
Sarmin
Faculty of Veterinary Medicine, Universitas Gadjah Mada, Yogyakarta, Indonesia
LiveDNA: 62.15329
Irkham Widiyono
Faculty of Veterinary Medicine, Universitas Gadjah Mada, Yogyakarta, Indonesia
Pudji Astuti
Faculty of Veterinary Medicine, Universitas Gadjah Mada, Yogyakarta, Indonesia
Prabowo Purwono Putro
Faculty of Veterinary Medicine, Universitas Gadjah Mada, Yogyakarta, Indonesia
Pakistan Journal of Nutrition
Year: 2017 | Volume: 16 | Issue: 3 | Page No.: 101-108







ABSTRACT
Background: The ability of ruminants to undergo metabolic and endocrine adaptation to feeding restrictions and refeeding varies depending on the species and physiological characteristics, the age at which the feed restriction and refeeding are imposed, the rate and length of the feed restriction and refeeding, the physiological responses of various body parts to the feed restriction and refeeding conditions, sex and breed. Objective: The objective of this study was to evaluate the metabolic rates and endocrine responses of Kacang goats, Indonesian-bred ruminants, during the restricted feeding and refeeding periods. Methodology: Nine 1.5-2.5 year-old male goats were subjected to feed adaptation for the first 60 days. Then, the goats were subjected to three-level feed restriction for the second 60 days. The goats received 40, 50 or 60% feed restriction of dry fodder. The goats allocated to the 40, 50 or 60% feed restriction groups were referred to as group 1, 2 and 3, respectively. For the final 60 days, the goats underwent 100% refeeding with dry fodder. The weights of the ruminants were measured and blood samples were obtained at the end of the feed adaptation, feed restriction and refeeding. Results: At the end of the 60-day restriction, the three goats in group 3 were less than the baseline of Average Daily Gain (ADG), while the six other goats in groups 1 and 2 were still greater than the baseline of ADG, although there were no significant differences among the groups (p>0.05). The feed restriction increased lipolysis in adipose tissue and protein mobilization, non-esterified fatty acids (NEFAs) and albumin as the carrier of NEFA, creatinine and total protein in serum. Nonetheless, there were no differences among the three groups (p>0.05). Thyroid hormones (T3 and T4) as lipolytic hormones were ignored because their rates were lower than the baseline. Lipolytic responses in the form of High Density Lipoprotein (HDL) occurred in the three groups and there were significant differences among the three groups (p>0.05), indicating that triglycerides exported as Low Density Lipoprotein (LDL) were also not different among the three groups (p>0.05). This finding showed that there were no LDL secretion differences among the three groups. The two metabolic ketogenic effects, namely, Blood Urea Nitrogen (BUN) and glucose showed that BUN decreased to less than the baseline in group 2, although there were no significant differences among the groups (p>0.05). The glucose rate in group 2 was the highest among the groups (p<0.05), likely because of the low catabolism of labile protein deposits in group 2. After the refeeding was completed, the ADG in groups 2 and 3 increased significantly (p<0.05). In contrast, the ADG in group 1 did not increase significantly (p>0.05). Lipolysis continued to occur after the refeeding, with high rates of NEFA and albumin in the three groups without any differences among the groups (p>0.05). The lipolysis of hormones T3 and T4 was ignored, with both hormones remaining low. The rates of HDL and triglycerides exported as LDL were also constant and were not different among the three groups (p>0.05). Creatinine decreased significantly in the three groups (p<0.05) and there were no differences among the groups. The total protein increased significantly (p<0.5), but there were no differences among the three groups. The BUN metabolite did not change after refeeding was imposed on the three groups (p>0.05), although the BUN metabolite in group 1 tended to be higher than that in group 2. The glucose rate increased significantly in all three of the groups (p<0.05), although none of them reached the baseline. The ruminants in group 2 possessed the lowest glucose rate (p<0.05). Conclusion: A group of goats with a 50% restriction level of dry fodder tended to use ketones as the source of energy by suppressing protein degradation during feed restriction. The responses of ruminants to refeeding took the forms of continued lipolysis and increased protein degradation.
