
Sri Mawati
Department of Animal Science and Agriculture, University of Diponegoro, Semarang, Central Java, Indonesia
Agung Purnomo Adi
Department of Animal Science and Agriculture, University of Diponegoro, Semarang, Central Java, Indonesia
Soedarsono
Department of Animal Science and Agriculture, University of Diponegoro, Semarang, Central Java, Indonesia
Sunarso
Department of Animal Science and Agriculture, University of Diponegoro, Semarang, Central Java, Indonesia
Pakistan Journal of Nutrition
Year: 2016 | Volume: 15 | Issue: 12 | Page No.: 1055-1060







ABSTRACT
Background: The development of sheep farming has a strategic role in efforts to fulfill protein requirements from animal-originated food. However, the farming of ruminants has environmental impacts on water, soil and air. The pollution from these impacts can be controlled externally and internally. Internal control is achieved through improvement in livestock production efficiency, which is possible with the provision of rations that supply protein of the quality and in the amounts appropriate to meet animal requirements and farming objectives. Objective: This study was conducted to increase productivity of sheep and control pollution impact by manipulating the energy-protein ratio of feed. Methodology: In an experiment, the six complete feed formula were fed to sheep with the following protein-energy ratios: 0.158, 0.166, 0.195, 0.198, 0.230 and 0.272. Feed was provided at 4% b.wt., twice a day, i.e., morning and afternoon. In rumen fluid, the following parameters of rumen fermentation were examined: pH, volatile fatty acids and NH3. Results: In all treatments, the pH of rumen fluid was higher before eating (0 h) than at 3 and 6 h post-feeding. Treatment with different protein-energy ratios of feed significantly affected the pH and VFA and NH3 content of the rumen fluid. Conclusion: The adjustment of the protein-energy ratio of feed can be used as a strategy to control pollution as the production of farming sheep increases. The decline in nitrogen pollution to the environment will reduce negative impacts on the environment damage, thereby improving the health of livestock farmers.
PDF Abstract XML References Citation
Received: July 12, 2016;
Accepted: September 07, 2016;
Published: November 15, 2016
How to cite this article
Sri Mawati, Agung Purnomo Adi, Soedarsono and Sunarso, 2016. Effect of Feed with Different Energy-protein Ratios on Parameters
of Sheep Ruminal Fermentation. Pakistan Journal of Nutrition, 15: 1055-1060.
DOI: 10.3923/pjn.2016.1055.1060
URL: https://scialert.net/abstract/?doi=pjn.2016.1055.1060
DOI: 10.3923/pjn.2016.1055.1060
URL: https://scialert.net/abstract/?doi=pjn.2016.1055.1060
INTRODUCTION
Livestock productivity must increase to meet the requirements for food that originates from animals, which can be achieved primarily with improvements in feed inputs (quantity and quality). Animal production (in this study, ruminants) is essentially a biochemical process with feed as a substrate that undergoes fermentation, digestion and metabolism into livestock products and residual metabolites, which are excreted in feces and urine. To improve livestock productivity, nutrients can be manipulated with adjustments to the feed protein-energy ratio1. As a consequence of the increased productivity of livestock, the production of excreta increases, as confirmed by Jarvis et al.2, who states that the intensification of livestock production systems is a primary cause of the increase in the release of ammonia (NH3) to the atmosphere (an increase of approximately 50% since 1950).
Pollution is controlled in two ways, externally and internally. External control includes, among other possibilities, the following: (1) Increased sanitation of farm micro-environments with waste management installations (e.g., biogas production) and (2) Use of ammonia in biofermentation for productive purposes (e.g., fertilizers and single-cell protein as high quality feed). Internal control is achieved by improving the efficiency of livestock production. A primary approach for the internal control of pollution is the selection of feed ingredients and preparation of appropriate rations to minimize N excreta but maintain high livestock productivity. Jarvis et al.2 described manipulating rations for the efficient use of N, which can be achieved by providing rations that supply protein of the appropriate quality and amount to meet the animal’s requirements and farming objectives. Protein feed entering the rumen consists of pure protein and nonprotein nitrogen (NPN) that are degraded into peptides and amino acids that ultimately produce ammonia3. Ammonia (NH3) is formed and combined with a carbon skeleton (alpha keto acid) during carbohydrate fermentation and is used to synthesize the microbial protein that is the primary protein source for ruminants4. The supply of energy precursors and Nitrogen (N) is often a primary limiting factor in the synthesis of microbial protein, but the synthesis also depends on the kinetic availability of nutrients throughout the day based on the intensity of microbial activity in the rumen5. Microbial protein synthesis occurs efficiently when ammonia is available, followed by the availability of energy and carbon. When the availability of ammonia exceeds the fermentation of carbohydrates, the ammonia is not used efficiently in the formation of microbial protein. According to Widyobroto6, the ideal condition to synthesize microbial protein is when the fermented carbohydrate source is available at the same time as the protein source, thus, a requirement for formulating concentrates for ruminants is the consideration of the ratio of protein to energy content. Parameters of rumen fermentation (i.e., pH, NH3 and VFA) are used as indicators of the availability of precursors for the growth and development of rumen microbes.
