
Nabila E. Abdelmeguid
Department of Zoology, Faculty of Science, Alexandria University, Alexandria, Egypt
Hassan E. Awad
Department of Oceanography, Faculty of Science, Alexandria University, Alexandria, Egypt
Ahmed M. Ibrahim
NIOF, Alexandria, Egypt
Nabiha A. Yousef
NIOF, Alexandria, Egypt
Pakistan Journal of Nutrition
Year: 2009 | Volume: 8 | Issue: 6 | Page No.: 770-781







ABSTRACT
The aim of this study was to determine the effects of Polyaromatic Hydrocarbons (PAHs) on the fine structure of the hepatopancreas of Palaemon serratus. For this purpose, shrimps were intramuscularly injected with sublethal dose (100 ng μL-1) of perylene, christen and benzo (a) pyrene. Samples of hepatopancreas were then taken for electron microscopic examinations (three weeks postinjection). Ultrathin sections revealed that the exposure changed the structure of hepatopancreatic epithelial cells (R- and F-cells) in perylene and christen injected specimens. However, benzo (a) pyrene treated group was less affected. Major changes observed in the hepatopancreatic cells included thickening and folding of basal lamina, distorted microvillous border, condensation and margination of heterochromatin, dilation and fragmentation of RER, well-developed lysosomal compartment and swelling mitochondria. Such ultrastructural changes could serve as a useful research tool and a general biomarker in studies of toxic stress in aquatic habitat.
PDF Abstract XML References Citation
How to cite this article
Nabila E. Abdelmeguid, Hassan E. Awad, Ahmed M. Ibrahim and Nabiha A. Yousef, 2009. Ultrastructural Changes in Hepatopancreas of Palaemon serratus, Following Treatment with Petroleum Carcinogenic Compounds. Pakistan Journal of Nutrition, 8: 770-781.
DOI: 10.3923/pjn.2009.770.781
URL: https://scialert.net/abstract/?doi=pjn.2009.770.781
DOI: 10.3923/pjn.2009.770.781
URL: https://scialert.net/abstract/?doi=pjn.2009.770.781
INTRODUCTION
The application of environmental toxicology studies on non-mammalian vertebrates is rapidly expanding and, for aquatic systems crustaceans have become indicators for the evaluation of the effects of noxious compounds (Ritterhoff et al., 1996). PAHs are ubiquitous microcomponents of freshwater, marine and terrestrial environments (Neff, 1979). The principal sources of PAHs in the marine environment are industrial wastes, atmospheric fallout, petroleum activities wastes, leaching or infiltration from soil, domestic sewage, roadway runoff, natural marine oil seepage and exhausts from internal combustion engine of motorboats and ships (Hellou et al., 1994). The cycle of PAHs in the aquatic environment appears to be relatively simple. After entering water bodies from various sources, PAHs quickly become adsorbed to particulate matter and usually are deposited in the bottom sediments (Neff, 1979; Kochany and Maguire, 1994). However, biological activity in the sediment may return a small fraction from sediment PAHs to the water column.
Because most PAHs are lipophilic in nature, they tend to accumulate in living organisms (Nikolaou et al., 1984) to levels higher than those in the ambient medium where they can be transformed biochemically (Kochany and Maguire, 1994). The negative health implications of PAHs in water are not limited to aquatic life. PAHs in polluted water can circulate from microscopic aquatic organisms such as phytoplanktons to crustaceans to fish and eventually to humans through the food chain (Newsted and Giesy, 1987). Over the few past decades there has been noticeable increasing concern over the possible harmful effects of PAH-type compounds to man and other living organisms. Hundreds of aromatic hydrocarbons have been tested for carcinogenicity. Historically, the observed high incidence of stomach cancer in Iceland was attributed probably to consumption of smocked fish and meat (Also, the onset of leukemia in workers including refinery workers was linked to occupational exposures to benzene (Infante and White, 1983). There was no adequate evidence for carcinogenicity of benzo(a)pyrene in humans but sufficient evidence for carcinogenicity was found in animals (Lemaire et al., 1990). In addition, PAHs in contaminated sediments have been associated with elevated frequencies of neoplastic lesions in wild fish (Myers et al., 1987).
The Crustacean hepatopancreas is composed of blind tubules, which are internally lined with a single epithelial layer delimiting the lumen and consisting of at least four cell types: E- (embryonic), R- (resorptive), F- (fibrillar) and B- (blister-like) cells (Zilli et al., 2003). The functions carried out by each cell type seem to be established (Gibson and Barker, 1979; Dall and Moriarty, 1983; Al-Mohanna and Nott, 1986; Paquet et al., 1993). E-cells are undifferentiated, show mitotic activity and do not seem to be involved in digestive processes. All the other cell types are derived from E-cells. R-cells are the most abundant cell type and are involved in lipid, glycogen and calcium storage and in nutrient absorption. The F-cells play an important role in the synthesis and secretion of digestive enzymes into the lumen, during the 2 h following ingestion (Paquet et al., 1993) and are incapable of absorbing any nutritional element. Some controversy exists with regard to the function of B-cells. It is generally assumed that B and F cell types are mostly concerned with the synthesis and secretion of digestive enzymes (Gibson and Barker, 1979; Dall and Moriarty, 1983). However, some authors (Hopkin and Nott, 1980; Al-Mohanna and Nott, 1986; Paquet et al., 1993) have proposed that only F-cells can perform this function. In this case B-cells are thought to be the main site for nutrient absorption and digestion (but not for their storage) and for the accumulation of waste products.
