
Deyv Pijoh
Department of Animal Production, Sam Ratulangi University, Manado, Indonesia
Sri Supraptini Mansjoer
Department of Animal Science, Sam Ratulangi University, Manado, Indonesia
Dondin Sajuthi
Faculty of Mathematics, Bogor Agricultural University, Bogor, Indonesia
Dewi Apri Astuti
Department of Animal Science, Sam Ratulangi University, Manado, Indonesia
Irma Herawati Suparto
Faculty of Mathematics, Bogor Agricultural University, Bogor, Indonesia
Pakistan Journal of Nutrition
Year: 2017 | Volume: 16 | Issue: 5 | Page No.: 364-371







ABSTRACT
Objective: This study was conducted to develop an animal model of obesity by monitoring the phenotypes of long-tailed monkeys (Macaca fascicularis) fed a high energy diet. Methodology: Fifteen adult male Long-Tailed Monkeys (LTM) weighing between 4 and 5 kg were divided into three groups of five that received one of three diets: Diet A: A commercial monkey chow diet, Diet B: Monkey chow supplemented with egg yolk and Diet C: In which wheat was the main ingredient and beef tallow was added. Body measurements, including body weight were collected monthly. Results: The LTM fed Diet B (Egg yolk-supplemented) consumed more food compared to those fed Diet A and C. Animals in the Diet B group had significantly (p<0.01) larger waist circumference, hip circumference, chest circumference and skin fold thickness compared to the other two groups. There was a close relationship between body weight and waist circumference (r = 0.863), chest circumference (r = 0.889) and hip circumference (r = 0.890). Based on the main component analysis, LTM fed Diet B had higher rates of obesity compared to LTM fed Diet A or C. Conclusion: Body characteristics of LTM fed different diets can be used to determine the presence of obesity. The LTM can serve as a useful animal model of obesity and findings from this model can be applied to studies of obesity in humans.
PDF Abstract XML References Citation
Received: December 02, 2016;
Accepted: March 17, 2017;
Published: April 15, 2017
Copyright: © 2017. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Deyv Pijoh, Sri Supraptini Mansjoer, Dondin Sajuthi, Dewi Apri Astuti and Irma Herawati Suparto, 2017. Phenotype Development of Long Tailed Monkey (Macaca fascicularis) Fed Obese Diet. Pakistan Journal of Nutrition, 16: 364-371.
DOI: 10.3923/pjn.2017.364.371
URL: https://scialert.net/abstract/?doi=pjn.2017.364.371
DOI: 10.3923/pjn.2017.364.371
URL: https://scialert.net/abstract/?doi=pjn.2017.364.371
INTRODUCTION
Obesity is a global problem that affects more than 300 million people worldwide1. The percentage of individuals who are overweight or obese is increasing in line with dietary changes and decreased amounts of physical activity. Obesity is associated with metabolic syndrome, which is a major risk factor for several diseases, including diabetes, as well as cardiovascular, hypertension and immunological disorders2. Metabolic syndrome increases the incidence of cardiovascular disease by more than 20%3. The prevalence of metabolic syndrome is high among obese children and adolescents and the condition can become more severe without appropriate interventions. The causes and mechanisms of obesity need to be better defined and thus increased biomedical research of obesity is needed as well as the development of animal models that closely mimic human physiology and anatomy.
Primates of the Macaca genus are an appropriate animal model of obesity not only because of their anatomical, physiological and genetic similarities to humans but their size and long life span allow for longer sampling periods. Meanwhile, some studies used adult bonnet monkeys (Macaca radiata) to determine whether stress factors are a cause of obesity and the effect of insulin resistance in genetic studies of obesity4. It was observed that body mass index in long-tailed macaques (LTM, Macaca fascicularis) can influence expression of obesity-related genes5. Chen et al.6 showed that, in addition to genetic influences, the environment can also affect obesity.
