
Mutiara Nugraheni
Department of Food Engineering Education, Faculty of Engineering, Yogyakarta State University, Karangmalang, Depok,
Sleman 55281 Yogyakarta, Indonesia
Badraningsih Lastariwati
Department of Food Engineering Education, Faculty of Engineering, Yogyakarta State University, Karangmalang, Depok,
Sleman 55281 Yogyakarta, Indonesia
Sutriyati Purwanti
Department of Food Engineering Education, Faculty of Engineering, Yogyakarta State University, Karangmalang, Depok,
Sleman 55281 Yogyakarta, Indonesia
Pakistan Journal of Nutrition
Year: 2017 | Volume: 16 | Issue: 5 | Page No.: 322-330







ABSTRACT
Background: Maranta arundinacea is a local tuber grown in Indonesia that is used as a source of carbohydrates. Maranta arundinacea flour modifications may increase levels of resistant starch, thereby increasing the functional properties of gluten-free flour. Materials and Methods: In this study, Maranta arundinacea flour was modified by 3-cycle autoclaving-cooling and then, three types of gluten-free enriched, resistant starch type 3 were identified, namely; high fiber, high protein and self-rising. These were evaluated in terms of chemical composition and ability to manage glucose, lipids and short-chain fatty acids in healthy mice. Results: The modified Maranta arundinacea flour increased the levels of resistant starch to 104.65%. The high-fiber flour was rich in dietary fiber as well as soluble dietary fiber (1.88±0.10%), insoluble dietary fiber (15.73±0.13%) and total dietary fiber (17.61±0.08%). The high-protein flour contained 14.43±0.02% protein. Self-rising flour was at ash level, indicating high levels of minerals. The consumption of the three types of gluten-free flour enriched with resistant starch type 3 from Maranta arundinacea flour kept glucose and lipids (total cholesterol, triglycerides, high density lipoprotein and low density lipoprotein) under normal conditions. The dominant short-chain fatty acids were acetate, propionate and butyrate. Conclusion: This study indicates that gluten-free enriched resistant, starch type 3 flour from Maranta arundinacea can help keep glucose and lipids under normal conditions, suggesting its potential use as a functional food, especially in people who have difficulties in managing glucose and lipid profiles.
PDF Abstract XML References Citation
Received: January 13, 2017;
Accepted: March 02, 2017;
Published: April 15, 2017
Copyright: © 2017. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Mutiara Nugraheni, Badraningsih Lastariwati and Sutriyati Purwanti, 2017. Proximate and Chemical Analysis of Gluten-free Enriched, Resistant Starch Type 3 from Maranta arundinacea Flour and its Potential as a Functional Food. Pakistan Journal of Nutrition, 16: 322-330.
DOI: 10.3923/pjn.2017.322.330
URL: https://scialert.net/abstract/?doi=pjn.2017.322.330
DOI: 10.3923/pjn.2017.322.330
URL: https://scialert.net/abstract/?doi=pjn.2017.322.330
INTRODUCTION
In 2016, Indonesia was ranked the 2nd largest wheat-importing country in the world1. Data from the United States Department of Agriculture indicates that Indonesia's wheat imports in 2016 reached 8.10 million tonnes or about 8% from the previous year, which was as much as 7.48 million tons1. With such imports, Indonesia became the 2nd largest wheat importer after Egypt, indicating that the need for flour in Indonesia has increased, although many processed wheat products are re-exported in other forms, such as instant and dried noodles1. The increase in wheat flour needs is paralleled by the development of large and small flour-based industries. Various efforts have been undertaken by the government to reduce its dependence on wheat flour, including the diversification of food products.
Food diversification efforts have not achieved the expected results2. Indonesia has the potential to produce food products derived from tubers, cereals and nuts3. One of the efforts to support the food diversification program to utilize the potential of Indonesia's food is the development of gluten-free flour4. Gluten-free flour consists of several carbohydrates, proteins and fats that can produce a food that has characteristics similar to flour. One local food that can be a source of carbohydrates is Maranta arundinacea.
Maranta arundinacea comes from tropical countries, such as Indonesia5 and is a local tuber commonly used as an ingredient for traditional foods. These tubers contain starch and other compounds and have a high amount of dietary fiber. Maranta arundinacea has various culinary uses in cookies, cakes, pudding, porridge, fruit pie fillings and glazes as a thickening agent and the Maranta arundinacea starch has high digestibility. Maranta arundinacea is bland, making it suitable for neutral diets, especially for people who have gastrointestinal illnesses, which is why many health food stores sell Maranta arundinacea cookies. Maranta arundinacea tubers contain 85.2% carbohydrate and 355 kcal energy and is often used as a substitute for carbohydrates6. Compared to other starches, Maranta arundinacea fibers have a shorter form so it is easy to digest and can be used as a food for babies, children with autism and down syndrome and patients in convalescence.