In addition, the hepatopancreas fulfils a key role in the temporary storage of exoskeletal calcium, phosphate, glycogen and lipids during the different phases of the moulting cycle (Mercado-Allen, 1991; Nicol et al., 1992; Scott-Fordsmand and Depledge, 1997; Wheatly, 1997). Thus, hepatopancreas represents a corner stone in the body metabolism. Also, it plays a major role in accumulating, neutralizing and eliminating harmful chemicals in both normal and pathological conditions. It receives blood draining the gastrointestinal tract; consequently it is exposed to toxins absorbed from gut. Therefore, the hepatopancreatic tissue was chosen in the present study as a suitable and special model for the study of PAHs intoxication.
Histopathological and ultrastructural studies demonstrating the impacts of toxic organic compounds on crustacean animals were performed. Lightner et al. (1982) studied the histopathological effect of aflatoxin B1 on marine shrimp Penaeus stylirostris and P. vannamei. They found that aflatoxin produced a similar histopathological effect in both species whether the toxin was administrated with feed or by intramuscular injection. This effect is expressed as necrosis of some hepatopancreatic tubule epithelium and a marked intertubular hemocytic inflammation followed by encapsulation and fibrosis of some affected tubules. In addition, various degrees of abnormalities were induced in the hepatopancreas of the grass shrimp (Palaemonetes pugio) when exposed to dithiocarbamate biocides (Doughtie and Rao, 1983). Anderson et al. (1997) reported swollen and vacuolated R-cells in the red swamp crayfish (Procabmbarus clarkii) exposed to a petroleum-contaminated Bayou in Louisiana.
Despite of the known undesirable mutagenic and/or carcinogenic effects of PAHs on aquatic organisms, few ecotoxicological studies on the effects of these compounds on marine organisms and especially on crustacean animals have been carried out. Thus, the proposed plan of work is devoted for studying the possible morphological changes of hepatopancreas of Palaemon serratus after PAHs administration through intramuscular injection to assess whether these effects could serve as biomarkers of PAHs exposure. The hepatopancreas is examined by electron microscopy.
MATERIALS AND METHODS
Test animal selection and care: Mature shrimps namely, Palaemon serratus (Pennant, 1777) ranging from 2.7-3.6 cm carapace length and 2-4 g wet wt were currently catched from both eastern and western harbor of Alexandria. Animals were transported very carefully to the laboratory, avoiding rough handling and injuries, then they stocked in well-aerated glass aquaria (30x40x40cm). Before starting any bioassay, acclimation was conducted at least for two days in natural sea water collected from El-Montazah region far away from any oil pollution point sources. The animals were fed ad libitum daily diet of minced dry prawns.
Compounds: The PAHs used in the study were perylene, christen and benzo(a)pyrene. They were dissolved firstly in a minimum quantity of a suitable organic solvent (n-hexane).
Administration of PAHs to Palaemon serratus through intramuscular injection: Preliminary trials have been done to determine the most suitable sublethal injection dose of PAH to ensure the survival of test organisms during conveyable duration. 1 μL-1 of working solutions (in n-hexane) having concentration of about 100 ng μL-1 was found to satisfy our target. The injection was carried out intramuscularly into the lateral abdominal muscle, between the pleural membrane of the third and fourth segments using microsyringe (Hamilton type, 1 μL cap.). Injected shrimps were transferred immediately to aquaria containing clean natural sea water. Aeration was applied continuously during the assays. The test individuals were fed regularly on minced dried shrimps with taking cautions for maintaining healthy conditions in the medium.
Preparation of materials for electron microscopy: Small pieces of hepatopancreas of shrimps injected with PAHs were removed (three weeks postinjection) and fixed by immersing them immediately in 4F1G in phosphate buffer solution (pH 7.2) at 4°C for 3 hours. Specimens were then postfixed in 2% OSO4 in the same buffer at 4°C for 2 h. Samples were washed in the buffer and dehydrated at 4°C through a graded series of ethanol. For Scanning Electron Microscopy (SEM), samples of hepatopancreas were dried by means of the critical point method, mounted using carbon paste on an Al-stub and coated with gold up to a thickness of 400 Ao in a sputter-coating unit (JFC-1100 E). Observations of the coded specimens were performed in a Jeol JSM-5300 scanning electron microscope operated between 15 and 20 KeV.
For Transmission Electron Microscopy (TEM), specimens of hepatopancreas were embedded in Epon-araldite mixture in labeled beam capsules. LKB ultramicrotome was used to obtain semithin sections (1 μm thick); these were mounted on glass slide and stained with toluidine blue. Ultrathin sections (50 nm thick) were cut from selected areas for TEM. These ultrathin sections were of either pale gold or silver interference colour and were picked upon 200 mesh naked copper grids. Grids were double stained with uranyl acetate for ½ h and lead citrate for 20-30 min. (Reynolds, 1963). Scoping the grids was achieved by using Jeol 100CX TEM.
