
Paulus Klau Tahuk
Department of Animal Science, Faculty of Agriculture, Timor University, Kefamenanu, East Nusa Tenggara, Indonesia
Subur Priyono Sasmito Budhi
Faculty of Animal Science, Gadjah Mada University, Yogyakarta, Indonesia
Panjono
Faculty of Animal Science, Gadjah Mada University, Yogyakarta, Indonesia
Endang Baliarti
Faculty of Animal Science, Gadjah Mada University, Yogyakarta, Indonesia
Pakistan Journal of Nutrition
Year: 2016 | Volume: 15 | Issue: 10 | Page No.: 897-904







ABSTRACT
Objective: This study investigated in vitro characteristics of fermentation of fattening rations with different protein-energy levels fed to male Bali cattle. Methodology: Rations were composed of grass, Gliricidia sepium, corn meal and rice bran with different proportions of protein and energy. Ration T1 was standard protein-standard energy/SS [(12.06% Crude Protein (CP) and 62.66% Total Digestible Nutrients (TDN)], T2 was standard protein-high energy/SH (10.14% CP and 65. 66% TDN), T3 was high protein-standard energy/HS (14.79% CP and 63.66% TDN) and T4 was high protein-high energy/HH (13% CP and 67.48% TDN). Data were analyzed by one-way analysis of variance. Results: Although, pH level of rumen fluid was similar for all treatments (p>0.05), digestibility of dry matter and organic matter in rations T4 and T3 was higher (p<0.01) than that for T1 and T2. For N-NH3 (mg/100 mL), the yield of T4 and T3 was higher (p<0.01) than that for T1 and T2. Meanwhile, total VFA, acetic and propionic acids in T2, T3 and T4 were higher (p<0.01) than for T1 but butyric acid levels for T4 were higher (p<0.01) than that for T1, T2 and T3 and the acetic: propionic acid ratio of T4 was lower (p<0.01) than that for T1, T2 and T3. Microbial protein synthesis (mg mL–1) for the T1 ration was higher (p<0.01) than that of T2, T3 and T4. Conclusion: Varying the protein and energy levels of rations fed to male Bali cattle did not affect rumen pH but the digestibility of dry matter and organic matter was unclear. Moreover, N-NH3 utilization and VFA yield were not optimal for protein biosynthesis by microbes.
PDF Abstract XML References Citation
Received: January 18, 2016;
Accepted: May 12, 2016;
Published: September 15, 2016
How to cite this article
Paulus Klau Tahuk, Subur Priyono Sasmito Budhi, Panjono and Endang Baliarti, 2016. In vitro Characteristics of Rumen Fermentation of Fattening Rations with Different Protein-energy Levels Fed to Bali Cattle. Pakistan Journal of Nutrition, 15: 897-904.
DOI: 10.3923/pjn.2016.897.904
URL: https://scialert.net/abstract/?doi=pjn.2016.897.904
DOI: 10.3923/pjn.2016.897.904
URL: https://scialert.net/abstract/?doi=pjn.2016.897.904
INTRODUCTION
Protein and energy are two essential feed components that are needed to improve cattle productivity because feeds with high protein and high energy contents can accelerate fermentation rates and feed digestibility to increase nutrient intake by cattle. Low-quality feed, particularly feeds with low dry matter and organic matter digestibility, negatively affects feed consumption and metabolism in rumens, in turn can lead to inadequate protein and energy supplies for cattle. Feeds with higher digestibility are preferable given that some protein sources are positively correlated with degradation levels in the rumen1. In addition, feeds must also provide an adequate supply of nitrogen (ammonia) to support microorganism growth in the rumen. Cattle given high protein feed can metabolize the feed to produce high nitrogen-ammonia (N-NH3) yields which is important for protein microbe synthesis when adequate digestible carbohydrates from Volatile Fatty Acids (VFA) are available. Inadequate energy causes N-NH3 imbalances in cattle and in turn discharge of this compound from the body. It is therefore essential to achieve a balance between energy levels and protein digestibility in animal feeds.
In cattle rumens, microbial proteins contribute upto 40-80% of amino acids available for protein synthesis2. Accordingly, cattle production can be positively affected by improving protein microbe yield3 through approaches such as modulating the availability of nitrogen precursors and energy derived from fermentation4.