Phenotype is manifested through the expression of genes that result in specific qualitative and quantitative characteristics of animals that can be seen and measured. Phenotypes related to body measurements are useful for quickly identifying obesity-related characteristics7,8. Currently there is limited information regarding the physical characteristics or phenotype of obese monkeys, which has presented as an obstacle for classifying obese and non-obese animals. Currently, the criteria for obesity in primate species are still based on those for humans. As such, this study aimed to obtain information on body dimensions that play a role in determining quantitative indicators of obesity in primates and how they can be applied to an animal model of obesity.
MATERIALS AND METHODS
Animals: A total of 15 adult male long-tailed monkeys (LTM) were used for this study. The LTM weighed between 4 and 5 kg and the animals were between 6 and 8 years-old. All LTM derived from captive-bred animals and were free of tuberculosis. All animal procedures and treatments used in this study were approved by the Bogor Agricultural University, Commission for Laboratory Animal Welfare or the Institutional Animal Care and Use Committee (IACUC; approval No: 02-IA-ACUC-08). The LTM were housed individually in 0.6×0.6×0.9 m stainless steel cages (back squeeze cage). Each cage was equipped with food and drink receptables that were affixed to the cage exterior. Animals were given environmental enrichment in the form of frozen fruits. The cages were positioned so that each animal could have audio-visual interactions with the other animals. The animals were divided into three groups that were fed three different diets.
Diets: The LTM in Group A were fed monkey chow consisting of 15, 5 and 3.49% protein, fat and crude fiber, respectively (Diet A). The Gross Energy (GE) content was 4,330 cal g–1. Treatment B was the Diet A monkey chow diet supplemented with egg yolk (Diet B). Diet B had 4,207 cal g–1 GE, 22% fat and 50.73% Nitrogen Free Extract (NFE). Group C received an obese diet formulated from high-energy materials including: Wheat 42%, sugar 8%, beef tallow (animal fat) 10%, cooking oil 10%, fish meal 4%, corn meal 8%, soybean meal 4%, rice bran 4%, jelly for cooking 1%, carboxymethyl cellulose (CMC) 1%, a mixture of minerals 2% (Diet C). Diet C had 4,480 cal g–1 GE, 21.7% fat and 52.34% NFE. The animals were fed the respective diets for 12 months.
Body measurements: The LTM body weight (kg) was measured monthly. The Body Mass Index (BMI) of the LTM was determined by dividing the weight (kg) by the square of the seated height (m2)9-11. The seated height or crown rump length is the distance between the top of the head to the base of the tail. Waistline Circumference (WC, cm) was measured at the midpoint between the peak of the iliac crest and the edge of the ribs on the center axillary line12. The thickness of skin folds on the stomach (cm) was measured just above the iliac bone using a caliper.
Statistical analysis: The data were subjected to ANOVA using 14:13 MINITAB software (State College, PA). To obtain the relationship between variables of experimental animal characteristics, the data were processed using Pearson correlation and Principal Component Analysis (PCU)13.
This study used a Completely Randomized Design (CRD). Treatments were Diet A, Diet B and Diet C and there were five replications for each treatment. Linear correlation analysis was used to characterize relationships between the variables measured. After data analysis, differences between the treatments were analyzed by a Tukey test with a level of confidence >95% indicating significance14.