RESULTS
Ultrastructure of hepatopancreas of Palaemon serratus of control group: SEM preparations demonstrated that the hepatopancreas consisted of numerous blind ending tubules loosely bounded together by connective tissue and branching in a racemose manner [Fig. 1(A)]. Thin sections prepared for transmission electron microscopy of hepatopancreatic tubules of Palaemon serratus showed that the cells possessed distinct boundaries. The apical membranes showed long microvilli which represented the brush border seen with the light microscopic study. The microvilli were uniformly arranged and revealed an average size of 3x0.1μm [Fig. 1(B)]. Many of the microvilli possessed visible core filaments which penetrated the apical cytoplasm of the cells to form part of organelle-free zone. It was also observed in the control group that the basal lamina enveloped the hepatopancreatic cells. Its thickness averaged 0.24 μm. Myoepithelial cells attached to the basal lamina and consisted of circular and longitudinal muscle network surrounding the tubules. Our preparations of control specimens showed also two kinds of cells that were easily differentiated. The light cells (resorptive, R- or absorptive cells) predominated over the dark cells (fibrillar or F-cells). The R-cells were columnar in shape, approximately 25 μm long and 10 μm wide [Fig. 1(B,C)]. The nuclei of the R-cells appeared with different sizes. These nuclei were generally spherical with a diameter ranged from 5.1- 9.3 μm. They were centric or eccentric in position. The nuclear envelope appeared regular, porous and distinct. The distribution of chromatin showed high degree of variability. The euchromatin in most cells predominated over the heterochromatin which appeared as small irregular clumps along the periphery of the nucleus, adjacent to the inner membrane. However, in some nuclei, the heterochromatin dominated over the euchromatin and was marginally distributed.
The cytoplasm of the R-cells was deficient in the rough endoplasmic reticulum which consisted of short cisternae with few attached ribosomes. The mitochondria were observed in the apical regions. They were spherical or oval in shape with distinct membranes and cristae. Their diameter ranged from 1.5-2.1 μm. Also, they appeared with moderately lucent matrices. In addition, dark deposits within lysosome-like bodies were observed adjacent to the nuclei [Fig. 1(B)].
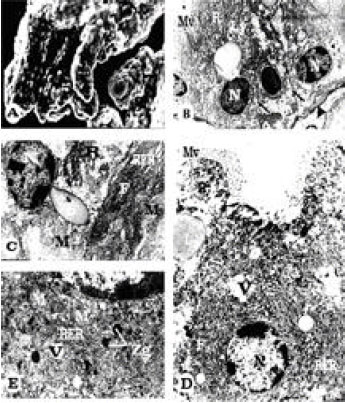 | |
| Fig. 1: | Electron micrographs of the hepatopancreas of marine shrimp, Palaemon serratus of the control group: (A) SEM showing the hepatopancreatic tubules (arrows). X350. (B) TEM demonstrating R-cells (R) with distinct cell boundaries (arrows), basally located nuclei (N), lipid droplets (L) and lysosome-like body (Ly). Arrowheads indicate basal lamina, Mv: microvilli. X2000. (C) TEM illustrating R-cell (R) with nucleus (N), relatively lucent cytoplasmic matrix and large lipid droplet (L). F-cell (F) has dark extensive RER and numerous mitochondria (M) with relatively lucent matrices. X3000. (D) TEM showing F-cell (F) containing nucleus (N) and extensive lamelliform RER. R-cell (R) having vacuolar cytoplasm and large lipid droplet (L). X3000. (E) TEM illustrating F-cell with nucleus (N), well developed RER cisternse, small vacuoles (V) and zymogen granules (Zg). X10000. |
A large apical vacuole occurred in most R-cells and contained multilayered membranes, vesicles and dark particles. Moreover, the R-cells usually contained vacuoles of irregular shapes. R-cells were also characterized by the presence of lipid droplets of different sizes adjacent to the nuclei. In addition, conspicuous quantities of glycogen granules were usually observed.
It was found that the dark (F-) cells were columnar in shape and the morphometric measurements revealed that they were approximately 26 μm long and 13.5 μm wide. Each cell had a basal nucleus which was
generally spherical in shape, and showed prominent nucleolus. The nucleus of the F-cell was generally larger (7-12 μm in diameter) than that of the R-cell and had a regular nuclear envelope with indistinct nuclear membranes [Fig. 1(D)]. In most cell nuclei, the euchromatin predominated over the heterochromatin which appeared peripherally distributed. Our preparations revealed also the presence of supranuclear vacuoles containing cellular debris.
The basal cytoplasm of the fibrillar cells were characterized by extensive rough endoplasmic reticulum which appeared slightly lamelliform at some sites with obvious electron dense ribosomes [Fig. 1(C-E)]. In addition, the mitochondria of the F-cells were observed near the cell apex and around the nucleus, but with more uniform distribution than that observed in the R-cells. They were typically spherical or ovoid in shape with a diameter ranged from 1.3-2.0 μm. In most preparations, mitochondria had moderately lucent matrices and cristae were observed [Fig. 1 (C,E)]. In few preparations, mitochondria had distinct membranes and vesicular cristae. Furthermore, zymogen granules were also found in the distal cytoplasm surrounding the nucleus of the F-cell [Fig. 1(E)].
Ultrastructural changes in hepatopancreas of Palaemon serratus treated with PAHs
Perylene-treated group: Our ultrathin sections illustrated cellular disruption and disappearance of lateral cell membranes [Fig. 2(A, B)]. The basal lamina appeared folded in some places. It was thicker (0.35 μm thick) than control (0.24 μm). Moreover, the cells were shrinked leaving the basal lamina behind. On the other hand, necrosis of the microvilli border was noted in some places and distorted appearance was observed in others. The average length of the microvilli was found to be 2 μm instead of being 3 μm in the control group. Our preparations also revealed lysis of cytoplasm in many areas. Many vacuoles containing cytoplasmic debris could be observed with highly increased frequency than normal.