On smallholder farms with beef cattle livestock, feeds with adequate energy and protein levels are not always available to maintain maximum cattle performance throughout the year. Farmers often provide forage as the sole feed in fattening rations which has been shown to be ineffective in improving Bali cattle production. The energy value of a feed is primarily determined by the total proportion of Dry Matter (DM) intake or digestible energy and forage flow in the digestive tract. Thus, forage with higher digestibility provides more energy for cattle per unit DM consumed. Tahuk et al.5 reported that male Bali cattle in a West Timor, East Nusa Tenggara feedlot consumed 7.079 kg head–1 day–1 DM fattening forage to yield 0.321 kg head–1 day–1 daily weight gain which translates to a 23.664 feed conversion ratio and 4.619% feed efficiency. This low performance likely correlates with low digestibility and energy levels in forage indicating that field legume proteins could not optimize cattle performance. Moreover, Buxton and Brasche6 stated that forage as an energy supply for cattle is frequently a primary obstacle to herbivore productivity.
Considering the aforementioned circumstances, local feed with various protein and energy levels used by smallholder farms producing beef cattle is one alternative for resolving nutrient imbalances. Gliricidia sepium and natural grasses can be used as protein and structural carbohydrate sources, respectively. Furthermore, supplementation of this forage with highly digestible carbohydrates such as corn meal and rice bran would be expected to improve nutrient adequacy and balance for cattle, particularly by increasing energy levels that are known to increase animal growth, meat production and carcass weight7.
This study examined male Bali cattle fattening rations with different protein and energy levels by taking in vitro measurements of rumen fermentation parameters such as pH, dry matter, organic matter digestibility, N-NH3 yield, VFA yield and protein synthesis by microbes.
MATERIALS AND METHODS
Study location: In vitro study was conducted for 30 days at the laboratory of Animal Feed Technology, Department of Nutrition and Animal Feed, Faculty of Animal Science, Gadjah Mada University, Indonesia. The N-NH3 analysis and microbe protein synthesis measurements were carried out at the Nutrition Biochemical laboratory, Animal Science Faculty, Gadjah Mada University, Indonesia and VFA analysis was conducted at the Biochemical Chemistry laboratory, Food and Nutrition Centre, Gadjah Mada University, Indonesia.
Research design and feed rations: Research materials were forage commonly used by local farmers, comprising natural grass and Gliricidia sepium leaves with additional corn meal and rice bran in the rations. Research equipment included laboratory apparatus for feed proximate analysis and Van Soest analysis Acid Detergent Fiber (ADF) and Neutral Detergent Fiber (NDF) fractions, analysis of VFA, N-NH3 and protein synthesis by microbes) and a pH meter to measure ruminal fluid pH. Materials for in vitro analysis were fistular ruminal liquid from male Bali cattle, 39 °C McDougall solution pH 6.55-6.99, fresh 39°C ruminal fluid, 0.2% pepsin HCl, distilled water, saturated HgCl2, saturated NaCO3 solution, H2SO4 (0.005 N solution and boric acid with indicator), 5 N HCl solution, 15% H2SO4, 0.5 N NaOH and 0.1% phenolphthalein (PP) indicator solution.
Rations consisted of four materials: Natural Grass (NG), Gliricidia sepium (G), Corn Meal (CM) and Rice Bran (RB). Protein and energy levels of the rations were either those specified by standard cattle provisions or were varied to observe the effects on various parameters in vitro. The first ration treatment (T1) was prepared according to Kearl8 for 200 kg male cattle with 0.75 kg Average Daily Gain (ADG) and contained standard protein-standard energy (SS) consisting of 5.6 kg DM, 3.7 kg (59.26%) Total Digestible Nutrients (TDN) and 577 g (11.56%) crude protein. The experimental rations were as follows; (T2) standard protein-high energy (SH) with 12% crude protein and 70% TDN, (T3) high protein-standard energy (HS) with 15% crude protein and 60% TDN and (T4) high protein-high energy (HH) with 15% crude protein and 70% TDN. The chemical composition of the rations after proximate analysis changed slightly in protein and TDN proportion, wherein T1 was 12.06 and 62.66%, T2 was 10.14 and 65.66%, T3 was 14.79 and 63.66% and T4 was 13.04 and 67.48%, respectively. This post proximate analysis composition was subjected to in vitro evaluation.
