were taken and 64.5% trichloroacetic acid (TCA) was added to each 10 mL sample (up to 2.5 mL per sample). The samples were then centrifuged at 15,000 rpm for 20 min, the supernatant was discarded and the obtained cells/sediment were retained and washed with distilled water. The cell/sediment precipitate was then centrifuged again at 15,000 rpm for 20 min. The supernatant was discarded and the cell/sediment was added to a 30 mL solution of NaOH (0.25 N). The precipitate was heated in boiling water for 10 min. The resulting supernatant was collected from each sample and 1 mL was used for the analysis of microbial protein according to Plummer14.
Experimental design and statistical analysis: The study was carried out using a randomized block design (3x5) with five replications (the addition of Ca, P and Mn as the treatments). The differences between the treatment means were analyzed using Duncan’s multiple range test with a confidence interval of 5% (p<0.05).
The following treatments were carried out:
T1 = 2000 ppm Ca+1000 ppm P+150 ppm Mn
T2 = 2000 ppm Ca+1500 ppm P+150 ppm Mn
T3 = 2000 ppm Ca+2000 ppm P+150 ppm Mn
RESULTS AND DISCUSSION
The analyses of the data for the rumen fluid characteristics and microbial protein synthesis due to the addition of Ca, P and Mn during the fermentation of OPFs by Phanerochaete chrysosporium are presented in Table 1 and 2.
The rumen pH value is the balance of interaction between the buffer capacity and the acidity or alkalinity of the fermentation products. The statistical analysis showed the addition of the minerals Ca, P and Mn had no significant effects (p>0.05) on the pH of rumen fluid. There were no differences in the average pH of each treatment (T1, T2 and T3) due to the balance between the production of VFA and NH3. The rumen fluid pH values in the study ranged from 6.74-6.76.
| Table 1: | Rumen fluid characteristics due to the addition of minerals during the fermentation of OPFs by Phanerochaete chrysosporium |
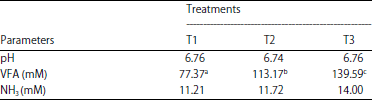 | |
| a-c Means in the same row with different letters are significantly different (p<0.05) | |
| Table 2: | Microbial protein synthesis due to the addition of minerals during the fermentation of OPFs by Phanerochaete chrysosporium |
 | |
| a-c Means in the same row with different letters are significantly different (p<0.05) | |
This value is within the normal range for the growth and development of rumen microbes. This demonstrated the addition of the minerals Ca, P and Mn during the fermentation of OPFs with Phanerochaete chrysosporium was able to maintain the rumen fluid pH levels. Church15 stated that a normal rumen pH range is from 6-7. The process of feed digestion will be disrupted if the pH of the rumen fluid is under 6. Maintenance of a suitable pH helps rumen microbes colonize the plant cell wall and can encourage bacterial cellulase activity to increase the production of VFA in normal amounts16.
Table 1 shows that treatment had no significant effect (p>0.05) on the concentrations of NH3 in the rumen fluid. The concentration of NH3 is one indicator that determines feed fermentability which is related to protein digestibility, rumen microbial activity and rumen microbial population17. The mean concentrations of NH3 in this study were 11.21-14.00 mM. Overall, it can be concluded that the availability of N-NH3 in the rumen fluid in the three treatments is within the normal range of NH3 availability. These values included the value for optimum rumen microbial growth needs, the concentration of ammonia needed to support microbial growth 6-21 mM18.
The T3 treatment resulted in the highest value of NH3, 14.00 mM. This indicated the quality of the protein in T3 was higher than the other treatments, the high concentrations of NH3 in this treatment showed the ability of microbes to degrade protein in feed was also better than the other treatments. Increasing the availability of NH3 in normal amounts will provide a balance of nitrogen and the energy required by the rumen microbes for growth. Erwanto et al.19 stated that the concentrations of NH3 in the rumen fluid can also determine the efficiency of microbial protein synthesis, which will ultimately affect the results of the fermentation of the organic materials of the feed in the form of VFAs, which are the main energy source in ruminants.
Volatile Fatty Acids (VFAs) are the main energy source in ruminant livestock feed and are produced via fermentation by rumen bacteria. The energy is used for the growth of the host animal and for the survival of the microorganisms. Based on the analysis of variance, the addition of minerals significantly increased (p<0.05) the total VFA production. Duncan’s multiple range test results showed the average total VFA production in treatment 3 was higher than those in T1 and T2 (Table 1). The average total VFA production ranged from 77.37-139.59 mM, while according to Waldron et al.20, the total rumen VFA concentration ranged from 60-120 mM. McDonald et al.18 suggested that the optimal VFA concentration for rumen microbe activity is 80-160 mM. This shows that in all treatments, the concentrations of total VFAs produced were sufficient for the growth of rumen microbes.