MATERIALS AND METHODS
Twenty four male, thin-tail sheep, aged 6-7 months, with an average initial weight of 13±1.56 kg (CV = 11.78%) were used in this study. The feed used in this study was a complete feed composed of elephant grass (Pennisetum purpureum), soy flour, rice bran, dried cassava and molasses. The complete feeds were formulated with a protein content of 10, 12 or 14% and a TDN of 60 or 65%.
The sheep were allocated in a completely randomized design with 6 treatments and 4 replications. The six treatments of complete feed were as follows: T1 = 10% protein and 60% TDN, T2 = 10% protein and 65% TDN, T3 = 12% protein and 60% TDN and T4 = 12% protein and 65% TDN, T5 = 14% protein and 60% TDN and T6 = 14% protein and 65% TDN. Based on the test results, the Crude Protein (CP) content of the six treatments was 9.91, 9.99, 11.86, 12.01, 14.08 and 14.10 and the TDN values were 62.70, 60.33, 60.86, 60.60, 61.33 and 51.89%, respectively. Accordingly, in this study, these actual values were used to determine the ratios that were the basis for discussion of results. Therefore, the actual values of the CP-TDN ratios were 0.158, 0.166, 0.195, 0.198, 0.230 and 0.272, respectively, for the six treatments. The CP formulations and test results are shown in Table 1.
Elevated, individual cages made of wood 150 cm in length, 75 cm in width and 120 cm in height, equipped with individual feed and water bunks were used in the experiment. The equipment used included a set of tools for maintenance and the metabolic cages and associated equipment.
The maintenance tools included tools for sanitation and different scales. To weigh animals, the scale had a capacity of 300 kg and precision of 0.01 kg. To weigh feed, the scale had a capacity of 6 kg and precision of 2 g. Feed was provided to the animals at 4% of their body weights twice daily, i.e., 07.00 and 16.00. Drinking water was provided ad lib. The residual feed was weighed before the morning feeding. Sheep were weighed once each week and the feed ration was adjusted accordingly.
| Table 1: | Complete feed formula and nutrient content for treatments with different CP-TDN ratios |
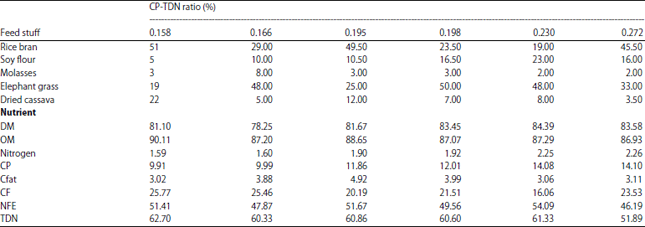 | |
| DM: Dry matter, OM: Organic matter, Cfat: Crude fat, CF: Crude fiber, CP: Crude protein, NFE: Nitrogen-free extract | |
Rumen fluid was collected at 0 h before feeding and at 3 and 6 h postfeeding. A sheep was held immobile, a hose connected to a vacuum pump was then inserted into the mouth until reaching the rumen and the vacuum pump was operated until the rumen fluid sample was collected. The pH was measured and then the samples were transferred to plastic bottles, which were centrifuged at 1500 rpm for 15 min to obtain supernatants. The samples were sealed and placed in freezer before transport to the laboratory for the analysis of the NH3 and VFA content.
The data were analyzed using analysis of variance and when the treatment effect was significant, an LSD test with a 5% error level was conducted to determine the effect among treatments7. The variables measured in the rumen fluid in this study were pH, VFAs (i.e., acetic, propionic and butyric acids) and NH3.
RESULTS AND DISCUSSION
The data on the parameters of sheep rumen fermentation for the treatments with different ratios of protein and TDN are shown in detail in Table 2. In all treatments, the pH of rumen fluid before eating (0 h) was higher than that at 3 and 6 h postfeeding.