PDF Abstract XML References Citation
Received: October 03, 2016;
Accepted: December 21, 2016;
Published: February 15, 2017
Copyright: © 2017. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Sarmin, Irkham Widiyono, Pudji Astuti and Prabowo Purwono Putro, 2017. Metabolic and Endocrine Responses to Feed Restriction and
Refeeding in Kacang Goats. Pakistan Journal of Nutrition, 16: 101-108.
DOI: 10.3923/pjn.2017.101.108
URL: https://scialert.net/abstract/?doi=pjn.2017.101.108
DOI: 10.3923/pjn.2017.101.108
URL: https://scialert.net/abstract/?doi=pjn.2017.101.108
INTRODUCTION
There are 16.841 million goats in Indonesia. Most of these goats are native Indonesian goats locally called Kacang goats, which are found in dry land areas. Goats play significant and strategic roles in boosting the economic security of Indonesian farmers. Goat farming apparently helps Indonesian farmers to increase their incomes and living standards1. The breeders’ dependence on feed availability from nature has been the primary problem for goat production in Indonesia. The feed quantity and quality are influenced by the seasons2. In dry seasons, goats experience decreases in feeding consumption of rough fiber intake and real rough protein3. During this period, an appropriate strategy for feeding management is needed to minimize the impacts of feeding restrictions on livestock production. Livestock producers could choose the option of feed restriction methods and delay growth to the extent that a sufficient supply of good feed is available in the rainy season and utilize compensatory growth4. The ability of ruminants to cope with feed limitations was the result of a long evolutionary process in response to the natural fluctuations of feed availability5. Kacang goats’ adaptability to feed restriction and subsequently to continued refeeding has not yet been reported. Adaptation ability to feed restriction has become an external strategy for increasing production and economic efficiency and for avoiding high fat deposition at the end of the phase6,7. Therefore, this adaptation ability is utilized as a "Biostimulation method"8. The ability to adapt to feed restriction and refeeding varies and depends on the species and on physiological characteristics and age, as well as on the intensity and duration of restriction and refeeding, physiological responses of the body to the feed restriction and refeeding conditions, sex and breed9. Adaptation to feed restriction requires adjustments to fat deposition, metabolism rate, hormone regulation and substrate mobilization of metabolism fuel sources10. The weight gain caused by feed restriction and refeeding involves physiologic hormone mechanisms and very complex metabolites11 and relevant research on Kacang goats has not yet been reported. Accordingly, this study was designed to study the effects of 60 days feed restriction and 60 days refeeding on the ADG, metabolites and endocrine systems of male Kacang goats fed similar quantities and qualities of feed during each phase of the experiment.
MATERIALS AND METHODS
All methods of this study were approved by the Ethics Commission of the Integrated Research and Testing Laboratory, Universitas Gadjah Mada (LPPT UGM), under No. 202/KEC-LPPT/XI/2014.
Animals: Nine healthy male Kacang goats with weights of 20-25 kg and without anatomical defects aged 1.5-2.5 years were used in this study. Each goat was kept individually in the research stables owned by the Faculty of Veterinary Medicine, Universitas Gadjah Mada. Control of the animals’ health was undertaken regularly, including monitoring for the possibility of microbial infection and ectoparasitic and endoparasitic infestations.
Feeding: All the goats were fed with similar quantity and quality feed, which during feed adaptation and refeeding consisted of green fodder (rendeng, part of the top of the peanut plant) and concentrate (Gemuk A, PT Comfeed, Indonesia) with a feeding proportion of 60% green fodder and 40% concentrate, while during feed restriction, the ruminants were fed different quantities, according to the levels of feed restriction.