RESULTS AND DISCUSSION
Food consumption and body weight: The amount of food consumed, weight gain and BMI for the different experimental groups was determined (Table 1). The monkeys consumed varying amounts of the different diets with animals fed Diet B having the highest average daily consumption rate (114.48 g per monkey) and Diet A the lowest (54.29 g per monkey). These differences were significant by ANOVA analysis. The difference in consumption may be due to favorable qualities imparted by the addition of egg yolk, such as enhanced flavor and color. Indeed, Muwarni15 found that the chemical properties of egg yolk added flavor to animals feeds, whereas a study by Pelletier et al.16 suggested that addition of egg yolk contributes an aroma that can stimulate appetite. Overall, McDonald et al.17 stated that other factors affecting food consumption are ambient temperature, taste and physiological status as well as the nutrient concentration, body weight and palatability. A report by NRC18 on the nutritional requirements of non-human primates stated that adult male LTM with a body weight of 5.7 kg consumed between 30.0 and 50.0 g per monkey day–1. Our results showed that the average daily dry matter intake of Diet B by the LTM exceeded this amount. The average consumption of dry matter feed by Group B was >90 g/monkey/day, an amount that can cause obesity in LTM, whereas the results of Astuti et al.19 showed that an increase in consumption fulfilled basic metabolic needs. The surplus food would presumably be stored as body fat.
The final body weight of the LTM at the end of the experiment ranged between 4.50 and 5.02 kg (Table 1). Those monkeys that received Diet B (chow+egg yolk) had the highest amount of weight gain and LTM fed Diet A (monkey chow) had the lowest, the differences were significant. The LTM fed monkey chow (Diet A) and the obese diet (Diet C) had slight differences in overall weight gain but these differences were not significant.
The BMI measurements ranged from 26.0-22.91 and the BMI of LTM fed Diet B was significantly different (p<0.05) from LTM that consumed Diet A and Diet C (monkey chow and obese diet, respectively (Table 1)). Monkeys that received Diet C (obese diet) qualified as overweight (23.63 kg m–2), whereas animals that were fed Diet B qualified as obese (26.0 kg m–2). In humans, the WHO1 classifies BMI values between 23.0 and 24.9 as overweight or pre-obese, whereas individuals with BMI values above 25.0 are considered to be obese. As noted by Putra et al. 20, obesity can be influenced by environmental and genetic factors10,21.
Measurement of LTM body dimensions: Obesity in LTM was also assessed in terms of changes in different body dimensions (Table 2). The mean Waist Circumference (WC) of the LTM during the 12 month study ranged between 28.54 and 32.97 cm (Fig. 1a).
| Table 1: | Average daily consumption, final weight and final body mass index of LTM fed different diets over a 12 month period |
 | |
A: Monkey chow (GE 4,330 kal g–1), B: Monkey chow supplemented with egg yolk (GE 4,207 kal g–1) and C: Obese diet with high energy lacking egg yolk (GE 4,480 kal g–1). Different superscript letters indicate significant differences | |
| Table 2: | Body dimensions of monkeys fed regular and obese diets |
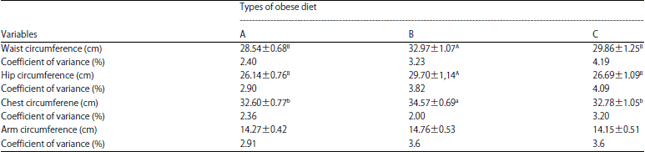 | |
A: Monkey chow (GE 4,330 kal g–1), B: Monkey chow supplemented with egg yolk (GE 4,207 kal g–1) and C: Obese diet with high energy lacking egg yolk (GE 4,480 kal g–1). Cova: Corelation of variance, Different superscript letters indicate significant differences | |
As with weight, LTM fed Diet B (chow+egg yolk) had a significantly higher WC compared to LTM fed Diets A and C, which had similar WC. Changes in abdominal skin fold thickness were consistent with those for WC, wherein animals fed Diet B had thicker skin folds relative to those fed Diets A and C (Fig. 2a).
The mean Hip Circumference (HC) of the LTM was also measured during the 12 month study. The values ranged between 29.70 and 26.14 cm and again the LTM fed Diet B with egg yolk had the highest value (29.70±0.20 cm) and LTM fed regular chow had the lowest at 26.14 cm. Results of variance analysis showed significant differences in HC between Diet B and Diets A and C (Table 2), which had small but not significant differences. Changes in HC across the 12 month period were also measured and were essentially constant throughout the study, although Diet B animals had higher values (Fig. 1b). These results are consistent with an earlier study showing that WC of monkeys increased concurrent with fat accumulation in the abdominal area, which can be classified as the central type of obesity22.