The lowest pH occurred at 3 h postfeeding, the pH then increased at 6 h postfeeding, indicating that the fermentation intensity was highest between 3 and 6 h postfeeding. The pH of rumen fluid after the food ration is provided tends to decrease gradually because of fermentation in the rumen, particularly that of carbohydrates, which are degraded into VFA products. Ruminal pH in combination with microbial populations, the nature of the substrate and environmental factors such as temperature and concentrations of cations and soluble carbohydrates may govern bacterial attachment8. Ruminal pH is one of the most important of these factors because fibrolytic bacteria are very sensitive to and dependent on changes in the pH9.
As a product of rumen fermentation, VFAs are acidic compounds that decrease rumen pH. Consistent with this relation, Kerley et al.10 observed a decrease in the pH of rumen fluid with the increased production of VFAs.
However, in all dietary treatments in this study, the range of pH values was 7.16-7.72. Below this range, the rumen function may be negatively affected11. The pH values for sheep in this study were higher than those of Abubakr et al.12 for goats fed diets containing palm oil byproduct with a pH range from 6.07-6.42 and those of Sharifi et al.13 with a range from 6.26-6.61. The high pH values of this study were presumably due to the increased degradation of peptides/proteolysis and deamination of amino acids in the rumen.
Rumen fluid is conducive to the process of fermentation. According to Van Soest14, the optimum pH for cellulolytic microbes is above 6.5 and when the pH is below 6.5, the rate of cell wall degradation decreases. Different types of feed stuffs in a ration may lead to differences in the palatability and nutrient content, which results in differences in the amount of feed consumed by animals15. The range of the pH in rumen fluid is 5.5-7.216,9, with the optimum activity of cellulolytic bacteria in the rumen at a pH of 6.7±0.514.
The pH of rumen fluid increased again 6 h after eating, which is consistent with the study of Baldwin and Denham17, who found that the highest fermentation intensity in the rumen occurs up to 5 h after eating. The increase in pH was likely due to the absorption of the fermentation products NH3 and VFA in the rumen.
| Table 2: | Parameters of sheep ruminal fermentation for treatments with different feed protein-TDN ratios |
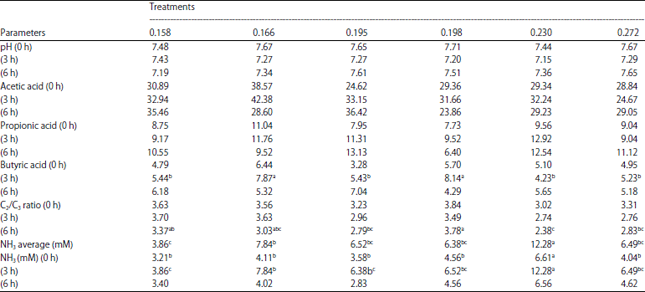 | |
| abcDifferent superscripts in the identical row indicate significant differences (p<0.05) | |
Based on the results of Javaid et al.18, who studied buffaloes (Bubalis bubalis) fed diets containing ascending levels of Ruminally Degradable Protein (RDP), a decrease in the pH is associated with an increase in the dietary RDP at 3 h postfeeding. The results of Baumann et al.19 are similar with decreases in ruminal pH with the addition of RDP to corn grain-based diets in steers.
VFA: In this experiment, the production of acetic acid, butyric acid and propionic acid increased in rumen fluid postfeeding. The acetic acid production in the treatments with different CP-TDN ratios ranged from 24.62-42.38 mM. This range in acetic acid production was caused by the type and composition of feed stuffs in a ration, which is consistent with the argument of Lestari et al.20 in that differences are likely due to differences in the type and amount or percentage of feed stuffs used.
In the rumen fluid of treatments with CP-TDN ratios of 0.166, 0.198 and 0.230, the production of acetic acid increased at 3 h postfeeding and then decreased at 6 h postfeeding. In general, the total VFA concentrations increased postfeeding because of the fermentation of the feed carbohydrates, whereas, the possible absorption of the fermentation products explains the decrease in pH at 6 h postfeeding. This decrease was consistent with the scenario described by McDonald et al.4, who found that after fermentation, VFAs are absorbed through the rumen walls of the reticulum and omasum, with some passing into the abomasum and of the small intestine as an energy source for the animals.
The results showed that the production of propionic acid increased at 3 h postfeeding, indicating that the fermentation of feed carbohydrates had occurred. According to Sulistyowati et al.21, the performance of rumen fermentation is influenced primarily by the feed consumed by the animals. In the treatments with CP-TDN ratios of 0.158, 0.195 and 0.272, the production of propionic acid continued to increase at 6 h postfeeding. This increase possibly occurred because rice bran was used a source of carbohydrates in the rations of treatments with CP-TDN ratios of 0.158, 0.195 and 0.272 with this use relatively higher than that in the other three treatments. According to Owen and Goetsch16, cellulolytic microbes actively produce more propionic acid when the ration contains soluble carbohydrates.