Research design: Nine goats were divided randomly into three groups (Group 1-3). Each group consisted of three goats. The research was performed in three phases. In the first phase, all of the goats underwent feeding adaptation for the first 60 days. All of the goats consumed similar quantities and qualities of feed starting on day 0 through the 60th day. The ruminants received full feeding to ensure that they possessed the same levels of similarity and comparability. On the 60th day, blood samples from all of the groups were obtained. During the second phase, all of the goats were subjected to feed restriction starting on the 61st-120th days. Group 1 received 40% dry fodder, group 2 received 50% dry fodder and group 3 received 50% dry fodder. Blood sample were obtained again on the 120th day. Weighing was performed every week during this phase. During the third phase, all of the goats received refeeding. The goats in all groups were fully provided with the same quantity and quality of feed from the 121st day until the 180th day. Weighing was performed every week during this phase, whereas, blood samples were obtained on the 180th day.
Sampling and blood analysis: Blood was drawn from the jugular vein and the separation of serum was immediately undertaken to investigate the chemical and clinic parameters of the blood. The assessment of blood serum chemical levels, such as total protein, albumin, triglycerides, High Density Lipoprotein (HDL), Low Density Lipoprotein (LDL), glucose, Blood Urea Nitrogen (BUN) and creatinine were performed using a Synchron CX® Clinical Systems spectrophotometer (Beckman Coulter Inc., Fullerton). The blood sampling and checking of blood chemical levels were performed using the standard methods described by Kraft and Duerr12. The metabolites that were measured, such as NEFA were determined by Max Discovery TM non-esterified fatty acids assay (Catalog No. 56, Bio Scientific, Austin, USA). The rate of thyroxine (T4) was determined by T4 ELISA (DRG® (Total thyroxine), EIA-1781, DRG International Inc., USA). The triiodothyronine level (T3) was determined by T3 ELISA (DRG® (Triiodothyronine), EIA-1780, DRG International Inc., USA).
Statistical analysis: The effects of feed restriction and refeeding were analyzed by ANOVA, while Duncan's Multiple Range Test (DMRT) was applied to show the difference between the treatments of feed restriction and refeeding and that there was a correlation of the percentage of treatment with the feed restriction and refeeding conditions. Correlations between the blood chemical levels or metabolites of each animal and ADG were analyzed with Pearson’s correlation coefficients.
RESULTS AND DISCUSSION
Effects of feed restriction: The results of blood and hormone analysis and ADG levels during the first phase were determined as the baseline. The data obtained from each individual goat during the feed restriction and refeeding phases were calculated based on the differences or deviations from the baseline. At the end of the 60 days restriction, group 3 was less than the baseline of ADG, whereas, groups 1 and 2 remained greater than the baseline, although there were no significant differences among the groups (p>0.05) (Fig. 1a).
Feed restriction resulted in a decrease in the glucose rate to less than the baseline for all the groups. Group 2 was the highest (p<0.05) among the groups (p>0.05) (Fig. 1b). The decrease of glucose increased lipolysis in the adipose and mobilization of proteins. Accordingly, NEFA (Fig. 1c) and albumin (Fig. 1d), as the carrier of NEFA, creatinine (Fig. 1e) and total protein (Fig. 1f) in the serum increased. Nonetheless, there were no differences among the three groups in these metabolites (p>0.05). The decrease in glucose rates encouraged the use of fatty acid as an alternative energy source in the body to fulfill the energy needs that were not met during the restriction phase13, then, there was an increase in lipolysis, which encouraged the increase in NEFA14. Lipolysis of adipose was controlled by hormone-sensitive lipase during the restriction phase15 in target tissues in the body. The increased NEFA concentration resulted in an increase in fatty acid absorption by the liver16 and free fatty acid then underwent re-esterification and oxidation17. Triglyceride was produced and then exported as VLDL18. Low triglyceride (Fig. 1g) occurred because the speed of hepatocytes in the triglyceride secretions as the result of NEFA synthesis was very slow, 25 times slower than that in mice19,20.