Variables, research method and data collection: The variables analyzed in this study included ruminal fluid pH, digestibility (dry matter and organic matter), Volatile Fatty Acid (VFA) yield (including acetic, propionic and butyric acids), N-NH3 yield and microbial protein synthesis.
To measure the initial pH of ruminal fluid, samples collected from male Bali cattle fistules were filtered through a three-layered cloth to avoid contamination with feed residue and then stored in a thermos warmed to approximately 37°C. The collected rumen fluid was transported to the laboratory, prepared and incubated in a water bath according to the method described by Tilley and Terry9. Data collection complied with the following procedures:
| • | Ruminal fluid pH was measured with a Hanna pH meter. Measurements were taken soon after collection in order to record the initial pH during the first 48 h and to monitor pH changes |
| • | Feed digestibility (dry matter, organic matter and protein) was measured using the method described by Tilley and Terry9 |
| • | Volatile Fatty Acid (VFA) yield including acetic, propionic and butyric acid was determined by gas chromatography using the method described by Filipek and Dvorak10. Briefly, a Shimadzu GC8 (standard 1 µL) gas chromatograph equipped with a Flame Ionisation Detector (FID, detector temperature 240°C) and a 3 m GP 10% SP 1200/10% H3P04 column running at 140°C was used for gas chromatography. The carrying gas was N2 (nitrogen), the gas and hydrogen pressures were 1.8 and 0.8 kg cm–3, respectively |
Measurement procedure: Ruminal fluid taken with a stomach tube was centrifuged at 3,000 rpm for 25 min. Aliquots (0.2 mL) of the supernatant were stored in sealed Eppendorf tubes before 1 mL, 25% metaphosphoric acid was added. The mixture was centrifuged at 15,000 rpm for 15 min and 1 μL of the supernatant was injected for gas chromatography; a standard VFA solution was injected prior to injecting the sample:
| • | The N-NH3 yield was analyzed by spectrophotometry according to the method of Chaney and Marbach11. In brief, solution A (1 mL, Tungstat) with 2 mL ruminal liquid was mixed and combined with 1 mL cold solution B. This sample was frozen for ≤48 h before analysis. The samples were then centrifuged at 15,000 g for 10 min. Solutions C and D (2.5 mL) were then added to 20 mL supernatant and incubated in a 40°C water bath for 30 min until the solution turned blue. Samples were cooled and the absorbance values were determined at 630 nm |
| • | Microbial protein synthesis in the rumen was measured spectrophotometrically based on the method by Lowry et al.12. Samples were first centrifuged at 3,000 g for 15 min and 1.5 mL of the resulting supernatant was further centrifuged at 10,000 g for 15 min. The supernatant was then removed and 0.5 mL distilled water was added to the precipitate. The mixture was centrifuged at 10,000 g for 15 min. The supernatant was removed, 1 mL distilled water was added and diluted as necessary. To determine protein content, 1 mL of the sample solution was mixed with 5 mL Lowry B reagent and incubated at room temperature for 10 min before 0.5 mL Lowry solution A was added. The solution was incubated for 30 min and the absorbance at 750 nm was determined |
Statistical analysis: The data were collected using a one-way completely randomized design with four treatments and six replicates. The data were analyzed using SPSS software version 19 and one-way analysis of variance. When differences among treatments were suspected, the Duncan’s Multiple Range Test (DMRT) was carried out13.
RESULTS AND DISCUSSION
Acidity level: The acidity level (pH) of the ruminal liquid from animals given the four different protein-energy level rations did not differ significantly. Overall, the ruminal liquid pH was relatively low due to the utilization of 19, 65, 2 and 43% of total digestible carbohydrate feed contained in T1, T2, T3 and T4, respectively. Although, the use of higher amounts of easily digestible carbohydrate (concentrate) would be expected to lower the rumen pH, there was non-significant decrease in pH among the four ration treatments because the forage used still contained high fiber which would neutralize decreases in pH. Even as the dry matter and organic matter digestibility increased, the rumen pH remained within the optimum range for bacterial growth and activity as well as for maximal cellulose digestion (6.4-6.8). Indeed, Erdman14 stated that decreases in rumen pH in cattle could result from an acid-base imbalance and restricted metabolism rather than feed content.