Butyric acid production, in general, increased postfeeding because of the fermentation of feed carbohydrates. The production of butyric acid declined at 6 h postfeeding, identical with the production of acetic acid, likely because the kinetics of VFAs decline after reaching the optimal concentrations. The decrease occurs because VFAs are absorbed through the walls of the rumen, reticulum and omasum; some VFAs can also pass into the abomasum and are absorbed in the small intestine4.
Based on the results, the ratios of acetic and propionic acids in sheep rumen fluid varied among the treatments. The treatment with a CP-TDN ratio of 0.230 had the lowest ratio of acetic and propionic acids compared with that of the other treatments. This result was consistent with that for ADG in the identical treatment, which was the optimum among all treatments. Syapura et al.22 reported that a low ratio of acetic and propionic acids stimulates fattening and as further discussed by Parakkasi23, increased levels of propionic acid in the rumen likely contribute to the production of glucose.
In this study, the treatment with the CP-TDN ratio of 0.230 resulted in the highest ammonia concentration in the rumen. Nitrogen-ammonia (N-NH3) in rumen fluid is derived from protein degradation and nonprotein nitrogen (NPN) compounds in feed and from N in urea in saliva24. The treatments with CP-TDN ratios of 0.230 and 0.272 were the rations with a level of crude protein of 14%. Therefore, the higher protein content resulted in a greater production of N-NH3. Increases in the rumen N-NH3 are associated with the consumption of CP. According to Arora25, the NH3 in the rumen is from feed CP and saliva urea degraded in the rumen, with increases in CP consumption expected to increase ammonia concentrations. As described by Colmenero and Broderick26, increases in rumen ammonia levels are consistent with increases in level of feed CP, primarily that of highly degradable protein. The level of NH3 in the treatment with a CP-TDN ratio of 0.272 was lower than that of the treatment with a CP-TDN ratio of 0.230, which was likely due to the higher CF content in the treatment with a CP-TDN ratio of 0.272, although the energy content was higher (65%). This result is consistent with that described by Hungate27, who found that the concentration of ammonia varies and is affected by protein degradation and the degradation of other organic materials. As Ranjhan28 explained, ammonia is a primary component of soluble nitrogen in rumen fluid that is required by rumen microbes to synthesize body proteins, which is dependent on the availability of a carbon skeleton (VFA).
The increase in the N-NH3 concentration in the rumen fluid at 3 h postfeeding was an indication of the degradation of feed N. The concentration of N-NH3 varies, among other factors, depending on the rate of protein degradation and the amount of feed protein27. Protein in the rumen is hydrolyzed with proteolytic enzymes produced by rumen microbes and at adequate concentrations, N-NH3 is used to synthesize microbial protein29. Rumen N-NH3 concentrations are also affected by the time after feeding25, with an increase in rumen N-NH3 concentrations after feeding, reaching an optimum at 3 h after consuming feed18.
Based on the results of this study, an increase in dietary protein was not always followed by an increase in N-NH3, however, the N-NH3 concentration increased at the dietary protein equilibrium of 14% and TDN of 60% (0.230) at 0 h and at 3 h postfeeding. Additionally, an increase in dietary source of protein without the proper addition of an energy source suppressed butyric acid production, particularly at 3 h post-feeding, although the concentrations of propionic and acetic acids were not affected. The results of this study differed from those of Javaid et al.18, who reported that an increase in dietary percentages of RDP increased the concentration of N-NH3 at either 3 or 6 h post-feeding.
The concentration of N-NH3 in the rumen fluid decreased at 6 h after eating. A decrease in the concentration of ammonia in rumen fluid depends on the rate of nitrogen use by rumen microbial biomass and ammonia absorption30. Microbes use some of the ammonia resulting from rumen microbial degradation to form body proteins and some is delivered to the liver through the portal vein and converted to urea31.
CONCLUSION
In this study, the treatment with a CP-TDN ratio of 0.230 resulted in the optimum parameters for rumen fermentation in sheep.
REFERENCES
- Mawati, S., S. Soedarsono, S. Sunarso and A. Purnomoadi, 2013. The effects of different energy and ratio to sheep's nutrient intake and digestibility. Int. J. Sci. Eng., 4: 76-79.
CrossRefDirect Link - Jarvis, S., N. Hutchings, F. Brentrup, J.E. Olesen and K.W. van de Hoek, 2011. Nitrogen Flows in Farming Systems Across Europe. In: The European Nitrogen Assessment, Sutton, M.A., C.M. Howard, J.W. Erisman G. Billen and A. Bleeker et al. (Eds)., Cambridge University Press, Cambridge, pp: 211-228.