Carbohydrate sources can be physically modified for use by people who are sensitive to gluten, have celiac disease or require glucose and lipid control. Physical modifications have been carried out to increase levels of resistant starch with the autoclaving-cooling process; this process causes the starch to undergo retrogradation and increases levels of resistant starch7-9.
Maranta arundinacea flour is rich in resistant starch, which results in the high-selling prices of Maranta arundinacea flour-based products rich in resistant starch type 3 (RS3). Maranta arundinacea flour with rich RS3 can be used as a gluten-free flour ingredient as gluten-free flour enriched with RS3 from Maranta arundinacea flour can be consumed by gluten allergy sufferers.
Gluten has been identified as the trigger of the onset of type 1 diabetes as well as diabetic mellitus, both of which are rising in Indonesia. Increased levels of resistant starch are beneficial to control glucose and lipids. There is strong evidence that removal of gluten from the diet can selectively protect against the development of diabetes10,11.
This study aims to identify the potential of gluten-free enriched RS3 from Maranta arundinacea as a functional food by evaluating its chemical composition and its ability to control glucose, lipids and short-chain fatty acids in healthy mice.
MATERIALS AND METHODS
Materials: Maranta arundinacea flour was obtained from farmers in Clereng, Kulon Progo, Yogyakarta, Indonesia. Maranta arundinacea flour was made from Maranta arundinacea tuber. The manufacturing of Maranta arundinacea flour began with the selection of fresh tuber after washing in running water. The next step included slicing and drying using a dryer cabinet until the moisture content was 12%. Then, it was milled and sifted using a tyler sieve size mesh 80. All constituent ingredients for the gluten-free flour were obtained from the local market. Cholesterol kit, triglycerides kit, Low Density Lipoprotein (LDL) kit and High Density Lipoprotein (HDL) kit were purchased from Diasys Diagnostic System Holzheim Germany and the glucose kit was purchased from Diagnostic Bavaria Germany; all reagents were of analytical reagent grade.
Physical modification of Maranta arundinacea flour or Coleus tuberosus flour with 3 cycles of autoclaving-cooling: Maranta arundinacea flour or Coleus tuberosus flour rich in RS3 was made using a 3-cycle of autoclaving-cooling. Maranta arundinacea flour or Coleus tuberosus flour was suspended in water 20% (b/v). Then, it was heated at a temperature of 70°C and stirred until homogeneous. Maranta arundinacea flour or Coleus tuberosus flour suspension was then heated using an autoclave at 121°C for 15 min, resulting in perfectly gelatinized flour.
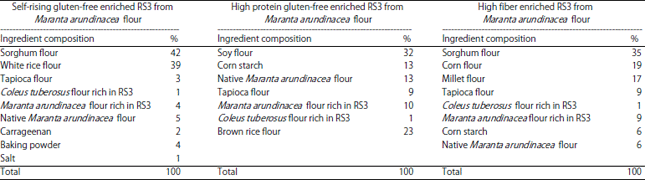
| Table 1: | Composition of three types of gluten-free enriched RS3 from Maranta arundinacea flour |
 | |
After refrigeration for 1 h, Maranta arundinacea flour or Coleus tuberosus flour paste was cooled at a temperature of 4°C for 24 h, autoclaving-cooling process was done as much 3 cycles, then dried using a dryer cabinet until reaching a moisture content of 12%. Finally, it was milled and sifted using Tyler sieve mesh 80. The RS3 levels can be improved by repeating autoclaving-cooling.
Formulation of gluten-free flour enriched RS3 from Maranta arundinacea flour: This study used three types of gluten-free flour: High-protein gluten-free flour, High-fiber gluten-free flour and self-rising gluten-free flour (Table 1). Gluten-free flour formulations were made by a combination of protein, fat and carbohydrates with raw materials that exist in Indonesia, which consist of tuber, cereals or legumes. The aim was to make gluten-free flour with sensory approach characteristics and sensory properties of wheat flour.