Dry matter and organic matter digestibility: Dry matter and organic matter digestibility (%) of rations fed to beef cattle in this study was relatively high (~65.06-74.67%) and the digestibility of T2 and T4 which had higher TDN values was higher (p<0.01) than that of T1 and T3. The increase in Dry Matter Digestibility (DMD) and Organic Matter Digestibility (OMD) in rations T2 and T4 showed that rations with different protein and energy levels derived from easily digestible carbohydrates provided optimum energy (ATP) for rumen microbes to degrade feed. Corn meal also played an essential role in DMD and OMD as well as providing an adequate energy source for rumen microbe growth and activity. McDonald et al.15 reported that highly soluble carbohydrates (non-structural) such as glucose, fructose and starch were optimal for digestion and metabolism and could be best used by several bacterial species in the rumen.
Rations T1 and T3 contained easily digestible carbohydrate contents of 19 and 2%, respectively and yielded relatively low DMD and OMD that were correlated with energy availability and low digestibility that in turn reduces the ability of rumen microbes to degrade feed. Carbohydrates are a main energy source for bacteria16 although, they can also be used as a carbon frame for protein synthesis combined with ammonia. This study found that increases in DMD and OMD showed a linear relationship with the amount of digestible carbohydrate present in the rations as an energy source (Table 1).
Nitrogen-ammonia (N-NH3) yield: The Nitrogen-ammonia (N-NH3) yield (mg/100 mL) in the T1 and T2 rations which had lower crude protein contents was lower than that for T3 and T4 (p<0.01). These differences in N-NH3 levels could be attributed both to the ration type and degradation rate. As mentioned above, rumen pH was not expected to negatively affect N-NH3 production as the pH values seen here were within the range for optimal microbial growth and development as well as for feed degradation needed for protein synthesis.
The N-NH3 yield in cattle fed the T1 ration was relatively low compared to the other ration treatments because the N-NH3 arising from degradation of proteins in the feed was supported by adequate energy levels from digestible carbohydrate that could be used for protein microbe synthesis. Accordingly, nitrogen and carbohydrate degradation were synchronized and positively impacted rumen N-NH3 utilization.
In contrast, for ration treatment T2 (standard protein levels with high energy levels) the N-NH3 yield tended to increase which indicated that the N-NH3 yield was not optimized despite an adequate supply of easily digestible carbohydrates. A lack of nitrogen-energy feed synchronization due to different feed degradation rates may also have contributed to this condition.
| Table 1: | Composition of chemicals, DMD and OMD (%) of fattening rations with different protein-energy levels fed to Bali cattle |
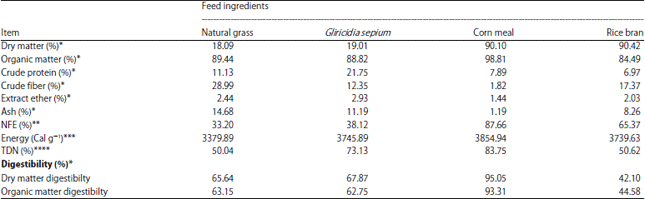 | |
*Analysis results from the Animal Feed Laboratory Technology, Faculty of Animal Science, Gadjah Mada University (2015), **NFE: [100–(Ash%+CF%+EE%+CP%)], ***Results Analysis of Laboratory Chemistry and Biochemistry, Study Center Food and Nutrition, Gadjah Mada University (2015), ****According to the equation Harris et al.17 quoted in Hartadi et al.18 | |
Corn meal and rice bran are easily digestible carbohydrate sources that have a higher degradation rate than feeds where Gliricidia sepium leaves are used as the nitrogen source. As such, when carbohydrate digestion reached optimum levels, nitrogen degradation rates for N-NH3 production remained sub-optimal. The higher crude protein level in the T3 ration (14.79%) contributed to the high N-NH3 concentration but the energy (TDN) was 63.66% and the amount of easily digestible carbohydrate was 2%. This composition led to faster protein degradation from the feed but did not provide adequate energy from digestible carbohydrates. Consequently, the N-NH3 derived from nitrogen feed degradation was not optimal for microbial protein synthesis. However, the complete amino acid content in Gliricidia sepium could stimulate microbe growth, so the ammonia yield was higher in animals fed rations T1 and T2. According to Nitis19, Gliricidia Sepium leaves are a source of Rumen Degradable Protein (RDP) where 60.73% of the protein contained in the leaves could be degraded in the rumen into N-NH3 that could be used for bacterial growth. These bacteria serve as sources for high quality protein while also improving crude fiber digestibility. Therefore, the high yield of N-NH3 in this study was likely defined by the ration type and feed composition. Rations with higher protein and low energy would result in higher N-NH3 accumulation because minimal amounts are used for protein microbe synthesis.