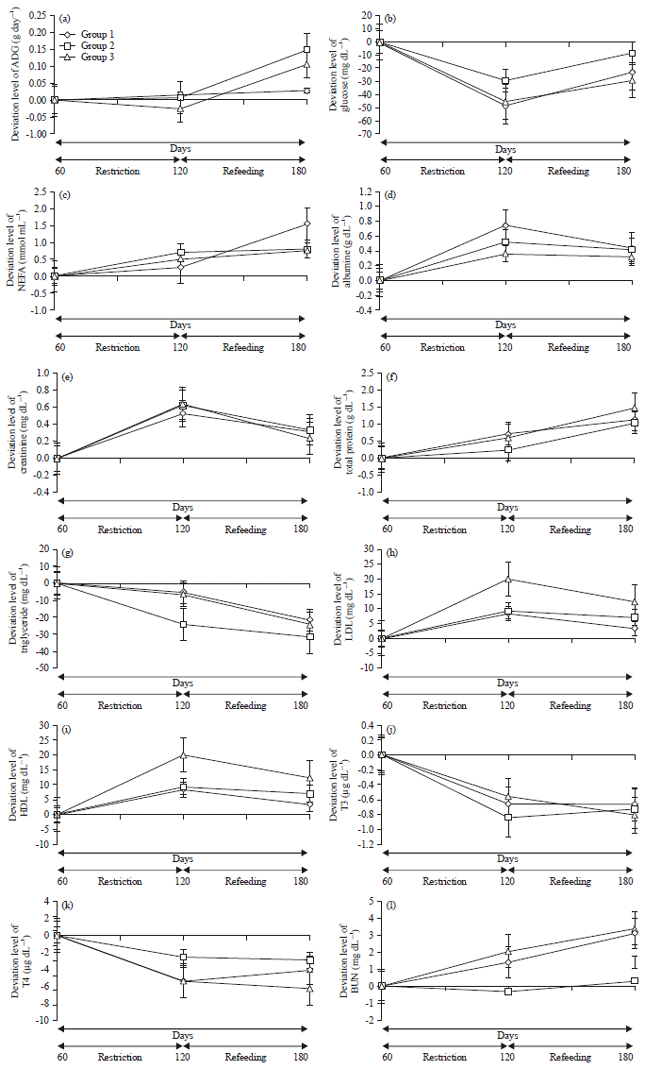 | |
| Fig. 1(a-l): | Deviation from the baseline of (a) ADG (g day–1), (b) Glucose (mg dL–1), (c) NEFA (mmol mL–1), (d) Albumin (g dL–1), (e) Creatinine (mg dL–1), (f) Total protein (g dL–1), (g) Triglyceride (mg dL–1), (h) LDL (mg dL–1), (i) HDL (mg dL–1), (j) T3 (μg dL–1), (k) T4 (μg dL–1) and (I) BUN (mg dL–1) at the feed restriction and refeeding phases of groups 1, 2 and 3 |
In addition, the ruminants’ ability to synthesize fatty acids was so low that efforts to boost the secretion of Low Density Lipoprotein (LDL) were needed. Accordingly, the uptake of free fatty acids by the liver increased21. The response to this increased LDL level did not differ among the three groups (p>0.05) (Fig. 1h). The increase in LDL secretion might have occurred as a response to the increased need for free fatty acid re-esterification in the cytosol of hepatocytes, although sometimes small amounts of free fatty acid were found to enter the mitochondria of hepatic cells. Lipolytic responses in the form of HDL increases occurred in the three groups and there were no significant differences among the three groups (p>0.05) (Fig. 1i), which indicated that triglyceride, exported as LDL as a result of HDL concentration increases, reflected an increase in VLDL turnover18,21. The role of high lipolysis was almost completely influenced by the energy deficit, whereas the role of the thyroid hormones T3 (Fig. 1j) and T4 (Fig. 1k) as lipolytic hormones were ignored because the rates of both hormone were low (less than baseline). This phenomenon was in accordance with the restriction phenomenon observed in Belgian Blue Bulls11.