Proximate and chemical analysis: Proximate analysis referred to the established standards, including moisture content, ash content, the fat content by Soxhlet method, protein by Kjeldahl method12 and carbohydrate content by difference. Total starch content13, amylose and amylopectin and dietary fiber content were referred as AOAC (soluble dietary fiber and insoluble dietary fiber) and resistant starch8,12,14.
Animals and experimental diets: In vivo tests were conducted in the Laboratory Animal Experiment, Center for Food and Nutrition Studies, Gadjah Mada University from July to August of 2016. Twenty four male Wistar mice that were 3 months old were retrieved from the unit maintenance of animal experiments at the Center for Food and Nutrition Studies at Gadjah Mada University and taken as samples with a weight of 110-150 g. They were sacrificed under closed conditions with uncontrolled light enclosure and stable ventilation at room temperature. Standard feed was given for 3 days by using standard AIN15.
Mice were divided into four groups: Six mice were fed diet standards, six mice were a high-fiber gluten-free enriched RS3 diet from Maranta arundinacea flour, six mice were fed a high-protein gluten-free flour enriched RS3 diet from Maranta arundinacea flour and six mice were fed a self-rising gluten-free flour enriched RS3 diet from Maranta arundinacea flour. The feed pellets were given to the mice for 21 days of maintenance. Water was provided ad libitum. The cages were cleaned every day and the shelters were cleaned of dirt or stool. Residual feed was weighed every day and feed was given each morning.
Blood was collected from retro-orbitally from the anthus of the eye using capillary tubes under ether anesthesia16. Triglycerides were measured using the GPO-PAP method17. Total cholesterol was determined enzymatically using the CHOD-PAP method18. The LDL-cholesterol and HDL were determined enzymatically using the CHOD-PAP method19.
The SCFAS analysis was conducted using gas chromatography (GC) GC-8A Shimadzu Brand with a FID detector; a glass column of 10% GP SP 1200/1% H3PO4 on 80/100 chromosorb WAW column with length of 2 m and a diameter of 3 mm was used. The injector temperature was 220°C, the detector temperature was 220°C, the column temperature was 1300°C, the gas pressure was H2 = 0.6 kg cm–2, the water pressure was 0.5 kg cm–2 and the N2 was 1.25 kg cm–2. The carrier was N2 gas.
Statistical analysis: The results are presented as the average and standard deviation of the three experiments. One-way ANOVA was used to analyze differences in means between the samples followed by a least significant difference test. Multiple comparison tests were performed to compare the mean values at p<0.05. The value of p was considered significant at p<0.05. The SPSS version 16.0 (SPSS Inc., South Wacker Drive, Chicago, United State of America) was used.
RESULTS AND DISCUSSION
Maranta arundinacea flour modification by 3 cycle autoclaving-cooling: One of the goals of this study was to produce gluten-free flour enriched with Maranta arundinacea rich in RS3. There are many methods that can be developed to produce flour rich in RS3 either chemically, physically or enzymatically; however, in this study, we used to 3-cycle autoclaving-cooling to improve the levels of resistant starch type 3 physically. Some studies have proven that these physical methods can increase levels of RS3 on the raw materials that contain carbohydrates20,21. Maranta arundinacea flour was modified by 3-cycle autoclaving-cooling to increase the levels of resistant starch.
Resistant starch is smaller than the conventional particle size of dietary fiber. It can be used to improve the appearance, texture and mouth feel of food products22. Resistant starch content in food can be increased by modifying the processing conditions, including temperature, number of heating and cooling cycles and water content. When the effect of multiple heating/cooling treatments on resistant starch content of Maranta arundinacea flour was studied, the mean RS content of the Maranta arundinacea flour increased from 0.43±0.01 to 0.88±0.01% after 3-cycle autoclaving-cooling (p<0.05).
This study indicates that the resistant starch content increased with the modification process (Table 2). Based on the results of this study, autoclaving process-cooling increased the levels of RS3 on Maranta arundinacea flour by 104.65% (Table 2).
Formulation of gluten-free enriched RS3 from Maranta arundinacea flour: Gluten-free flour processing was carried out by taking into account the source of carbohydrates, protein and fat by making use of local cereals, tubers and legumes. Three types of gluten-free flour have a composition that is similar to its characteristics (Table 3, 4). The high-fiber gluten-free flour had total dietary fiber of 17.61±0.08%, consisting of insoluble dietary fiber (15.73±0.13) and soluble dietary fiber (1.88±0.10) (Table 4). Insoluble fiber consists of cellulose, hemicellulose and lignin and can be found in grains and beans. The high-protein gluten-free flour had a protein content of 14.43±0.07% (Table 3). Meanwhile, the self-rising gluten-free flour had a level of ash of 4.19±0.02% (Table 3).