The T3 treatment showed the highest concentration of total VFAs due to the combination of the addition of the minerals Ca, P and Mn, this treatment is capable of optimally producing the ligninolytic enzymes and causing the release of lignin from cellulose and hemicellulose, thereby increasing the digestibility of the feed. The degradation of cellulose and hemicellulose produces glucose and oligosaccharides that were utilized by rumen microbes for growth and VFA production and high VFA production provides sufficient energy for livestock. The VFAs can be used for the synthesis of microbial protein to balance the production of NH3 and VFAs in the rumen can also be used as an energy source for ruminants21. The high concentration of total VFAs in the T3 treatment was also caused by increased fermentation due to the increasing number of rumen microbes (Table 2). The results obtained are also consistent with the increasing availability of NH3 in the rumen fluid, this allows microbes to grow and produce high levels of VFAs, resulting in a source of energy for microbial growth and development. Zain et al.9 and Febrina et al.10 reported that increasing the number of microbial cells would be able to increase the VFA production so that rumen microbes can grow efficiently. The VFA production in the rumen fluid can be used as a benchmark rate of the feed fermentability, where the higher the level of feed fermentability, the greater the VFA produced22.
The low total VFA in T1 was due to the low digestibility of nutrients and the low activity of the ligninolytic enzymes in Phanerochaete chrysosporium. This condition the rumen microbes, which cannot feed optimally, to degrade because the fermentation processes in the rumen are not optimal. This result showed that the lower the digestibility of the nutrient substrate, the lower the concentration of VFA produced. The low number of microbes in T1 will affect the total VFA production. The lower the number of cellulolytic rumen microbes, the lower the total VFA production. Fathul and Wajizah16 reported that the VFA content is the result of bacterial activity performing the activity of fermentation in the rumen, thus, more bacteria will produce more VFAs and vice versa.
Table 2 shows that the addition of the minerals Ca, P and Mn had no significant effects (p>0.05) on the populations of rumen bacteria and protozoa. This is because the treatments did not affect the pH of the rumen fluid. Uhi et al.23 reported that the changes in the populations of bacteria and protozoa were positively correlated with rumen fluid pH. The population of protozoa in this study ranged from 5.345-6295 cells mL–1 (log 10). Similar results were also reported by Febrina et al.10, the numbers of protozoa with the addition of the minerals P, S and Mg in OPFs fermented by Phanerochaete chrysosporium ranged from 5.851-5.864 cells mL–1 (log 10), which was lower than reported by Suryapratama and Suhartati24, ranged from 9.40-9.89 cells mL–1 (log 10).
The bacterial population in this study ranged from 9.060-9121 cells mL–1 (log 10), consistent with the rumen bacterial populations that were reported by Stewart25, where the rumen bacterial populations ranged from 109-1012 cells mL–1 of rumen content, which was equivalent to 9-12 cells mL–1 (log 10). The absence of treatment effects (p>0.05) on rumen bacterial populations due to the treatment also did not affect (p>0.05) the rumen protozoa population. This indicated the populations of protozoa and bacteria in the rumen are in a state of balance; a decrease in the number of protozoa would increase the total microbia and vice versa, given the presence of protozoa that prey on bacteria. The presence of a sufficiently high number of rumen bacteria is also related to ruminal pH conditions, which were ideal for cellulolytic bacterial activity.
The high total number of bacterial colonies in the T3 treatment was closely related to the content of the NH3 rumen fluid in T3, which was also the highest of any of the treatments. The NH3 was reclaimed by rumen microbes for growth, thus, the growth of rumen microbia depends on the availability of NH3 in the rumen fluid. Fathul and Wajizah16 reported that any increase in NH3 in the rumen fluid will increase the number of bacterial colonies because the number of bacterial colonies present in the rumen fluid was 45% dependent on the concentration of NH3 and 55% dependent on other factors.
The treatments had no significant effects (p>0.05) on MPS. The MPS value in this study ranged from 79,515-88,071 mg mL–1, with an average of 83,431 mg mL–1. The differences in MPS values between the treatments were within normal levels. The treatments were added to a basal diet with the addition of rice bran, cassava and wet and dry corn, which produce MPS that is consistent at approximately 68.21-72.93 mg mL–1. Sources of carbohydrates included fermentable carbohydrate groups that would be easily digested by rumen microbes26. Febrina et al.10 reported that the addition of minerals P, S and Mg in OPF fermentation did not significantly affect MPS (p>0.05), which remained at 36.878-46.816 mg mL–1.