Morphometric measurements of the R-cells [Fig. 2(A)] in perylene-treated animals revealed that the average length of these cells was 20 μm indicating slight decrease than normal (25 μm long). The nuclei of the R-cells varied from being spherical to oval in shape. A decrease in their diameter was observed (their average diameter ranged from 4.5-7 μm instead of being 5.1- 9.3 μm in the control group). In addition, some nuclei were elongated with eccentrically placed heterochromatin. In general, the nuclei were rich in heterochromatin content that were distributed in haphazard manner in the nucleoplasm. The R-cells exhibited a general loss of cytoplasmic density and were frequently characterized by the presence of damaged areas around the nuclei and all over the cytoplasm. This phenomenon was not observed in the normal R-cells. Scanty RER were found throughout the cytoplasm and consisted of cisternae of short lengths. Some of the R-cells were rich in accumulating dark deposits within the cytoplasm and others had numerous vacuoles containing cytoplasmic debris. Mitochondria and lipid droplets could not be easily detected in the cytoplasm.
The F-cells [Fig. 2(A,B)] were more or less columnar in shape with average length 15 μm and with 10μm width indicating a decrease in their size than in case of the control. Some of the basally located nuclei of the F-cells were normal in appearance although decreased in diameter (ranged from 5-7.5 μm) than that of the normal (ranged from 7-12 μm). Most of them contained abundant heterochromatin distributed both centrally and peripherally. Few cells showed lightly irregular nuclear envelope. Their nuclei were poor in heterochromatin content and present a paler appearance. It is of interest to observe a nuclear pocket-like structure [Fig. 2(C)] resembling that described in tumor cells of vertebrates (type 2 nuclear pocket). Although the cytoplasm of most fibrillar cells possessed an extensive RER, the cisternae had disorganized arrangement. Electron micrographs revealed even more dramatically the extremes of bizarre forms of the RER of the F-cells could assume. Moreover, our preparations revealed few cells with moderately dense background cytoplasm containing relatively inconspicuous lamellar cisternae of RER and unusual large vacuolar structures. In comparison with that observed in the control animals, neither mitochondria nor zymogen granules could be detected.
Christen-treated group: The fine structural observations of the hepatopancreatic cells of shrimps injected with christen solution revealed severe pathological features. These cells lacked cellular organization and their basal lamina increased in thickness (average thickness of which was 0.35 μm) if compared with that of the control shrimps. In addition, the apical membranes of the hepatopancreatic cells appeared with distorted, somewhat dilated microvillous border and the microvilli were short (average length of which was 1.8 μm and width was 0.1 μm). Generally, degeneration of the hepatopancreatic epithelium was noted and it showed a distinct cellular destruction.
Ultrastructurally, the R-cells [Fig. 3 (A, B)] appeared columnar in shape with rounded or oval shaped nuclei. Morphometric measurements of the nuclei of these cells revealed that their average diameter ranged from 5-8.5 μm. They had dilated nuclear envelope. The heterochromatin of these preparations predominated over the euchromatin and was centrally and peripherally placed. The cytoplasm of the R-cells contained large vacuoles with debris and had scarce RER. Moreover, densely staining deposits have been detected throughout the cytoplasm.
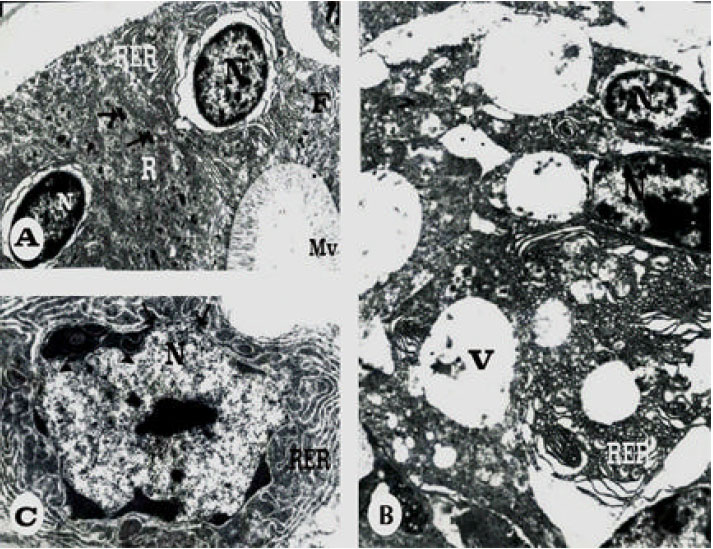 | |
| Fig. 2: | Transmission electron micrographs of the hepatopancreas of marine shrimp, Palaemon serratus intramuscularly injected with 100 ng μL-1 of perylene showing: (A) R-cell (R) with dark deposits (arrows) all over the cytoplasm, F-cell (F) with well developed RER and shrinkage of the nuclei (N) with lucent area around them. X3000. (B) Epithelial cells with nuclei (N), disorganized RER and numerous vacuoles (V) in the cytoplasm. X3000. (C) Nucleus (N) of F-cell with irregular nuclear envelope, slightly dilated nuclear pores (arrows) and nuclear pocket-like structures (arrowheads). Well developed cisternae of RER are also seen around nucleus. X7500. |
In contrast with the control group, the R-cells were characterized by reduction in lipid droplets. On the other hand, the mitochondria of the light cells were spherical or oval and their average diameter was approximately 1.8 μm. They had indistinct membranes and cristae and with electron lucent matrices. A special form of mitochondria (megamitochondria), the average diameter of which was 4 μm, could be observed in christen-treated group. These forms were pleomrphic, electron dense and surrounded with densely stained droplets.