In addition to NEFA, indications of energy reserve mobilization also arose from the increase in creatinine in all of the groups during feed restriction22. Creatinine is the end product of phosphocreatine breakdown and muscle creatine catabolism. Under normal metabolism conditions, phosphocreatine, a molecule of energy reserve, spontaneously undergoes a cycle to become creatine and inorganic phosphor as energy reserves in tissues. Creatine decomposes to form creatinine and systemic concentration of creatinine is related to weight and muscle mass. The increase in creatinine concentration during feed restriction in all of the experiment groups was in accordance with the results of feed restriction in Belgian Blue Bulls11,13,23. The increase in creatine concentration occurred because of the loss of muscle mass and protein catabolism13 to meet energy needs22. Due to the increase in energy needs as a result of low feed intake, creatine phosphorylation occurred in the muscle. Creatine produced in the liver, kidneys and pancreas was transported to the muscles to be metabolized. It was then converted to creatinine, as indicated by the creatinine increase in the blood22 and excreted as a calorie source14, as indicated by the BUN increase as a result of the glucose deficit. The BUN rates in the three groups were not different during the restriction phase (p>0.05), although a tendency was found in BUN to be less than baseline in group 2, whereas, the glucose rate of group 2 was the highest among the groups (p<0.05) (Fig. 1l). This outcome might have been caused by the tendency toward low catabolism of labile protein reserves in the body in group 2. The ruminants in group 2 tended to avoid protein degradation from hypoglycemia. In contrast, groups 1 and 3 increased protein degradation to boost gluconeogenesis mechanism. The difference in these two important metabolites in the metabolism of the three groups was caused by a difference in ketogenesis. The presence of ketones from the ketogenesis process increased the speed of protein degradation, causing gluconeogenesis to decelerate14,24 in groups 1 and 2. The real role of thyroid hormone in these goats was not in lipolysis but in efforts to increase the density of the kidneys, liver and cardiac ventricles by T3 hormone. This hormone also induced the relaxation of blood vessels, by which it decreased vascular resistance and increased renin and angiotensinogen activity. This hormone was also related to the creatinine increase as a result of decreases in the GFR or in the renin-angiotensin system. Both BUN and creatinine were related to thyroid hormone and their changes, which were associated with metabolism changes were the result of feed restriction25.
Effects of refeeding: After the refeeding phase ended, there was a significant increase in ADG in groups 2 and 3 (p<0.05) but not in group 1. The large increase in ADG in groups 2 and 3 showed that there was compensatory growth26,27. The recovery stage after refeeding was associated with a high lipid turnover and an increase in the lipolytic rate21,28, there was also a constant indication of high NEFA and albumin in the three groups, with no differences among the groups (p>0.05). The results of this study were in accordance with the refeeding phenomenon observed in Kamphaengsaen cows21. The high energy needs associated with recovery in addition to high lipid turnover, led to high utilization of glucose, as a result, the glucose rate never returned to baseline in any of the groups, even after the 60 days refeeding period. The glucose rate started increasing significantly in all of the groups but never returned to baseline (p<0.05). Group 2 was the lowest among the groups (p<0.05), perhaps because of the low catabolism of labile protein reserves in the body in group 2. Therefore, high lipolysis was a phenomenon found at refeeding28. The results of this study were in accordance with NEFA increasing after refeeding, as reported in Kamphaengsaen cows21. In bulls, it was reported that NEFA decreased to close to baseline after 31 days refeeding29 or 8 days13. The high rate of NEFA after refeeding was also reported in Kamphaengsaen cows21, apparently because of the high uptake of NEFA by peripheral tissue. Albumin as a NEFA carrier also increased29,30-32. The increase in albumin as a NEFA carrier also indicated the use of body protein reserves to fulfill energy needs13. A similar result was reported in cows33.
The time for NEFA to return to close to baseline differs depending on the treatment, species and efficiency rate of energy use at the beginning of refeeding due to low needs for energy maintenance during refeeding13. The effect of thyroid hormone as a lipolytic hormone11 seemed to be ignored while NEFA was elevated, since the differences in T3 and T4 were less than baseline despite having passed the refeeding phase. Nonetheless, it effected triglyceride since T3 was positively correlated with triglyceride (r = 0.69252, p<0.05) and T4 was negatively correlated with triglyceride (r = -0.76241, p<0.05).