Optimal MPS requires a supply of nutrients, nitrogen and acid. The supply of nitrogen comes from ammonia production and acid, while nutrient requirements are met from VFA production, which results from the fermentation of carbohydrates. Table 1 and 2 indicated the supplementation of minerals in T3 was able to increase the production of VFA, which was accompanied by the high number of microbes in the treatment. This indicated the amount of MPS increased due to the growing number of rumen microbes, with mineral supplementation causing an increase in energy availability. The high value of MPS with T3 indicated faster degradation of carbohydrates than the other treatments. The dissolving of the bonds in lignocellulose and lignohemicellulose with this treatment made nutrients more available and made carbohydrates in the rumen more easily degraded by rumen microbes. The rumen microbial population will increase when the availability of nutrients meets the needs of microbes and this will increase MPS. The high MPS in T3 suggested that the donation of mineral P generated from mold growth on OPFs also contributed to the availability of mineral P, which was optimal for the rumen microbes. The results of this study indicated the addition of the minerals Ca, P and Mn in the process of fermentation by Phanerochaete chrysosporium was able to improve the rumen bioprocess as demonstrated by the changes in the rumen fluid characteristics and microbial protein synthesis.
CONCLUSION
Supplementation of OPFs fermented by Phanerochaete chrysosporium with treatments of Ca (2000 ppm), P (2000 ppm) and Mn (150 ppm) (T3) provided the best rumen fluid characteristics and microbial protein synthesis.
SIGNIFICANCE STATEMENT
This study evaluated the effects of supplementing minerals Ca, P and Mn during the fermentation of oil palm fronds by Phanerochaete chrysosporium on rumen fluid characteristics and microbial protein synthesis, showing this supplementation can be beneficial for ruminants that consume forage-based waste from oil palm plantations. This study will help researchers uncover the critical functions of Ca, P and Mn supplementation in the process of fermentation by Phanerochaete chrysosporium, resulting in the full utilization of the nutrients in the palm fronds by the rumen microbes. This is a process that few researchers have been able to explore. Thus, this study may lead to a new theory on the optimal combinations of minerals for the fermentation process and their effects on rumen fluid characteristics and microbial protein synthesis.
ACKNOWLEDGMENTS
This research was funded by the Andalas University, Professor Grant 2016, contract No. 524/XIV/Unand-2016 and was part of the Ph.D. dissertation by Roni Pazla in Animal Nutrition, Faculty of Animal Science, Andalas University.
REFERENCES
- Febrina, D., N. Jamarun, M. Zain, Khasrad and R. Mariani, 2014. Biological delignification by Phanerochaete chrysosporium with addition of mineral Mn and its effect on nutrient content of oil palm frond. Proceedings of the 16th AAAP Animal Science Congress, November 10-14, 2014, Yogyakarta, Indonesia, pp: 1723-1726.
- Zain, M., J. Rahman and Khasrad, 2014. Effect of palm oil by products on in vitro fermentation and nutrient digestibility. Anim. Nutr. Feed Technol., 14: 175-181.
Direct Link - Singh, D. and S. Chen, 2008. The white-rot fungus Phanerochaete chrysosporium: Conditions for the production of lignin-degrading enzymes. Applied Microbiol. Biotechnol., 81: 399-417.
CrossRefDirect Link - Febrina, D., N. Jamarun, M. Zain and Khasrad, 2016. Effects of calcium (Ca) and manganese (Mn) supplementation during oil palm frond fermentation by Phanerochaete chrysosporium on in vitro digestibility and rumen fluid characteristics. Pak. J. Nutr., 15: 352-358.
CrossRefDirect Link - Zain, M., N. Jamarun and Zulkarnaini, 2010. Effect of phosphorus and sulphur supplementation in growing beef cattle diet based on rice straw ammoniated. Asian J. Scient. Res., 3: 184-188.
CrossRefDirect Link - Zain, M., N. Jamarun and A.S. Tjakradidjaja, 2010. Phosphorus supplementation of ammoniated rice straw on rumen fermentability, synthesised microbial protein and degradability in vitro. World Acad. Sci. Eng. Technol., 4: 357-359.
Direct Link - Febrina, D., N. Jamarun, M. Zain and Khasrad, 2016. The effects of P, S and Mg supplementation of oil palm fronds fermented by Phanerochaete chrysosporium on rumen fluid characteristics and microbial protein synthesis. Pak. J. Nutr., 15: 299-304.
CrossRefDirect Link - Brook, E.J., W.R. Stanton and A. Wall-Bridge, 1969. Fermentation methods for protein enrichment of cassava. Biotechnol. Bioeng., 11: 1271-1284.
CrossRefDirect Link - Ogimoto, K. and S. Imai, 1981. Atlas of Rumen Microbiology. Japan Scientific Societies Press, Tokyo, ISBN: 9784762202643, Pages: 231.
Direct Link - Waldron, M.R., F.N. Schrick, J.D. Quigley, J.L. Klotz, A.M. Saxton and R.N. Heitmann, 2002. Volatile fatty acid metabolism by epithelial cells isolated from different areas of the ewe rumen. J. Anim Sci., 80: 270-278.
PubMedDirect Link - Suryapratama, W. and F.M. Suhartati, 2012. Increasing rumen microbial protein synthesis with additional dietary substrate of Saccharomyces cerevisiae and soybean oil. Anim. Prod., 14: 155-159.
Direct Link