The mean Chest Circumference (CC) of LTM during the 12 month study period was between 32.57 and 34.57. As with WC and HC, the CC was highest for LTM fed Diet B contaning egg yolks and the lowest CC was seen for LTM given monkey chow (Table 2). The CC values for Diet B animals differed significantly from those for Diets A and C, which were similar (Table 2). Changes in CC over the study period also mirrored those seen for WC and HC (Fig. 1c). A 2009 study by Putra et al.20 reported that animals having a CC>34 cm met the criteria used for obesity.
The mean Arm Circumference (AC) of LTM at 12 months was between 14.15 and 14.76 cm and the three diet groups showed no significant differences in AC (Table 2). Changes in AC were also measured and were similar throughout the study period (Fig. 1d). The small change in AC is in agreement with findings of a study by Adam7 that showed there was little subcutaneous fat storage in the upper arm region and that storage of fat is closely associated with muscle tissue.
Measurement of skin fold thickness in LTM fed different diets: Measurement of skin fold thickness can be used to determine the amount of fat in the subcutaneous simian crease20. Thick skin folds can be associated with both genetic factors23 and environmental factors6. It can be measured that the thickness of skin folds on the stomach, as well as thickness of skin folds on the back, the rear of the arm and the forearm (Table 3).
| Table 3: | Skin fold thicknesses of monkeys fed different obese diets |
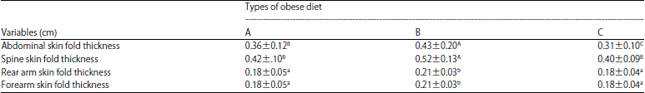 | |
A: Monkey chow (GE 4,330 kal g–1), B: Monkey chow supplemented with egg yolk (GE 4,207 kal g–1), C: Obese diet with high energy lacking egg yolk (GE 4,480 kal g–1). Different superscript letters indicate significant differences | |
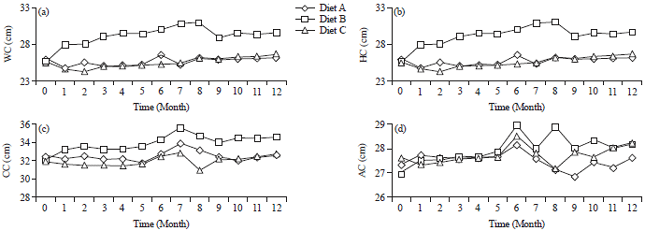 | |
| Fig. 1(a-d): | Changes over time in (a) Waist Circumference (WC), (b) Hip Circumference (HC), (c) Chest Circumference (CC) and (d) Arm Circumference (AC) of LTM fed different diets |
| A: Monkey chow (GE 4,330 kal g–1), B: Monkey chow supplemented with egg yolk (GE 4,207 kal g–1) and C: Obese diet with high energy lacking egg yolk (GE 4,480 kal g–1) | |
The LTM fed Diet B had the thickest abdominal skin folds, whereas the thickness of abdominal skin folds of LTM fed Diet A was higher than those for LTM fed Diet C. There were significant differences in fold thickness among all three diets (Table 3). Changes in skin fold thickness in the abdomen were also followed throughout the 12 month study period (Fig. 2a). The differences in the abdominal skin folds can reflect variations in subcutaneous fatty deposits in the abdominal region.
The thickness of skin folds in the back region had variations that were similar to those seen for the abdominal skin folds, in that LTM fed Diet B had significantly thicker back skin folds than LTM fed Diet A and C, which were similar (Table 2, Fig. 2b).