This result was in accordance with that in Belgian Blue Bulls11. The decrease in T3 at the end of the restriction and refeeding phases in this study indicated a decrease in metabolic activity13, which occurred to decrease energy needs for maintenance34. Similar results have been reported in mice35, cows13,36 and Suffolk-cross goats37.
The HDL rate remained high, while in contrast, the triglyceride rate remained less than baseline and was exported as LDL, which also remained high and there were no differences among the three groups in the three metabolites (p>0.05). The low rate of triglyceride after refeeding showed that the gluconeogenesis process continued regardless the availability of amino acids in the body in the goats and led to cytosis36 in all of the experiment groups. The hepatocyte speed of the goats in secreting triglyceride from the synthesis of NEFA remained low 25% lower than that in mice-causing the triglyceride difference to be small during the refeeding phase19,20. In contrast, the high HDL and LDL at the end of refeeding showed the high use of lipids by the livers of ruminants21. A significant change was found in creatinine which declined increasingly significantly in the three groups (p<0.05). However, there were no differences among the three groups. The decrease in creatinine indicated a decrease in creatine phosphorylation because of the energy requirement was fulfilled after refeeding, showing the change in energy use from fat to protein. The decrease in creatine after refeeding was in accordance with refeeding in cows11,13.
The total protein increased significantly after refeeding (p<0.05), but there were no differences among the three groups. Urea-nitrogen metabolism did not change after refeeding in any of the groups (p>0.05), although groups 1 and 3 tended to have higher metabolism than group 2. The increase in total protein might have been caused by the lower use of nitrogen or lower retention of protein in the tissue13, although the levels in groups 1 and 3 tended to be higher than in group 2. The BUN metabolites did not change after refeeding in any of the groups (p>0.05) and there were no differences among the three groups (p>0.05). This result was different from that in Kamphaengsaen cows, in which urea nitrogen was lower after refeeding21 and from that in young cows29. This outcome probably occurred because the refeeding was associated with lipolysis and the constant rate of NEFA as a source of energy converted the carbon oxidation of glucose to alanine, by which the protein degradation of tissue decreased to produce glucose through the glucose-alanine cycle. Free fatty acid, which was used as an energy source was lower so that the protein degradation was still high, as indicated by the high rate of BUN. The increase in BUN during the refeeding phase also demonstrated a high synthesis mechanism in the liver as a result of ammonia production by microbes in the rumen, amino acid degradation in the liver11 because of high mobilization of proteins or increased protein/nitrogen intake after refeeding13, whether from feed concentrate or green feed (leaf fodder).
CONCLUSION
In response to 50% feed restriction of dry fodder, the goats tended to use ketones as sources of energy by reducing the degradation of protein during the restriction phase, while the responses to 40 and 60% restriction occurred in the form of breaking down high protein levels. The response to refeeding was due to the continuation of lipolysis and protein degradation tended to increase.
ACKNOWLEDGMENTS
The authors gratefully acknowledge the financial support of ‘Hibah Penelitian Unggulan Perguruan Tinggi 2014-2016 from Indonesia Directorate General for Higher Education (DIKTI), Ministry of Education and Culture, Republic of Indonesia.
REFERENCES
- Dahlanuddin, 2001. Forages commonly available to goats under farm conditions on Lombok Island, Indonesia. Livest. Res. Rural Dev., Vol. 13.
Direct Link - Chilliard, Y., F. Bocquier and M. Doreau, 1998. Digestive and metabolic adaptations of ruminants to undernutrition and consequences on reproduction. Reprod. Nutr. Dev., 38: 131-152.
PubMedDirect Link - Lovatto, P.A., D. Sauvant, J. Noblet, S. Dubois and J. van Milgen, 2006. Effects of feed restriction and subsequent refeeding on energy utilization in growing pigs. J. Anim. Sci., 84: 3329-3336.
CrossRefPubMedDirect Link - Mahmud, A., F.M. Khattak, Z. Ali, T.N. Pasha and U. Farooq, 2006. Early feed restriction, a tool to improve the feed efficiency in broilers. Pak. J. Biol. Sci., 9: 1178-1180.