Ration treatment T4 (high protein-high energy) resulted in higher production of N-NH3 that is likely due to the absence of synchronization between energy use and protein degradation. Thus, using a high proportion of easily digestible carbohydrate sources to balance high nitrogen levels was not effective. Corn meal as an energy source is more soluble than Gliricidia sepium leaves, the protein source in forage. When carbohydrate degradation was optimum, nitrogen degradation decreased such that the use of N-NH3 for protein microbe synthesis was not optimized which resulted in an accumulation of N-NH3 in the rumen. Devant et al.20 reported that such decreases in protein degradation were not solely due to changes in pH but instead correlated with the type of fermented substrate or predominant microbe population that arose in response to particular feed ratios.
In general this study showed that variations in the N-NH3 concentration among the four treatments largely arose from different types of feed and the degradation rate. Decreases in carbohydrate-nitrogen degradation synchronization to produce N-NH3 was a contributing factor to the low degradibility of crude protein that resulted from lowered microbe activity in the rumen. Although, Satter and Slyter21 showed that 50 mg L–1, N-NH3 was adequate to support maximum growth of rumen bacteria, appropriate levels of N-NH3 for rumen bacteria levels could be as low as 20 mg L–1 while, higher levels led to an excessive margin. The N-NH3 yield was essentially optimal due to feed nitrogen degradation but also could have accumulated in the collecting syringe and was not absorbed during measurement due to the effect of buffer solutions containing N-NH3.
Volatile Fatty Acid (VFA) yield: The molar amount of each VFA in the rumen can be determined by the amount of forage in the ration, dry matter intake and feeding method22. Total VFA of rations T2, T3 and T4 was higher (p<0.01) than that for T1 and the VFA yield increased concurrently with increased amounts of easily digestible carbohydrate. The VFA concentration in the rumen reflects feed fermentability and is the main energy source for ruminants and along with ammonia is a major component of microbial protein. As such, the VFA concentration is directly related to the degree to which the feed is fermentable23.
The highest acetic acid level produced (mM) was observed in ration T3 which had a higher forage proportion and was followed by T2, T4 and T1, respectively (p<0.01). The VFA level also correlated with a higher proportion of crude fiber, Acid Detergent Fiber (ADF) and Neutral Detergent Fiber (NDF) in ration T3 relative to the other rations (Table 2). Although, McDonald et al.15 found that a high fiber content in forage resulted in higher production of acetic acid, the acetic acid values for rations T1, T2 and T4 in this study did not reflect this trend treatment. Ration treatment T1 had higher crude fiber, ADF and NDF than T2 and T4 but still had a higher proportion of acetic acid (Table 2, 3). The fatty acid proportion in ruminant digestion is determined by the type and composition of structural and non-structural carbohydrates as well as the amount of forage in the feed24. A high forage composition results in high acetic acid levels while, propionic acid levels increase when the concentration of degradable carbohydrate in the ration exceeds that of crude fiber25.
The propionic acid yield (mM) of T2, T3 and T4 was higher than that of T1 (p<0.01) indicating that the increased amount of protein and energy in both T2 (high energy-standard protein) and T4 (high energy-high protein) positively affected propionic acid yield. Pilajun and Wanapat26 reported that in vitro digestibility and VFA level, especially that of propionic acid increased along with substrate concentration. These increases are correlated with an adequate amount of degradable carbohydrate in the ration. A sufficient propionic yield is a beneficial component of cattle fattening rations because this acid increases the rate of body tissue synthesis.