Meanwhile, the thickness of skin folds on the back of the arm and the forearm was highest for Diet B animals and Diet A and C animals had similar thicknesses (Table 2, Fig. 2c, d). Overall, Diet B animals had the largest changes in different regions of the body.
Phenotype correlation analysis variables: All phenotype variables were positively correlated with each other (Table 4). Analysis of these correlations, in which body weight was positively correlated to abdominal skin fold thickness and the thickness of folds on the skin on the back, hind legs, forelimbs, soles of the feet and palms, as well as the circumference of the back, waist, chest, arm, foot, thigh, calf and head. In this study, the LTM body weight was highly and positively correlated with waist circumference (r = 0.807; p<0.01), hip circumference (r = 0,849; p<0.01) and chest circumference (r = 0.701; p<0.01).
Main component analysis: The variant of the first main component (PC1) was 87.64, which explains that 83.6% was from the total variance, whereas the second variant of the main component (PC2) was 8.37, which explains that 8.0% was from the total variance. Together, PC1 and PC2 explains the observed data variance of 91.6% from the total variance. The primary component is the first high-seated PC1 = -0.033, -0.008 thickness fold of abdominal skin, -0.011 thickness of back skin folds, -0.003 thickness of back foot skin folds, -0.002 thickness of forelimbs skin folds, -0.008 thickness of soles, -0.007 thickness of palms, -0.512 hip circumference, chest circumference of 0.453, -0.150 of arm circumference, waist circumference of -0.197, - 0.640 of thigh circumference, head circumference of -0.199, -0,130 of calf circumference and -0.060 of body weight. The second major component of the equation is PC2 = -0.223 for seated height, abdominal skin fold thickness, 0.006 back skin fold thickness, -0.004 thickness of the skin folds on the back of the foot, -0.003 forelimb skin fold thickness, thickness of skin folds on the soles of the feet -0.015, -0.010 for the skin fold thickness on the palms, 0.230 waist circumference, chest circumference -0.291, -0.244 arm circumference, thigh circumference -0.312, 0.421 hip circumference, head circumference of -0.663, -0.178 of calf circumference, body weight of 0.082. The equation of PC1 and PC2 is based on the loading factor for each variable. The determination of the coefficient of the variable is arbitrary24.
Based on a study by Putra et al.20, the determination of the coefficient of the variable that is more than 0.4 can be used as a determinant of major components.
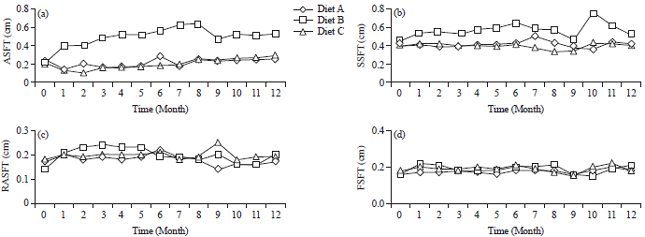 | |
| Fig. 2(a-d): | Changes over time in (a) Skin fold thickness in the stomach region (TSFS), (b) Skin fold thickness on the back (TSFB), (c) Skin fold thickness of the rear arm (TSFR) and (d) Skin fold thickness of the forelimbs (TSFF) of LTM fed different diets |
| A: Monkey chow (GE 4,330 kal g–1), B: Monkey chow supplemented with egg yolk (GE 4,207 kal g–1) and C: Obese diet with high energy lacking egg yolk (GE 4,480 kal g–1) | |
| Table 4: | Correlation of measurements after 12 months of feeding different diets |
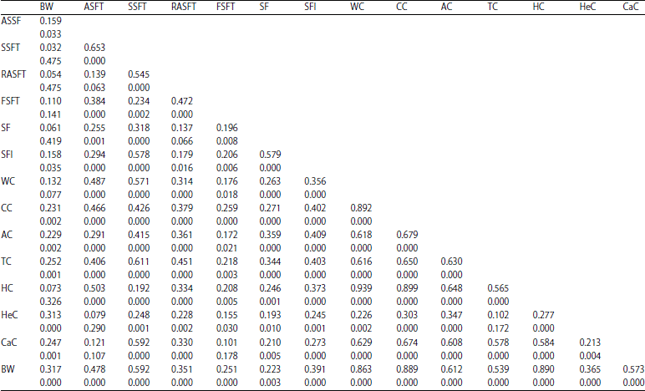 | |
BW: Body weight, ASFT: Abdominal skin fold thickness, SSFT: Spine skin fold thickness, RASFT: Rear arm skin fold thickness, FSFT: Forearm skin fold thickness, SF: Soles of foot, SFl: Soles of forlimbs, WC: Weist circumference, CC: Chest circumference, AC: Arms circumference, TC: Thight circumference, HC: Hip circumference, HeC: Head circumference, CaC: Calf circumference, BW: Body weight | |