The other chemical compositions, such as moisture content, fat content and carbohydrates, followed the same tendency. The water content in food affects the long-term durability of the food against microbes, expressed as the water activity (Aw). Water content refers to the amount of free water that can be used by microorganisms for growth. The greater the water content, the greater the value of Aw, such that the weaker the grocery long-term durability against invading microorganisms, the shorter the shelf life. Table 3 shows that the water content of gluten free was below 14% and can be stored for a year. Determination of carbohydrates by differences yielded an estimation of the overall amount of carbohydrates, both simple and complex carbohydrates.
The water content of gluten free flour was below 14% (Table 3) and can be stored for a year. Food fiber levels of high-fiber gluten-free flour relate to the composition of the constituent ingredients, sorghum flour, namely flour, millet and corn flour, including high fiber-containing materials. Sorghum fiber-flour food consists of 5.2%, millet (2.7%)23 and fiber food corn starch of 7.3%24.
High-protein gluten-free flour is related to the composition of the constituent ingredients of flour, although the material containing most high-protein soy flour is flour and brown rice; soy flour contain25 41.7% protein and brown rice contains 7-14% protein26. Self-rising flour contains the highest levels of ash, which is related to the composition of the raw materials, namely baking powder, salt and carrageenan. Ash content indicates the amount of mineral content in the material. The maximum ash content of carrageenan was 35% (baking powder sodium bicarbonate). Table 4 shows the highest levels of resistant starch on high-fiber gluten-free flour. Resistant starch levels of RS2 and RS3 were detected in three types of gluten-free flour.
| Table 2: | Resistant starch content in native and 3-cycle autoclaving-cooling of Maranta arundinacea flour |
| Values are expressed as Mean±SD | |
| Table 3: | Proximate analysis of gluten-free enriched RS3 from Maranta arundinacea flour |
 | |
| Values are expressed as Mean±SD. Values followed by the different letters in the same column showed a significant difference | |
The type of resistant starch in all types of gluten-free flour was RS2, which is contained in raw materials, such as potatoes, bananas and some nuts27. The majority of the components were native flour/starch, not through processing that causes gelatinization, whereas the RS3 was in Maranta arundinacea flour with 3 cycles of autoclaving-cooling.
Starch is an important form of polysaccharides in plant tissue. It is stored in the form of granules in the chloroplasts of leaves and in the seeds and tubers22. The starch consists of amylopectin and amylose. The three types of gluten-free flour had different levels of amylose and amylopectin. High-fiber gluten-free flour had significantly higher amylose levels than high-protein gluten-free flour or self-rising gluten-free flour, whereas the amylopectin content of self-rising gluten-free flour was significantly higher than high-fiber gluten-free flour or high-protein gluten-free flour. These differences were affected by the composition of the gluten-free flours. Amylose content in starchy foods is classified into four groups: Very low amylose content of less than 10%, low amylose content of about 10-20%, medium amylose content of about 20-24% and high amylose content with a level higher than 25%28.
Based on the classification, the results of this study indicate that gluten-free flour is classified as a low amylose content starch. Amylose content in high-fiber gluten-free flour was higher than high-protein and gluten-free self-rising flour. Amylose has a long straight chain, making it more difficult to be degraded by enzymes compared to amylopectin, which has more branches. Amylose has a 1,4-α-glycoside bond of unbranched amylose, also making it harder to digest29. Therefore, amylose is more widely used as a raw material for resistant starch and high resistant starch content is correlated with high amylose content30. In general, amylopectin content is higher than amylose and amylopectin content in all types of gluten-free flour is higher than amylose. The modification did not alter amylose and amylopectin content but changed the structure of amylose to be more resistant. The hydroxyl group was replaced by an acetyl group. Therefore, the substitution did not increase the total content of amylose and amylopectin.
Profile of blood glucose and lipid in diets with standard and gluten-free flour: The blood glucose of mice fed a standard feed, high-fiber gluten-free flour enriched RS3 from Maranta arundinacea flour, high-protein enriched RS3 from Maranta arundinacea flour and self-rising gluten-free enriched RS3 from Maranta arundinacea flour was in the normal range, under 110 mg dL–1 for 21 days of treatment (Table 5).
The profile of total cholesterol in the diet of three types of gluten-free enriched RS3 from Maranta arundinacea flour was no higher than standard feed and total cholesterol levels were normal (<200 mg dL–1). Triglyceride levels of mice fed the three types of gluten-free flour tended to be higher than that of those fed the standard feed but were still in the normal levels (<145 mg dL–1).