| Table 2: | In vitro chemical composition of fattening rations with different protein-energy levels fed to Bali cattle |
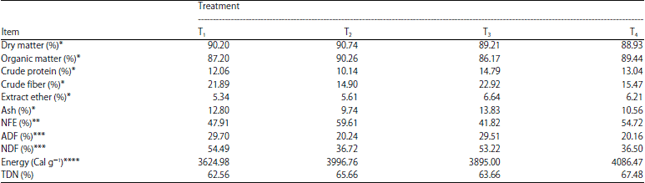 | |
*Results analysis of Animal Feed Laboratory Technology, Faculty of Animal Science, Gadjah Mada University (2015), **NFE: [100–(Ash%+CF%+EE%+CP%)], ***Analysis results from the Forage Laboratory, Faculty of Animal Science, Gadjah Mada University (2015) and ****Analysis results from the Laboratory Chemistry and Biochemistry, Study Center Food And Nutrition, Gadjah Mada University (2015) | |
| Table 3: | In vitro average pH, DMD, OMD, N-NH3 yield, VFA yield and microbial protein synthesis (Mean±SD) of fattening rations with different protein-energy levels fed to Bali cattle |
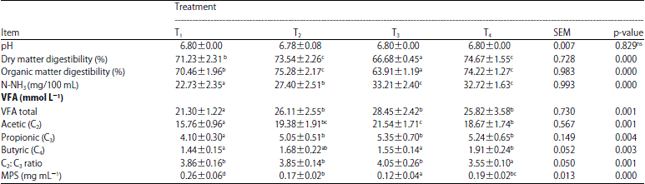 | |
| *Different superscripts in the same column indicates the effect of significantly different at p<0.05, Ns: Not-significant, MPS: Microbial protein synthesis and SEM: Standar error of mean | |
Accordingly, the propionic yield could be improved by including more concentrated and degradable carbohydrate in feed rations27. Nevertheless, T1 and T3 showed inconsistent results in terms of propionic acid wherein T1 (standard protein-standard energy) with 19% degradable carbohydrate had a lower propionic yield than did T3 (high protein-standard energy) which included only 2% degradable carbohydrates.
The butyric acid yield increased linearly with the concentration of degradable carbohydrate in rations with different protein and energy levels (p<0.01). Treatment T4 was significantly higher (p<0.05) than T1 and T3 but essentially equal to that of T2 while, T1 was equal to that of T2 and T3.
The acetic:propionic acid ratio (C2:C3 ratio) of T4 was lower (p<0.01) than that of T1, T2 and T3. An increase in the C2:C3 ratio can increase the rate of body tissue synthesis which in turn enhances cattle growth performance. Moreover, the acetic (C2): propionic (C3) acid synthesis ratio is generally used as an efficiency standard of energy allocation in ruminants wherein a high ratio translates to low energy efficiency, particularly in fattening rations28.
Perry et al.29 showed that increased starch concentrations in rations could diminish the acetic:propionic acid ratio meanwhile, structural carbohydrates such as hemicellulose produce higher acetic:propionic acid ratios. Thus, lower C2:C3 ratios are correlated with more efficient energy use because very little energy is devoted to CH4 (methane) production30.
Protein microbe synthesis: Protein microbe synthesis (mg mL–1) by cattle fed T1 was higher (p<0.01) than for cattle fed T4, T2 and T3, respectively (Table 3). The relatively high amount of microbial protein synthesis was closely related to the low proportion of N-NH3 that is caused by adequate and synchronized N-NH3 and energy supply for protein microbe synthesis which was likely responsible for the higher values seen for the T2 and T4 rations. Ration T1 had a better balance between protein and carbohydrate degradation that resulted in an optimum efficiency of microbial protein synthesis. Carbohydrates are the main energy source for bacteria although, carbohydrates also are viable carbon frames for protein synthesis combined with ammonia16. Protein synthesis by rumen bacteria depends on the amount and type of carbohydrates that are used as energy sources for peptide bond synthesis. Highly fermentable carbohydrates such as starch or sugar are more effective for stimulating and inducing microbe growth than alternate sources (e.g., cellulose)31.
Ration treatment T2 (high energy-standard protein) consisted of 35% forage and 65% carbohydrate and was easily digestible while, T4 (high protein-high energy) had 57% forage and 43% digestible carbohydrate. Both showed significant protein microbe synthesis was observed from the high N-NH3 yields. This result indicated that the N-NH3 yield was not optimized, even though these rations had adequate amounts of available energy in the form of digestible carbohydrates. The suboptimal N-NH3 yield could be due to a lack of synchronization between protein and carbohydrate degradation rates that restricted microbial protein synthesis in rations T2 and T4. When the level of digestible protein was higher than that of carbohydrate fermentation occurs and most nitrogen is dissipated as ammonia. Conversely, when the carbohydrate fermentation rate was higher than that of protein degradation and microbial protein synthesis could decrease32. Although, the effect of an unsynchronized supply of carbohydrate and nitrogen for rumen microorganisms is unclear, the combined ecosystem of rumen microorganisms is sufficiently complex such that the nutrition supply could be synchronized to certain subpopulations but not to others33. Rumen fermentation products, VFAs expressed as molar concentrations of C2 (acetic), C3 (propionic) and C4 (butyric) acids, multiple chain fatty acids and N-NH3 significantly determined the energy efficiency and the production rate of microbial protein synthesis in the rumen. As such, consideration of rumen fermentation patterns and how they can be manipulated through changes in feed ration composition is essential to optimize cattle productivity34.