Furthermore, the main component is determined by a minimum of three variables. Based on the PC1 equation, there are three variables that determine PC1, namely hip circumference, chest circumference and waist circumference with a coefficient (loading factor) of 0.512, 0.452 and 0.640, respectively. When linked with obesity, these three variables are highly influential in the classification of obesity in monkeys. Putra et al.20 further showed that other variables that can be used to categorize LTM are abdominal skin fold thickness, waist circumference and BMI.
Based on the results of the principal component analysis, LTM fed a diet containing high-energy egg yolk (Diet B) experienced greater body weight gain and could be categorized as obese. Only a small fraction of LTM fed monkey chow (Diet A) could be categorized as obese, whereas most were within the normal body weight range. The same conditions were seen for LTM fed Diet C (obese) in that only a small percentage of animals were obese.
Cheng et al.6 reported that the incidence of obesity increases under conditions where there is an imbalance between energy intake and expenditure. The excess energy that is not used is stored as body fat. Sediaoetama25 found that excessive consumption of fat leads to storage of fats as triglycerides that form a readily available energy supply. Excess fat intake can also lead to fatty deposits under the skin, around the abdominal organs and among muscle tissues 26 as shown by Frandson27 wherein triacylglycerol (TAG) in adipose tissue exists as potential energy reserves that are synthesized by anabolic lipogenesis processes. In conditions where nutrients are scarce, fat can be broken down through a process of β-oxidation to form fatty acids and glycerol.
Excess glucose can be converted to glycogen stored in the liver and muscle through glycogenesis. The ability of the liver and muscles to store glycogen is limited such that in conditions of blood glucose is overload, fat deposits will be generated through lipogenesis28. Although there are many methods by which obesity can be determined, one simple approach is to measure BMI, as was done in this study for animals fed monkey chow (Diet A), chow supplemented with egg yolk (Diet B) and an obese diet (Diet C). The LTM receiving the egg yolk-supplemented diet were most likely to have type 1 obesity characterized by excess abdominal fat.
CONCLUSION
| • | Monkeys fed obese diet containing egg yolk had a greater tendency to develop obeseity |
| • | Waist circumference, hip circumference and abdominal skin fold thickness can be qualitatively used as indicators of an obese phenotype in monkeys |
RECOMMENDATION
Developing techniques for obesity study can be used to help define biological properties such as behavior and physiology that are associated with obesity.
ACKNOWLEDGMENT
This study was supported by the Primate Research Center at Bogor Agricultural University and PT Wanara Satwa Loka. We thank our colleagues from Primatology Science at Bogor Agricultural University, whose insight and expertise greatly assisted this study.
REFERENCES
- Ford, E.S., 2005. Risks for all-cause mortality, cardiovascular disease and diabetes associated with the metabolic syndrome. Diabetes Care, 28: 1769-1778.