The lipid profile (total cholesterol, triglycerides, LDL and HDL) of mice fed a standard diet, high in protein and high fiber, was normal. Nevertheless, LDL levels of mice fed a self-rising gluten-free diet were different compared to the other groups; however, it remained in the normal range (LDL<100 mg dL–1)31. The HDL levels of mice fed a diet of self-rising gluten-free flour were decreased compared to those fed the other three types of flour (Table 5). The normal HDL levels in this strain32 are 35-85 mg dL–1. This difference was caused by the difference in the composition of the gluten-free flour ingredients; HDL levels of mice fed self-rising flour were a little below normal.
Soluble or insoluble dietary fiber in food as well as existing resistant starch in the gluten-free flour, can affect glucose levels. Cellulose, hemicellulose and lignin contain insoluble dietary fiber as well as pectin, gum, mucilage, glucan and algae. Soluble dietary fibers reduce glucose levels after a meal, which improves insulin profiles. The increase of viscosity in the digestive tract affects the speed of absorption of glucose. Insoluble dietary fiber slows the gastric emptying process of soluble fiber, which can form a layer of water that does not move in the intestine and can reduce the absorption of sugar33.
The mechanisms behind the insoluble dietary fiber are peripheral and not limited to nutrient absorption. First, accelerated secretion of glucose-dependent insulin tropic polypeptide (GIP) occurs directly after the ingestion of an insoluble fiber. The GIP is an incertain hormone which stimulates the post-prandial insulin release. Second, insoluble fiber can result in reduced appetite and food intake, which may lead to a decreased caloric intake and BMI in the U.S. which is described in the obesity section of this study34. Third, short-chain fatty acids, namely acetate, propionate and butyrate via fermentation have been shown to reduce post-prandial glucose. Short-chain fatty acids may reduce blood glucose levels through competition in insulin-sensitive tissues.
Consumption of soluble dietary be reduces post-prandial glucose response after carbohydrate-rich meals as well as lowers total and LDL cholesterol levels35. These effects are likely to explain the viscous and/or gel-forming properties of soluble dietary fiber, which thereby slows gastric emptying and macro-nutrient absorption from the gut.
| Table 4: | Resistant starch, amylose, amylopectin and dietary fiber content in gluten-free enriched RS3 from Maranta arudinacea flour |
| Values are expressed as Mean±SD. Values followed by the different letters in the same column showed a significant difference | |
| Table 5: | Profile of glucose and lipids in mice with diet feed standards and three types of gluten free flour enriched RS3 from Maranta arudinacea flour |
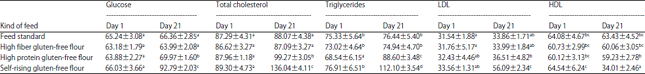 | |
| Values are expressed as Mean±SD. Values followed by the different letters in the same column showed a significant difference, LDL: Low density lipoprotein, HDL: High density lipoprotein | |
Highly fermentable dietary fiber increases glucagon-like peptides 1 levels and may play a role in the regulation of post-prandial satiety in diverse animal species36. Increased intake of total dietary fiber was inversely associated with markers of insulin resistance in several studies.
Dietary fiber from cocoa plays an important hypolipidemic role, returning triacylglycerol levels in hypercholesterolemic animals to normal values and reducing total and low density lipoprotein-cholesterol37. Regular inclusion of resistant starch in the diet positively regulates lipid levels38.
Total short-chain fatty acids profiles on high-protein and high-fiber gluten-free diets were no different than a standard diet (p<0.5). Meanwhile, in the self-rising flour diet, the amounts of acetate, propionate, butyrate and total SCFAs were much less lower compared to the standard feed and two types of gluten free flour (Table 6). Short-chain fatty acids, such as acetate, propionate and butyrate are produced by bacterial fermentation of insoluble dietary fiber in the colon, with the proportion of short-chain fatty acids not being different, depending on the substrate (the composition of the ingredient of gluten-free flour) and eventually, the gut microflora.