The less than optimal production of protein by rumen microbes in cattle fed T2 and T4 correlated with the high amount of digestible carbohydrate in these rations. Several previous studies reported that the efficiency of protein microbe synthesis tended to increase if the fermentable carbohydrate supply represented less than 30% of the total ration35 but decreased when the supply was above 70%. This decreased efficiency of microbial protein synthesis in the small intestines of cattle occurred when the feed contained more than 70% concentrate which could accelerate the degradation rate of non-structural carbohydrates and lead to unsynchronized fermentation36. Indeed, Devant et al.20 reported that rations with high carbohydrate levels and low protein concentrations had adequate nitrogen to support microbe growth but restricted microbe protein synthesis and nutrition digestibility.
Ration treatment T3 (high protein and standard energy) composed of 98% forage and 2% carbohydrate was easily digestible and had a higher N-NH3 yield that indicated the lowest level of microbial protein synthesis relative to the other rations. This difference was due to the use of a carbohydrate source with lower digestibility as an energy source to convert N-NH3 into microbial protein. This lowered digestibility resulted in an imbalance between protein and carbohydrate degradation and in turn the high yield of N-NH3 was not optimized for microbial protein synthesis because of an imbalanced supply of carbon frames for use as an energy source.
Generally, the effects produced by T3 were consistent with several previous findings showing that feed with high protein levels must also have a high availability of energy sources such that N-NH3 derived from degradation of feed protein could be used to synthesize microbial protein. According to Huber and Herrera-Saldana37, such synchronized energy-protein release in the rumen is one contributing factor that affects the efficiency of microbial protein synthesis. This synchronization makes energy and protein simultaneously available during cattle growth3.
CONCLUSION
Conclusively, rations with different protein-energy levels fed to male Bali cattle being raised on smallholder farms did not severely affect rumen pH which remained within normal ranges. Dry matter and organic matter digestibility of these rations could be improved but N-NH3 and VFA yields were not optimal to support microbial protein synthesis.
REFERENCES
- Madsen, J. and T. Hvelplund, 1994. Prediction of in situ protein degradability in the rumen. Results of a European ringtest. Livestock Prod. Sci., 39: 201-212.
CrossRefDirect Link - Owens, F.N. and W.G. Bergen, 1983. Nitrogen metabolism of ruminant animals: Historical perspective, current understanding and future implications. J. Anim. Sci., 57: 498-518.
Direct Link - Mullik, M.L., 2006. [Supplementation strategies to improve efficiency of rumen microbial protein synthesis on cattles fed with tropical grass hay]. Jurnal Ilmu Ternak dan Veteriner, 11: 15-23, (In Indonesian).
Direct Link - Klusmeyer, T.H., R.D. McCarthy Jr., J.H. Clark and D.R. Nelson, 1990. Effects of source and amount of protein on ruminal fermentation and passage of nutrients to the small intestine of lactating cows. J. Dairy Sci., 73: 3526-3537.
CrossRefDirect Link - Tahuk, P.K., A.A. Dethan and Stefanus Sio, 2013. Kinerja sapi bali jantan pada penggemukan menggunakan pakan tunggal hijauan serta kombinasi hijauan dan konsentrat dengan level protein pakan berbeda di pulau timor. Laporan Penelitian Hibah Bersaing Perguruan Tinggi, Direktorat Jenderal Pendidikan Tinggi.
- Buxton, D.R. and M.R. Brasche, 1991. Digestibility of structural carbohydrates in cool-season grass and legume forages. Crop Sci., 31: 1338-1345.
CrossRefDirect Link - Soeparno and H.L. Davies, 1987. Studies on the growth and carcass composition in Daldale wether lambs. 2. The effect ofdietary protein/energy ratio. Crop Pasture Sci., 38: 417-426.