Ultrathin sections revealed that the dark (F-) cells [Fig. 3(C)] varied from being cuboidal to triangular in shape. Their nuclei were rounded in shape and basally located (approximately 7 μm in diameter). Some of them possessed regular nuclear envelope with distinct membranes, while others showed dilated space between the nuclear membranes and surrounded by cytoplasm which exhibited lysis in many places. Most of the nuclei contained heterochromatin distributed all over the nucleoplasm. Disorganized nuclei with abnormal heterochromatin distribution could be also detected. Numerous electron dense granules, probably zymogen granules were distributed in the cytoplasm proximal to the nuclei [Fig. 3(D)]. In our electron micrographs, the F-cells had many vacuolar bodies and dense inclusions distributed all over the cytoplasm. Some cells showed scanty cisternae of RER of short lengths. Numerous polyribosomes were also seen in the cytoplasm. In contrast, other preparations showed cytoplasm loaded with RER.
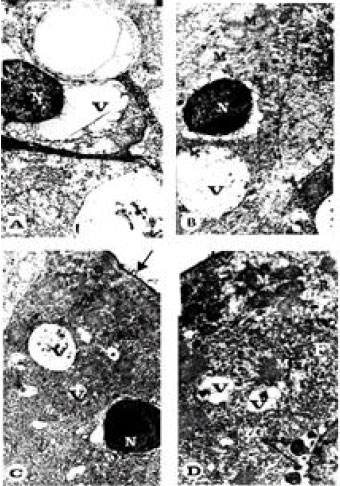 | |
| Fig. 3: | Transmission electron micrographs of the hepatopancreas of marine shrimp, Palaemon serratus intramuscularly injected with 100 ng μL-1 of christen showing: (A) R-cell with nucleus (N) and loss of cytoplasmic density due to the presence of numerous vacuoles (V). X3000. (B) R-cell (R) with disorganized nucleus (N), dense scattered granules evenly distributed in the cytoplasm, degenerative large mitochondria (M), large vacuoles (V) and lysis of cytoplasmic area surrounding the nucleus (N). X4000. (C) F-cell with nucleus (N) having abnormal heterochromatin distribution, electron dense mitochondria (M), dense inclusions and many vacuolar structures (V) in the cytoplasm. Arrowhead points at basal lamina. X3000. (D) F-cell (F) ground cytoplasm and R-cell (R), both with pyknotic mitochondria. Digestive vacuoles (V) and Zymogen Granules (Zg) are seen adjacent to the nucleus of F-cell. X5000. |
Mitochondria revealed striking variation in distribution, shape and amount even from cell to cell. Irregular shaped and pyknotic mitochondria with electron dense matrices were observed in some cells. Other mitochondria were seen degraded in the cytoplasm. In other cells, mitochondria were densely stained with vesiculated cristae.
Benzo(a)pyrene-treated group: Electron microscopic examinations showed slight cytological alterations of the hepatopancreatic tissues of benzo(a)pyrene-injected shrimps. The basal lamina in these preparations appeared normal with average thickness of 0.25 μm. Uniform striated microvillous border was observed in the hepatopancreatic cells (the average length of which was 2.7 μm and the average width was 0.1 μm) [Fig. 4(A)]. All cells possessed electron dense lateral cell membranes.
Ultrastructurally, the R-cells [Fig. 4(A)] appeared columnar in shape. Morphometric measurements revealed that their average length was 23 μm and their average width was 10.5 μm. In general, the nuclei were rounded in shape with a diameter ranged from 4.9-9.1 μm. They were basally located. Polymorphic dense area surrounding nuclear elements were observed in few nuclei of some R-cells. The heterochromatin of nuclei of the R-cells of benzo(a)pyrene-injected shrimps appeared as dense masses scattered in the nucleoplasm and as electron dense aggregates along the inner nuclear membrane. In these preparations, the presence of numerous areas of lysis all over the cytoplasm gave a lucent appearance to the cells. Besides, scanty RER were distributed in the cytoplasm. Also, dark deposits within lysosome-like bodies appeared adjacent to the nuclei of some cells. Similar to the control group, the R-cells possessed electron lucent lipid droplets, approximately from 3.3-6.3 μm in diameter. Moreover, the mitochondria of many R-cells were deteriorated. Some of them appeared disorganized with a diameter ranged from 1.4-2 μm indicating normal size.
Our electron micrographs revealed that most of the F-cells [Fig. 4(B)] were cuboidal while some of them appeared filamentous. The average cell length was 23 μm and the average width was 16 μm indicating slight changes than normal which was 26.4 μm long and 13.5 μm wide. The nuclei of the F-cells were centrally located with a diameter ranged from 8-12.8 μm. Most of them were spherical in shape. The nuclei possessed a regular nuclear envelope with distinct nuclear membranes. The heterochromatin dominated over the euchromatin and was normally distributed. In addition, the cytoplasm of F-cells possessed well developed RER with obvious electron dense ribosomes [Fig. 4(B,C)]. The preparations of this group revealed also lysis of some areas in the cytoplasm. Lysosome-like bodies (average diameter was 5 μm) containing electron dense granules were present in the cytoplasm of some F-cells [Fig. 4(C)]. Furthermore, benzo(a)pyrene-treated specimens revealed the presence of many mitochondria with relatively lucent matrices and indistinct cristae.