CrossRefDirect Link - Daoud, N.M., A.H. Abd-El-Rahman, K.F. Mahrous, W.B. Khalil and O.H. Ezzo, 2012. GDF-9 gene expression, oocyte quality, hematological and biochemical profiles affected by feed restriction as a biostimulant method on rabbits' fertility. Global Veterinaria, 8: 532-540.
Direct Link - Yagoub, Y.M. and S.A. Babiker, 2009. 1. Effect of compensatory growth on performance of Sudanese female goats. Pak. J. Nutr., 8: 1802-1805.
CrossRefDirect Link - Fuglei, E., A.M. Mustonen and P. Nieminen, 2004. Effects of season, food deprivation and re-feeding on leptin, ghrelin and growth hormone in arctic foxes (Alopex lagopus) on Svalbard, Norway. J. Comp. Physiol. B, 174: 157-162.
CrossRefPubMedDirect Link - Hornick, J.L., C. van Eenaeme, M. Diez, V. Minet and L. Istasse, 1998. Different periods of feed restriction before compensatory growth in Belgian Blue bulls: II. Plasma metabolites and hormones. J. Anim. Sci., 76: 260-271.
CrossRefPubMedDirect Link - Keogh, K., S.M. Waters, A.K. Kelly, A.R.G. Wylie, H. Sauerwein, T. Sweeney and D.A. Kenny, 2015. Feed restriction and realimentation in Holstein-Friesian Bulls: II. Effect on blood pressure and systemic concentrations of metabolites and metabolic hormones. J. Anim. Sci., 93: 3590-3601.
CrossRefPubMedDirect Link - Klinhom, P., K. Markvichitr, P. Vijchulata, S. Tumwasorn, C. Bunchasak and A. Choothesa, 2006. Effect of restricted feeding on metabolic adaptations of Kamphaengsaen and crossbred Brahman heifers. Anim. Sci. J., 77: 399-406.
CrossRefDirect Link - Konigsson, K., G. Savoini, N. Govoni, G. Invernizzi, A. Prandi, H. Kindahl and M.C. Veronesi, 2008. Energy balance, leptin, NEFA and IGF-I plasma concentrations and resumption of post partum ovarian activity in Swedish red and white breed cows. Acta Vet. Scand., Vol. 50.
CrossRefDirect Link - Herdt, T.H., T. Wensing, H.P. Haagsman, L.M.G. van Gold and H.J. Breukink, 1988. Hepatic triacylglycerol synthesis during a period of fatty liver development in sheep. J. Anim. Sci., 66: 1997-2013.
CrossRefDirect Link - McNamara, J.P. and J.K. Hillers, 1986. Adaptations in lipid metabolism of bovine adipose tissue in lactogenesis and lactation. J. Lipid Res., 27: 150-157.
PubMedDirect Link - Kessler, E.C., J.J. Gross, R.M. Bruckmaier and C. Albrecht, 2014. Cholesterol metabolism, transport and hepatic regulation in dairy cows during transition and early lactation. J. Dairy Sci., 97: 5481-5490.
CrossRefPubMedDirect Link - Kleppe, B.B., R.J. Aiello, R.R. Grummer and L.E. Armentano, 1988. Triglyceride accumulation and very low density lipoprotein secretion by rat and goat hepatocytes in vitro. J. Dairy Sci., 71: 1813-1822.
CrossRefPubMedDirect Link - Pullen, D.L., J.S. Liesman and R.S. Emery, 1990. A species comparison of liver slice synthesis and secretion of triacylglycerol from nonesterified fatty acids in media. J. Anim. Sci., 68: 1395-1399.
CrossRefDirect Link - Klinhom, P., K. Markvichitr, P. Vijchulata, S. Tumwasorn, C. Bunchasak and A. Choothesa, 2006. Effect of refeeding on lipid metabolism in Kamphaengsaen beef heifers. Kasetsart J. (Nat. Sci.), 40: 420-429.
Direct Link - Cabaraux, J.F., M. Kerrour, C. van Eenaeme, I. Dufrasne, L. Istasse and J.L. Hornick, 2003. Different modes of food restriction and compensatory growth in double-muscled Belgian blue bulls: Plasma metabolites and hormones. Anim. Sci., 77: 205-214.