Direct Link - Miron, J., D. Ben-Ghedalia and M. Morrison, 2001. Invited review: Adhesion mechanisms of rumen cellulolytic bacteria. J. Dairy Sci., 84: 1294-1309.
CrossRefDirect Link - Sung, H.G., Y. Kobayashi, J. Chang, A. Ha, I.H. Hwang and J.K. Ha, 2007. Low ruminal pH reduces dietary fiber digestion via reduced microbial attachment. Asian-Australasian J. Anim. Sci., 20: 200-207.
CrossRefDirect Link - Kerley, M.S., G.C. Fahey Jr., L.L. Berger, N.R. Merchen and J.M. Gould, 1987. Effects of treating wheat straw with pH-regulated solutions of alkaline hydrogen peroxide on nutrient digestion by sheep. J. Dairy Sci., 70: 2078-2084.
CrossRefDirect Link - De Veth, M.J. and E.S. Kolver, 2001. Diurnal variation in pH reduces digestion and synthesis of microbial protein when pasture is fermented in continuous culture. J. Dairy Sci., 84: 2066-2072.
CrossRefPubMedDirect Link - Abubakr, A.R., A.R. Alimon, H. Yaakub, N. Abdullah and M. Ivan, 2013. Digestibility, rumen protozoa and ruminal fermentation in goats receiving dietary palm oil by-products. J. Saudi Soc. Agric. Sci., 12: 147-154.
CrossRefDirect Link - Sharifi, M., M. Bashtani, A.A. Naserian and H. Khorasani, 2013. Effect of dietary crude protein level on the performance and apparent digestibility of Iranian Saanen kids. Afr. J. Biotechnol., 12: 4202-4205.
Direct Link - van Soest, P.J., 1994. Nutritional Ecology of the Ruminant. 2nd Edn., Cornell University Press, Ithaca, New York, Pages: 476.
Direct Link - Nusi, M., R. Utomo and Suparno, 2011. [Effect of utilization of corn cobs in complete feed and undegraded protein suplementation on gain and meat quality of ongole crossbred cattle]. Bull. Peternakan, 35: 1-9.
Direct Link - Baldwin, R.L. and S.C. Denham, 1979. Quantitative and dynamic aspects of nitrogen metabolism in the rumen: A modeling analysis. J. Anim. Sci., 49: 1631-1637.
PubMedDirect Link - Javaid, A., Mahr-un-Nisa, M. Sarwar and M.A. Shahzad, 2008. Ruminal characteristics, blood pH, blood urea nitrogen and nitrogen balance in Nili-ravi buffalo (Bubalus bubalis) bulls fed diets containing various levels of ruminally degradable protein. Asian-Aust. J. Anim. Sci., 21: 51-58.
Direct Link - Baumann, T.A., G.P. Lardy, J.S. Caton and V.L. Anderson, 2004. Effect of energy source and ruminally degradable protein addition on performance of lactating beef cows and digestion characteristics of steers. J. Anim. Sci., 82: 2667-2678.
PubMedDirect Link - Lestari, D.A., L. Abdullah and Despal, 2015. Comparative study of milk production and feed efficiency based on farmers best practices and National Research Council. Med. Peternakan: J. Anim. Sci. Technol., 38: 110-117.
CrossRefDirect Link - Sulistyowati, E., A. Sudarman, K.G. Wiryawan and T. Toharmat, 2014. In vitro goat fermentation of PUFA-diet supplemented with yeast and curcuma xanthorrhiza Roxb. Media Peternakan: J. Anim. Sci. Technol., 37: 175-181.
CrossRefDirect Link - Egan, A.R. and M.J. Ulyatt, 1980. Quantitative digestion of fresh herbage by sheep. J. Agric. Sci., 94: 47-56.
CrossRefDirect Link - Colmenero, J.J.O. and G.A. Broderick, 2006. Effect of dietary crude protein concentration on milk production and nitrogen utilization in lactating dairy cows. J. Dairy Sci., 89: 1704-1712.
CrossRefDirect Link - Hungate, R.E., 1966. The Rumen and its Microbes. 1st Edn., Academic Press, London, Pages: 553.
Direct Link - Mayulu, H., M. Sunarso, M. Christiyanto and F. Ballo, 2013. Intake and digestibility of cattle's ration on complete feed based-on fermented ammonization rice straw with different protein level. Int. J. Sci. Eng., 4: 86-91.
CrossRefDirect Link