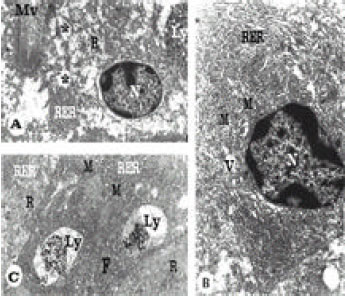 | |
| Fig. 4: | Transmission electron micrographs of the hepatopancreas of marine shrimp, Palaemon serratus intramuscularly injected with 100 ng μL-1 of benzo(a)pyrene showing: (A) R-cell with nucleus (N) having normal chromatin, numerous polymorphic voids (*), uniform apical microvillous border (Mv) and densely stained droplets accumulated within lysosome-like body (arrows). X3000. (B) F-cell with large polygonal nucleus (N) and well developed lamelliform RER. X4000. (C) F-cell with well-observed electron dense granules accumulated within lysosome-like bodies (Ly). Well developed RER with numerous mitochondria are also seen in the cytoplasm. R: R-cells. X4000. |
They were either spherical or oval in shape. Some preparations were characterized by disorganized mitochondria. Similar to that observed in the F-cells of the control group, mitochondria had a diameter ranged from 1.2-2 μm. The zymogen granules found in normal specimens could not be easily identified.
DISCUSSION
Our results demonstrated that the hepatopancreas of Palaemon serratus consists of numerous blind-ending tubules. This accord with the findings observed for Crangon crangon (Andersen and Baatrup, 1988). In addition, sections of electron microscopy were restricted to the mid-region of the hepatopancreas. Thus, the R- and F-cells were the only cell types present in the tubule and were fully differentiated. This observation confirmed the results of Davis and Burnett (1964) and Hopkin and Nott (1979, 1980) who reported that the hepatopancreatic tubule is divided into three zones: distal region containing E-cells, mid-region containing R- and F-cells and proximal region containing B-, R- and F-cells. Furthermore, the present study supports the findings of Loizzi (1971) concerning the hepatopancreas of the crayfish (Orconectes virilis and Procambarus clarkii), Lyon and Simkiss (1984) concerning the hepatopancreas of the crayfish (Austropotamobius pallipes) and Trinadaha Babu et al. (1989) concerning the hepatopancreas of the crab (Portunus sanguinolentus) in that the R-cells are the most abundant cell type in the hepatopancreas.
Ultrastructurally, perylene injection to the shrimps resulted in hepatopancreatic cellular disruption, disappearance of lateral cell membranes and folding of the basal lamina in some places. This might provide the morphological proof for the cell decomposing necrotic process (Asztalos et al., 1988). Besides, the hepatopancreatic cells of both christen and perylene injected shrimps lacked cellular organization. Similar change was reported in flounders chronically exposed to pulp mill effluents (Lehtinen et al., 1984), in rainbow trout and bullhead catfish exposed to coal derived materials (Stoker et al., 1985), in carp due to the effects of paraquat and methidation (Asztalos et al., 1988) and in tilapias collected from Lake Mariut (Abdel-Moneim, 1996).
Moreover, both perylene and christen injection caused necrosis, dilated and distorted appearance of the microvillous border of the hepatopancreatic cells. The length of these microvilli indicated reasonable decrease than normal. This effect may lead to the decrease of the absorptive capacity of the epithelial cells. Similarly, Kohler (1990) observed regression of the microvillous border of the space of Disse and severe atrophy of the endothelial sheets around the sinusoids in the hepatocytes of fish caught from the polluted Elbe estuaries. Nunez et al. (1991) in his study of the fine structure of hepatocellular neoplasms from aflatoxin B1 exposed rainbow trout mentioned that the presence of poorly developed microvilli in the space of Disse and in the bile canaliculi suggest loss of inter-relationships between hepatocytes and the microvascular system (sinusoids), between hepatocytes and the biliary system, and between individual hepatocytes, respectively. In addition, El-Elaimy et al. (1993) showed that the microvilli became irregular and collapse in liver of fishes exposed to neopybuthrin (pesticide).
Alteration in the heterochromatin content was generally observed after PAHs treatment. Abundant heterochromatin was observed in most nuclei of the hepatopancreatic cells of shrimps injected with either perylene or christen. The present finding is in agreement with Franchini et al. (1991) who studied the effect of acute lead exposure on hepatocyte ultrastructure of goldfish. Also, Hirano et al. (1991) reported condensation of the nuclear chromatin in cultured neonatal rat hepatocytes exposed to cadmium. Furthermore, some nuclei of hepatopancreatic cells of shrimps injected with perylene appeared with chromatin marginally distributed and condensed around the nucleoli. Faith and Trump (1965) suggested that with necrosis in vitro system chromatin tended to collect along the nuclear membrane and around the nucleoli and this was followed by fragmentation of chromatin strands and dissolution of interchromatin granules. In agreement with the present observation, earlier study of Gritzka and Trump (1968) revealed condensation and margination of chromatin after mercuric chloride intoxication. Similar change was also reported in hepatocytes of the white sea bass exposed to cadmium (Thophon et al., 2004). Heterochromatin condensation and marginalization suggest progressive inactivation of nuclear component (Braunbeck, 1994).
In parallel, few hepatopancreatic cells were also found with heterochromatin poor nuclei especially in shrimps injected with perylene. Rojik et al. (1983) found similar change in liver of fish exposed to copper sulphate and zinc chloride. El-Elaimy et al. (1993) also reported decreased chromatin content in liver nuclei of tilapias exposed to pesticide. The discrepancy observed in the content and distribution of chromatin present in the nuclei of both R- and F-cells in both perylene and christen injected shrimps can be explained by the concept of Ghadially (1985) who showed that the hyperchromatic nucleus may be of diagnostic importance as an indicator of polyploidy and the pale euchromatic nucleus with enlarged nucleolus is an indicator of active synthesis of rRNA which is important in active cell proliferation and growth of tumors. Moreover, other nuclear changes included dilation in the space between the nuclear membranes in either perylene and christen group. Stoker et al. (1985) found that the nuclear envelope lost its bilayer structure in hepatocytes of fish exposed to cool derived materials. Braunbeck et al. (1989) noticed similar observation in zebra fish exposed to 4-nitrophenol. In addition, El-Elaimy et al. (1993) observed rupture of nuclear envelope of liver of tilapias exposed to nopybuthrin. Severe deformation of nuclear membranes was also observed by Abdel-Moneim (1996) in hepatocytes of fish from heavily polluted lake. Furthermore, the nuclei of the F-cells of perylene injected shrimps possessed nuclear pocket-like structures resembling that described in tumor cells of vertebrate type 2 nuclear pocket. These structures arise by invagination of the inner nuclear membrane producing a meandering cleft, which sequestrates nuclear material (Ghadially, 1985).