Direct Link - Holtenius, P. and K. Holtenius, 1996. New aspects of ketone bodies in energy metabolism of dairy cows: A review. J. Vet. Med. Ser. A, 43: 579-587.
CrossRefDirect Link - Tayal, D., R. Chawla, S. Arora, V.K. Gupta, J.S. Sohi and V. Mallika, 2009. Dynamic changes in biochemical markers of renal function with thyroid status-A study in Indian population. Internet J. Med. Update, 4: 36-41.
CrossRefDirect Link - Abouheif, M., H. Al-Sornokh, A. Swelum, H. Yaqoob and A. Al-Owaimer, 2015. Effect of different feed restriction regimens on lamb performance and carcass traits. Rev. Bras. Zootec., 44: 76-82.
CrossRefDirect Link - Dashtizadeh, M., M.J. Zamiri, A. Kamalzadeh and A. Kamali, 2008. Effect of feed restriction on compensatory growth response of young male goats. Iran. J. Vet. Res., 9: 109-120.
Direct Link - Dobrohorska, H., D. Sulima, K. Oskaldowicz, M. Gruszczynska and I. Golebiowska, 1981. Effect of refeeding after starvation on basal and tolbutamide-stimulated insulin secretion and beta-adrenergic receptor function in the regulation of insulin release and lipolysis in obese patients. Acta Physiologica Polonica, 32: 703-712.
PubMedDirect Link - Hayden, I.M., J.E. Williams and J.J. Collier, 1993. Plasma growth hormone, insulin-like growth factor, insulin and thyroid hormone association with body protein and fat accretion in steers undergoing compensatory gain after dietary energy restriction. J. Anim. Sci., 71: 3327-3338.
PubMedDirect Link - Kalscheur, K.F., B.B. Teter, L.S. Piperova and R.A. Erdman, 1997. Effect of fat source on duodenal flow of Trans-C18:1 fatty acids and milk fat production in dairy cows. J. Dairy Sci., 80: 2115-2126.
CrossRefDirect Link - Reynolds, C.K., P.C. Aikman, B. Lupoli, D.J. Humphries and D.E. Beever, 2003. Splanchnic metabolism of dairy cows during the transition from late gestation through early lactation. J. Dairy Sci., 86: 1201-1217.
CrossRefPubMedDirect Link - Rukkwamsuk, T., T.A.M. Kruip and T. Wensing, 1999. Relationship between overfeeding and over conditioning in the dry period and the problems of high producing dairy cows during the postparturient period. Vet. Quart., 21: 71-77.
CrossRefDirect Link - Blum, J.W., W. Schnyder, P.L. Kunz, A.K. Blom, H. Bickel and A. Schurch, 1985. Reduced and compensatory growth: Endocrine and metabolic changes during food restriction and refeeding in steers. J. Nutr., 115: 417-424.
PubMedDirect Link - Murphy, T.A. and S.C. Loerch, 1994. Effects of restricted feeding of growing steers on performance, carcass characteristics and composition. J. Anim. Sci., 72: 2497-2507.
Direct Link - Blake, N.G., D.J.A. Eckland, O.J.F. Foster and S.L. Lightman, 1991. Inhibition of hypothalamic thyrotropin-releasing hormone messenger ribonucleic acid during food deprivation. Endocrinology, 129: 2714-2718.
CrossRefPubMedDirect Link - Eski, F., I. Tasal, M.A. Karsli, S. Sendag, B.A. Uslu, H. Wagner and A. Wehrend, 2015. Concentrations of NEFA, β-HBA, triglycerides and certain blood metabolites in healthy colored Angora goats during the peripartum period. Turk. J. Vet. Anim. Sci., 39: 401-405.
CrossRefDirect Link - Ekpe, E.D. and R.J. Christopherson, 2000. Metabolic and endocrine responses to cold and feed restriction in ruminants. Can. J. Anim. Sci., 80: 87-95.
CrossRefDirect Link