The presence of dietary fiber and resistant starch in the diet reduces the digestion and absorption of carbohydrates and increases the sensitivity of muscle and adipose tissue to insulin, allowing for greater gluconeogenesis, thereby contributing to the reduction of hyperglycemia39. Short-chain fatty acids are produced when resistant starch and dietary fibers undergo fermentation by colonic microbiota. Both the consumption of fermentable carbohydrates and the administration of short-chain fatty acids have been reported to result in health benefits, including improvements in glucose homeostasis, blood lipid profiles, body composition, reduced body weight and reduced colon cancer risk. Several mechanisms have been proposed by which resistant starch exerts beneficial effects. A major portion of the resistant starch is insoluble and fermented by gut microbes, which release short-chain fatty acids, such as butyrate, propionate and acetate40.
In previous studies, the short-chain fatty acids led to lipogenesis down regulation and increased lipid catabolism41,42. They have been linked to increased GLP-1 secretion in both animal and human models43-46. The GLP-1 is an incretin hormone that participates in glucose homeostasis by lowering plasma glucose concentration, improving insulin secretion and resistance and preserving pancreatic β-cell function47,48.
| Table 6: | Profile of short-chain fatty acid mice fed a standard diet and gluten-free diet enriched RS3 from Maranta arundinacea flour |
 | |
| Values are expressed as Mean±SD. Values followed by the different letters in the same column showed a significant difference, SCFAs: Short chain fatty acids | |
The GLP-1 is secreted by intestinal L cells, an open-type intestinal epithelial endocrine cell, in response to a variety of nutrients40,49,50.
Three types of short-chain fatty acids were studied here: Acetate, propionate and butyrate. Short-chain fatty acids (acetate and propionate) play a key role in lowering serum cholesterol and triglycerides51. Propionate is mainly metabolized in the liver and has been shown to inhibit gluconeogenesis and increase glycolysis in rat hepatocytes. It has also been proposed that propionic acid may lower plasma cholesterol concentrations by inhibiting hepatic cholesterogenesis52. Increased consumption of resistant starch produces more butyrate, which is associated with higher insulin sensitivity and improved glycemic control. The beneficial effects of butyrate on insulin sensitivity were related to the reduced fat deposition and lower levels of blood lipids, thereby, linking butyrate to improved insulin sensitivity through the mechanisms involving adipose tissues53.
CONCLUSION
Physical modification with 3-cycle autoclaving-cooling can increase levels of resistant starch type 3 in Maranta arundinacea flour compared to native Maranta arundinacea flour. The three types of gluten-free flour produced contained different characteristics. High-fiber gluten-free flour had high soluble dietary fiber, insoluble dietary fiber and total dietary fiber content. High-protein gluten-free flour had high protein content, whereas self-rising flour had high levels of ash and high levels of minerals. The three types of gluten-free flour positively impacted glucose, lipids and short-chain fatty acids at normal levels. These results indicate that gluten-free enriched RS3 flour from Maranta arundinacea can potentially be used as a functional food.
SIGNIFICANCE STATEMENTS
This study evaluated the proximate levels and chemical content of gluten-free flour enriched, resistant starch type 3 and its potential as a functional food. These findings will help other researchers develop gluten-free flour enriched resistant starch type 3 from Maranta arundinacea that has benefits for people with diabetes type 1 and 2 in the management of glucose and lipids and for those who are allergic to gluten.
ACKNOWLEDGMENT
The authors would like to thank the Directorate General of Higher Education of the Republic of Indonesia, with contract number 155/S-2H/LT/DRPM/III/2016, which has funded this study.
REFERENCES
- Chandrasekara, A. and T.J. Kumar, 2016. Roots and tuber crops as functional foods: A review on phytochemical constituents and their potential health benefits. Int. J. Food Sci., Vol. 2016.
CrossRefDirect Link - Shi, M.M. and Q.Y. Gao, 2011. Physicochemical properties, structure and in vitro digestion of resistant starch from waxy rice starch. Carbohydr. Polym., 84: 1151-1157.
CrossRefDirect Link - Ozturk, S., H. Koksel and P.K.w. Ng, 2011. Production of resistant starch from acid-modified amylotype starches with enhanced functional properties. J. Food Eng., 103: 156-164.
CrossRef - Nasrin, T.A.A. and A.K. Anal, 2014. Resistant starch III from culled banana and its functional properties in fish oil emulsion. Food Hydrocolloids, 35: 403-409.
CrossRefDirect Link - Marietta, E.V., A.M. Gomez, C. Yeoman, A.Y. Tilahun and C.R. Clark et al., 2013. Low incidence of spontaneous type 1 diabetes in non-obese diabetic mice raised on gluten-free diets is associated with changes in the intestinal microbiome. PloS One, Vol. 8.