A dominant feature in hepatic ultrastructure following exposure to petroleum aromatic hydrocarbons involves changes in the endoplasmic reticulum system (Faith and Fortner, 1987). A support of this assumption was obtained from the present study. F-cells of perylene injected specimens had extensive RER with disorganized arrangement and inconspicuous highly dilated lamellar cisternae. Similarly, Trump et al. (1965) have observed enlargement of RER cisternae in the in vitro necrotic system of mouse liver. It was also reported that the disorganization and dilation of RER are common reactions of liver cells to injury (Smuckler and Arcasoy, 1969) as well as during the period immediately following CCl4 poisoning (Reynolds and Ree, 1971). Hoffmann et al. (1974, 1975) also observed proliferation and dilation of the ER in liver cells following lead acetate and cadmium acetate intoxication. Rojik et al. (1983) postulated that the degree of damage in the liver of fish affected by copper sulphate was indicated by dilation and vacuolation of RER. In addition, Abdel-Moneim (1996) observed dilation and disorganization in liver of fish obtained from heavily polluted lake. Similar observation was reported in fish hepatocytes due to cadmium toxicity (Thophon et al., 2004). Dilation of RER, from water moving in and distorting it, is one sign of cell injury, causing the cell to stop functioning and leading to cell lysis (Cheville, 1994). In addition, fragmentation of RER cisternae with numerous polyribosomes in the cytoplasm indicating their detachment from the cisternae was also observed in some F-cells of christen injected specimens. The deformation in the RER was reported by Braunbeck et al. (1990) in liver cells of fish exposed to chemical spill. The disintegration of RER might be a consequence of final hyperactivity prior to cell necrosis (Roncero et al., 1992).
Ultrastructural observations revealed the presence of lysosome-like structures containing electron dense granules only in the R-cells of normal hepatopancreas. However, these structures could not be easily detected within the cytoplasm of hepatopancreatic cells either in perylene or in christen-injected shrimps. In contrast, benzo(a)pyrene treatment resulted in the presence of lysosome-like bodies in both R- and F-cells. The lysosomes in the F-cells of these preparations were significantly greater in number than those of the R-cells. This observation confirms with the study of Rojik et al. (1983), Segner (1987), Kohler (1989, 1990 and 1991), Gerundo et al. (1991) and Abdel Moneim (1996) who observed proliferation and enlargement of lysosomes in hepatocytes of fish exposed to pollutants. In general, hydrocarbons are known to accumulate in lysosomes (Kohler, 1989). Several authors suggested that when the storage capacity of lysosomes is exceeded, or as a result of direct damage to lysosomal membrane, hydrolytic enzymes are released and provoke degenerative processes and necrosis (Moore, 1985; Myers et al., 1987; Kohler, 1990). It would seen to be these degenerative processes that give rise to the final formation of numerous residual bodies which reflect the accumulation (Fowler, 1987) not of toxicants but of cell debris and undigested materials.
The F-cells of christen-injected animals showed large variability in distribution of mitochondria. Some of them were electron-dense and pyknotic with irregular shape, others were degraded. In most specimens, cristae could not be detected, however some of them possessed vesicular cristae. Mitochondrial changes represent a very clear demarcation of future pathological development (Segner, 1987) and the presently observed deterioration and degradation of mitochondria is supported by many investigations (Hoffmann et al., 1975; Bhatnagar et al., 1982; Franchini et al., 1991; Roncero et al., 1992). Ultrathin sections of christen injected group demonstrated the presence of electron-dense pleomorphic mitochondria "megamitochondria" in the R-cells, resulted due to fusion between small-sized mitochondria. A support for the present finding could be obtained from the study of Doughtie and Rao (1983). The mechanism by which mitochondria increase in size derived the attention of many investigator. It has been known that metal chelating reagents reduce the content of Ca2+ and Mg2+ in liver mitochondria and this may play a key role in megamitochondria formation (Wakabayashi et al., 1975). In addition, the enlargement of mitochondria might be due to loss of membrane integrity which led to fusion with other membranes of adjacent organelles as reported previously by Goyer (1968).
Also, in the current study, mitochondrial swelling was noted in some hepatopancreatic cells of the PAHs treated specimens. Mitochondrial swelling has been reported to follow virtually every type of cell injury that has thus far been studied. Hoffmann et al. (1975) reported similar change in liver and kidney of rats after cadmium toxicity. Initial mitochondrial matrix swelling was also observed in hepatocytes of fish exposed to several stressors (Rojik et al., 1983; Gerundo et al., 1991; Roncero et al., 1992; Abdel-Moneim, 1996; Thophon et al., 2004). Mitochondrial swelling might be due to pollutant-induced inhibition of Na+/H+ transporter and impairment of the overall osmoregulatory process of the cell (Vilella et al., 1991), thus swelling reflects the entry of solutes and water into mitochondrial matrix (Ghadially, 1988; Cheville, 1994).