CrossRefDirect Link - Klein, S., N.F. Sheard, X. Pi-Sunyer, A. Daly, J. Wylie-Rosett, K. Kulkarni and N.G. Clark, 2004. Weight management through lifestyle modification for the prevention and management of type 2 diabetes: Rationale and strategies. A statement of the American Diabetes Association, the North American Association for the Study of Obesity and the American Society for Clinical Nutrition. Diabetes Care, 27: 2067-2073.
CrossRefPubMedDirect Link - Comuzzie, A.G., S.A. Cole, L. Martin, K.D. Carey, M.C. Mahaney, J. Blangero and J.L. VandeBerg, 2003. The baboon as a nonhuman primate model for the study of the genetics of obesity. Obesity Res. J., 11: 75-80.
CrossRefDirect Link - Putra, I.G.A.A., I.N. Wandia, I.G. Soma and D. Sajuthi, 2006. Body mass index and morphometry of the long tailed macaques (Macaca fascicularis) on Bali. J. Vet., 7: 119-124.
Direct Link - Chen, Y., H. Ogawa, H. Narita, K. Ohtoh, T. Yoshida and Y. Yoshikawa, 2003. Ratio of leptin to adiponectin as an obesity index of cynomolgus monkeys (Macaca fascicularis). Exp. Anim., 52: 137-143.
CrossRefDirect Link - Kaufman, D., M.A. Banerji, I. Shorman, E.L.P. Smith, J.D. Coplan, L.A. Rosenblum and J.G. Kral, 2007. Early-life stress and the development of obesity and insulin resistance in juvenile bonnet macaques. Diabetes, 56: 1382-1386.
CrossRefDirect Link - Angeloni, S.V., N. Glynn, G. Ambrosini, M.J. Garant, J.D. Higley, S. Suomi and B.C. Hansen, 2004. Characterization of the rhesus monkey ghrelin gene and factors influencing ghrelin gene expression and fasting plasma levels. Endocrinology, 145: 2197-2205.
Direct Link - Kemnitz, J.W., E.B. Roecker, R. Weindruch, D.F. Elson, S.T. Baum and R.N. Bergman, 1994. Dietary restriction increases insulin sensitivity and lowers blood glucose in rhesus monkeys. Am. J. Physiol. Endocrinol. Metab., 266: E540-E547.
Direct Link - Pelletier, X., P. Thouvenot, S. Belbraouet, J.A. Chayvialle, B. Hanesse, D. Mayeux and G. Debry, 1996. Effect of egg consumption in healthy volunteers: Influence of yolk, white or whole-egg on gastric emptying and on glycemic and hormonal responses. Ann. Nutr. Metab., 40: 109-115.
CrossRefDirect Link - NRC., 2003. Nutrient Requirements of Nonhuman Primates. 2nd Edn., National Academies Press, Washington, DC., ISBN: 9780309172042, Pages: 306.
CrossRefDirect Link - Astuti, D.A., I.H. Suparto, D. Sajuthi and I.N. Budiarsa, 2007. Nutrient intake and digestibility of cynomolgus monkey (Macaca fascicularis) fed with obese diet compared to monkey chow. Proceedings of the 2nd International Symposium on Food Security, Agricultural Development and Environmental Conservation in Southeast and East Asia, September 4-6, 2007, Bogor Agricultural University, Bogor, pp: 4-6.
- Rankinen, T., A. Zuberi, Y.C. Chagnon, S.J. Weisnagel and G. Argyropoulos et al., 2006. The human obesity gene map: The 2005 update. Obesity, 14: 529-644.
CrossRefDirect Link - Costello, A.B. and J.W. Osborne, 2005. Best practices in exploratory factor analysis: Four recommendations for getting the most from your analysis. Pract. Assess. Res. Eval., 10: 1-9.
CrossRefDirect Link - Howard, C.F.Jr. and M. Yasuda, 1990. Diabetes mellitus in nonhuman primates: Recent research advances and current husbandry practices. J. Med. Primatol., 19: 609-625.
PubMedDirect Link