CrossRef - Sildorf, S.M., S. Fredheim, J. Svensson and K. Buschard, 2012. Remission without insulin therapy on gluten-free diet in a 6-year old boy with type 1 diabetes mellitus. BMJ Case Rep., Vol. 2012.
CrossRef - Englyst, H.N., S.M. Kingman and J.H. Cummings, 1992. Classification and measurement of nutritionally important starch fractions. Eur. J. Clin. Nutr., 46: S33-S50.
PubMed - Reeves, P.G., F.H. Nielsen and G.C. Fahey Jr., 1993. AIN-93 purified diets for laboratory rodents: Final report of the American Institute of Nutrition ad hoc writing committee on the reformulation of the AIN-76A rodent diet. J. Nutr., 123: 1939-1951.
CrossRefPubMedDirect Link - McGowan, M.W., J.D. Artiss, D.R. Strandbergh and B. Zak, 1983. A peroxidase-coupled method for the colorimetric determination of serum triglycerides. Clin. Chem., 29: 538-542.
PubMedDirect Link - Allain, C.C., L.S. Poon, C.S.G. Chan, W. Richmond and P.C. Fu, 1974. Enzymatic determination of total serum cholesterol. Clin. Chem., 20: 470-475.
CrossRefPubMedDirect Link - Dundar, A.N. and D. Gocmen, 2013. Effects of autoclaving temperature and storing time on resistant starch formation and its functional and physicochemical properties. Carbohydr. Polymer, 97: 764-771.
CrossRefDirect Link - Yadav, B.S., A. Sharma and R.B. Yadav, 2009. Studies on effect of multiple heating/cooling cycles on the resistant starch formation in cereals, legumes and tubers. Int. J. Food Sci. Nutr., 60: 258-272.
CrossRefDirect Link - Sajilata, M.G., R.S. Singhal and P.R. Kulkarni, 2006. Resistant starch: A review. Compreh. Rev. Food Sci. Food Saf., 5: 1-17.
CrossRefDirect Link - Devi, P.B., R. Vijayabharathi, S. Sathyabama, N.G. Malleshi and V.B. Priyadarisini, 2014. Health benefits of finger millet (Eleusine coracana L.) polyphenols and dietary fiber: A review. J. Food Sci. Technol., 51: 1021-1040.
CrossRefPubMedDirect Link - Gwirtz, J.A. and M.N. Garcia‐Casal, 2014. Processing maize flour and corn meal food products. Ann. N. Y. Acad. Sci., 1312: 66-75.
CrossRefDirect Link - Gealy, D.R. and R.J. Bryant, 2009. Seed physicochemical characteristics of field-grown US weedy red rice (Oryza sativa) biotypes: Contrasts with commercial cultivars. J. Cereal Sci., 49: 239-245.
CrossRefDirect Link - Leszczynski, W., 2004. Resistant starch-classification, structure, production. Polish J. Food Nutr. Sci., 13: 37-50.
Direct Link - Shu, X., L. Jia, J. Gao, Y. Song, H. Zhao, Y. Nakamura and D. Wu, 2007. The influences of chain length of amylopectin on resistant starch in rice (Oryza sativa L.). Starch‐Starke, 59: 504-509.
CrossRefDirect Link - Sigit, S., H.P.E. Bimo Aksono, R. Damayanti, R. Bijanti, I. Herwiyarirasanta and H. Setyono, 2010. Effect of black soybean extract supplementation in low density lipoprotein level of rats (Rattus norvegicus) with high fat diet. Majalah Ilmu Faal Indonesia, 9: 217-221.
Direct Link - Torsdottir, I., M. Alpsten, G. Holm, A.S. Sandberg and J. Tolli, 1991. A small dose of soluble alginate-fiber affects postprandial glycemia and gastric emptying in humans with diabetes. J. Nutr., 121: 795-799.
PubMedDirect Link - Samra, R.A. and G.H. Anderson, 2007. Insoluble cereal fiber reduces appetite and short-term food intake and glycemic response to food consumed 75 min later by healthy men. Am. J. Clin. Nutr., 86: 972-979.
Direct Link - Lattimer, J.M. and M.D. Haub, 2010. Effects of dietary fiber and its components on metabolic health. Nutrients, 2: 1266-1289.
CrossRefDirect Link - Massimino, S.P., M.I. McBurney, C.J. Field, A.B. Thomson, M. Keelan, M.G. Hayek and G.D. Sunvold, 1998. Fermentable dietary fiber increases GLP-1 secretion and improves glucose homeostasis despite increased intestinal glucose transport capacity in healthy dogs. J. Nutr., 128: 1786-1793.