CrossRefDirect Link - Tilley, J.M.A. and R.A. Terry, 1963. A two-stage technique for the in vitro digestion of forage crops. Grass Forage Sci., 18: 104-111.
CrossRefDirect Link - Filipek, J. and R. Dvorak, 2009. Determination of the volatile fatty acid content in the rumen liquid: Comparison of gas chromatography and capillary isotachophoresis. Acta Veterinaria Brno, 78: 627-633.
CrossRefDirect Link - Chaney, A.L. and E.P. Marbach, 1962. Modified reagents for determination of urea and ammonia. Clin. Chem., 8: 130-132.
PubMedDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Erdman, R.A., 1988. Dietary buffering requirements of the lactating dairy cow: A review. J. Dairy Sci., 71: 3246-3266.
CrossRefDirect Link - Bach, A., S. Calsamiglia and M.D. Stern, 2005. Nitrogen metabolism in the rumen. J. Dairy Sci., 88: E9-E21.
CrossRefDirect Link - Harris, L.E., L.C. Kearl and P.V. Fonnesbeck, 1972. Use of regression equations in predicting availability of energy and protein. J. Anim. Sci., 35: 658-680.
PubMedDirect Link - Devant, M., A. Ferret, S. Calsamiglia, R. Casals and J. Gasa, 2001. Effect of nitrogen source in high-concentrate, low-protein beef cattle diets on microbial fermentation studied in vivo and in vitro. J. Anim. Sci., 79: 1944-1953.
CrossRefDirect Link - Satter, L.D. and L.L. Slyter, 1974. Effect of ammonia concentration on rumen microbial protein production in vitro. Br. J. Nutr., 32: 199-208.
CrossRefPubMedDirect Link - Min, B.R., T.N. Barry, G.T. Attwood and W.C. McNabb, 2003. The effect of condensed tannins on the nutrition and health of ruminants fed fresh temperate forages: A review. Anim. Feed Sci. Technol., 106: 3-19.
CrossRefDirect Link - Maynard, L.A., J.K. Loosli, H.S. Hintz and R.G. Warner, 1979. Animal Nutrition. 7th Edn., Tata McGraw-Hill Publishing Company, New Delhi, India, Pages: 602.
Direct Link - Pilajun, R. and M. Wanapat, 2014. Effect of roughage to concentrate ratio and plant oil supplementation on in vitro fermentation end-products. Pak. J. Nutr., 13: 492-499.
CrossRefDirect Link - Cherney, D.J.R., J.H. Cherney and L.E. Chase, 2003. Influence of dietary nonfiber carbohydrate concentration and supplementation of sucrose on lactation performance of cows fed fescue silage. J. Dairy Sci., 86: 3983-3991.
CrossRefPubMedDirect Link - Suwandyastuti, S.N.O., 2007. [The Rumen metabolism product in male sheep]. Anim. Prod., 9: 9-13, (In Indonesian).
Direct Link - Perry, T.W., A.E. Cullison and R.S. Lowrey, 2003. Feed and Feeding. 6th Edn., Prentice Hall, New Jersey, USA., ISBN-13: 978-0130970473, Pages: 675.
Direct Link - Suwandyastuti, S.N.O., 2013. [Rumen metabolism product on growing cattle ongole grade]. Agripet, 13: 31-35, (In Indonesian).
Direct Link - Stern, M.D. and W.H. Hoover, 1979. Methods for determining and factors affecting rumen microbial protein synthesis: A review. J. Dairy Sci., 49: 1590-1603.
Direct Link - Nocek, J.E. and J.B. Russell, 1988. Protein and energy as an integrated system. Relationship of ruminal protein and carbohydrate availability to microbial synthesis and milk production. J. Dairy Sci., 71: 2070-2107.
CrossRefDirect Link - Suwandyastuti, S.N.O. and E.A. Rimbawanto, 2015. [Rumen metabolism product on lactating dairy cattle]. Agripet, 15: 1-6, (In Indonesian).
Direct Link - Huber, J.T. and L. Kung Jr., 1981. Protein and nonprotein nitrogen utilization in dairy cattle. J. Dairy Sci., 64: 1170-1195.
CrossRefDirect Link - Polan, C.E., 1988. Update: Dietary protein and microbial protein contribution. J. Nutr., 118: 242-248.
PubMedDirect Link