Electron micrographs of hepatopancreatic cells of both perylene and christen injected animals showed the presence of large vacuoles containing cytoplasmic debris distributed all over the cytoplasm. The vacuoles were much larger in case of R-cells of christen treated group. These vacuoles probably arisen to digest the destructed cellular organelles as a result of PAHs treatments. The same vacuolation of the hepatocytes was described as a result of the effect of many toxicants such oil (Sabo et al., 1983), pulp mill effluents (Lehtinen et al., 1984; Axelsson and Norrgren, 1991), coal derived materials (Stoker et al., 1985), malachite green (Gerundo et al., 1991), CuSO4 (Roncero et al., 1992) and heavy metals/ petroleum oil (Abdel-Moneim, 1996). Asztalos et al. (1988) proposed that the focal development of empty vacuoles might be the starting point of cellular autolysis process.
In contrast to control group, lipid droplets could not be easily detected within the cytoplasm of the R-cells of both perylene and christen injected shrimps. A support of this finding reported by Lightner et al. (1982) who observed a marked reduction in the relative abundance of lipid vacuoles as a result of subacute aflatoxicosis. Similar observation was reported by El-Elaimy et al. (1988) who noticed marked depletion of total lipids in liver cells, brain and muscles of fish exposed to dimethoate malathione and hostathion. In contrast, Cruzan et al. (1986) and Khan et al. (1987) recorded rise of triglyceride total lipids and individual lipids in the liver of mouse subsequent to base oil and crude oil administration.
Shrimps (Palaemon serratus) is an economically important crustacean in Alexandria, therefore special care should be taken to protect this organism from polyaromatic hydrocarbon contaminated area. The finding of this study indicate that ultrastructural changes observer serve as biomarker in studies of toxic stress in aquatic environment.
REFERENCES
- Al-Mohanna, S.Y. and J.A. Nott, 1986. B-cells and digestion in the hepatopancreas of Penaeus semisulcatus (Crustacea: Decapoda). J. Mar. Biol. Assoc. UK., 66: 403-414.
Direct Link - Doughtie, D.G. and K.R. Rao, 1983. Ultrastructural and histological study of degenerative changes in the antennal glands, hepatopancreas and midgut of grass shrimp exposed to two dithiocarbamate biocides. J. Invertebrate Pathol., 41: 281-300.
CrossRef - Fowler, B.A., 1987. Intracellular compartmentation of metals in aquatic organisms: Roles in mechanisms of cell injury. Environ. Health Perspect., 71: 121-128.
PubMed - Hopkin, S.P. and J.A. Nott, 1979. Some observations on concentrically structured intracellular granules in the hepatopancreas of the shore crab Carcinus maenas (L.). J. Mar. Biol. Assoc. UK., 59: 867-877.
Direct Link - Ritterhoff, J., G.P. Zauke and R. Dallinger, 1996. Calibration of the estuarine amphipods, Gammarus zaddachi Sexton (1912), as biomonitors: Toxicokinetics of cadmium and possible role of inducible metal binding proteins in Cd detoxification. Aquat. Toxicol., 34: 351-369.
Direct Link - Rojik, I., J. Nemcsok and L. Boross, 1983. Morphological and biochemical studies on liver, kidney and gill of fishes affected by pesticides. Acta Biologica Hungarica, 34: 81-92.
PubMed - Roncero, V., E. Duran, F. Soler, J. Masot and L. Gomez, 1992. Morphometric structural and ultrastructural studies of tench (Tinca tinca L.) hepatocytes after copper sulphate administration. Environ. Res., 57: 45-58.
Direct Link - Scott-Fordsmand, J.J. and M.H. Depledge, 1997. Changes in the tissue concentrations and contents of calcium, copper and zinc in the shore crab Carcinus maenas (L.) (Crustacea: Decapoda) during the moult cycle and following copper exposure during ecdysis. Mar. Environ. Res., 44: 397-414.
CrossRef - Thophon, S., P. Pokethitiyook, K. Chalermwat, E.S. Upatham and S. Sahaphong, 2004. Ultrastructural alterations in the liver and kidney of white sea bass, Lates calcarifer, in acute and subchronic cadmium exposure. Environ. Toxicol., 19: 11-19.
CrossRefDirect Link - Vilella, S., V. Zonno, G. Cassano, M. Maffia and C. Storelli, 1991. Na+/H+ exchange in the kidney of eels Anguilla anguilla adapted to sea water or to freshwater environments: Studies with brush border membrane vesicles. Comp. Biochem. Physiol. Part A: Physiol., 102: 445-560.
Direct Link - Zilli, L., R. Schiavone, G. Scordella, V. Zonno, T. Verri, C. Storelli and S. Vilella, 2003. Changes in cell type composition and enzymatic activities in the hepatopancreas of Marsupenaeus japonicus during the moulting cycle. J. Comp. Physiol. B, 173: 355-363.
Direct Link - Braunbeck, T., 1994. Detection of Environmentally Relevant Concentrations of Toxic Organic Compounds Using Histological and Cytological Parameters: Substance-Specificity in the Reaction of Rainbow Trout Liver. In: Sublethal and Chronic Effects of Pollutants on Freshwater Fish, Maller, R. and R. Lloyd (Eds.). Blackwell Science Publishers, Boston, pp: 15-21.
- Neff, J.M., 1979. Polycyclic Aromatic Hydrocarbons in the Aquatic Environment: Sources, Fates and Biological Effects. Applied Science Publishers Ltd., London, pp: 7-71.
Direct Link