Direct Link - Lecumberri, E., L. Goya, R. Mateos, M. Alia, S. Ramos, M. Izquierdo-Pulido and L. Bravo, 2007. A diet rich in dietary fiber from cocoa improves lipid profile and reduces malondialdehyde in hypercholesterolemic rats. Nutrition, 23: 332-341.
CrossRefPubMedDirect Link - Birt, D.F., T. Boylston, S. Hendrich, J.L. Jane and J. Hollis et al., 2013. Resistant starch: Promise for improving human health. Adv. Nutr.: Int. Rev. J., 4: 587-601.
Direct Link - Robertson, M.D., A.S. Bickerton, A.L. Dennis, H. Vidal and K.N. Frayn, 2005. Insulin-sensitizing effects of dietary resistant starch and effects on skeletal muscle and adipose tissue metabolism. Am. J. Clin. Nutr., 82: 559-567.
PubMedDirect Link - Byrne, C.S., E.S. Chambers, D.J. Morrison and G. Frost, 2015. The role of short chain fatty acids in appetite regulation and energy homeostasis. Int. J. Obesity, 39: 1331-1338.
CrossRefDirect Link - Arora, T., R. Sharma and G. Frost, 2011. Propionate. Anti-obesity and satiety enhancing factor? Appetite, 56: 511-515.
CrossRefDirect Link - Delzenne, N.M., C. Daubioul, A. Neyrinck, M. Lasa and H.S. Taper, 2002. Inulin and oligofructose modulate lipid metabolism in animals: Review of biochemical events and future prospects. Br. J. Nutr., 87: S255-S259.
Direct Link - Yadav, H., J.H. Lee, J. Lloyd, P. Walter and S.G. Rane, 2013. Beneficial metabolic effects of a probiotic via butyrate-induced GLP-1 hormone secretion. J. Biol. Chem., 288: 25088-25097.
CrossRefDirect Link - Regmi, P.R., T.A.T.G. van Kempen, J.J. Matte and R.T. Zijlstra, 2011. Starch with high amylose and low in vitro digestibility increases short-chain fatty acid absorption, reduces peak insulin secretion and modulates incretin secretion in pigs. J. Nutr., 141: 398-405.
CrossRefDirect Link - Fukumori, R., T. Mita, T. Sugino, T. Obitsu and K. Taniguchi, 2012. Plasma concentrations and effects of glucagon-like peptide-1 (7-36) amide in calves before and after weaning. Domestic Anim. Endocrinol., 43: 299-306.
CrossRefDirect Link - Freeland, K.R. and T.M.S. Wolever, 2010. Acute effects of intravenous and rectal acetate on glucagon-like peptide-1, peptide YY, ghrelin, adiponectin and tumour necrosis factor-α. Br. J. Nutr., 103: 460-466.
CrossRefDirect Link - Holst, J.J., 2007. The physiology of glucagon-like peptide 1. Phys. Rev., 87: 1409-1439.
CrossRefPubMedDirect Link - Ross, S.A. and J.M. Ekoe, 2010. Incretin agents in type 2 diabetes. Can. Family Phys., 56: 639-648.
Direct Link - Kieffer, T.J. and J.F. Habener, 1999. The glucagon-like peptides. Endocrine Rev., 20: 876-913.
CrossRefDirect Link - Elliott, R.M., L.M. Morgan, J.A. Tredger, S. Deacon, J. Wright and V. Marks, 1993. Glucagon-like peptide-1 (7-36) amide and glucose-dependent insulinotropic polypeptide secretion in response to nutrient ingestion in man: Acute post-prandial and 24-h secretion patterns. J. Endocrinol., 138: 159-166.
CrossRefDirect Link - Herrmann, C., R. Goke, G. Richter, H.C. Fehmann, R. Arnold and B. Goke, 1995. Glucagon-like peptide-1 and glucose-dependent insulin-releasing polypeptide plasma levels in response to nutrients. Digestion, 56: 117-126.
CrossRefDirect Link - Henningsson, A., I. Bjorck and M. Nyman, 2001. Short-chain fatty acid formation at fermentation of indigestible carbohydrates. Naringsforskning, 45: 165-168.
CrossRefDirect Link - Gao, Z., J. Yin, J. Zhang, R.E. Ward, R.J. Martin and M. Lefevre et al., 2009. Butyrate improves insulin sensitivity and increases energy expenditure in mice. Diabetes, 58: 1509-1517.
CrossRefDirect Link