
E.U. Ezeji
Department of Biotechnology,
O. Obidua
Department of Biochemistry, University of Nigeria, Nsukka, Nigeria
I.G. Kalu
Department of Biotechnology,
I.N. Nwachukwu
Department of Microbiology, Federal University of Technology, P.M. B. 1526, Owerri, Nigeria
Pakistan Journal of Nutrition
Year: 2009 | Volume: 8 | Issue: 4 | Page No.: 414-418







ABSTRACT
Male and female albino mice were fed on maize (control) and gari (a dried cassava product) based diets for 5 weeks. Liver mitochondrial fractions were assayed for succinate dehydrogenase and Cytochrome C oxidase activities. There was an overall reduction in the activity of the two enzymes studied throughout the experiment. The reduction in activity was pronounced in Cytochrome c oxidase. Kinetic studies of succinate dehydrogenase showed an apparent increase in the Km of the gari fed animals-further supporting that inhibition took place. There was also a reduction in the mitochondrial respiratory control ratio (RCR) for the gari fed mice. The percentage reduction in RCR increased from 4.4% in the 2nd week to about 45% in the 5th week.
PDF Abstract XML References Citation
How to cite this article
E.U. Ezeji, O. Obidua, I.G. Kalu and I.N. Nwachukwu, 2009. Effect of Gari Diet on Marker Enzymes of Mice Liver Mitochondria. Pakistan Journal of Nutrition, 8: 414-418.
DOI: 10.3923/pjn.2009.414.418
URL: https://scialert.net/abstract/?doi=pjn.2009.414.418
DOI: 10.3923/pjn.2009.414.418
URL: https://scialert.net/abstract/?doi=pjn.2009.414.418
INTRODUCTION
Gari is a starchy food obtained from cassava processing. Cassava is a staple food in human diets in over 80 countries (Gomez et al., 1988). Gari is one of the most popular staple foods of the people of the rain forest belt of West Africa and contains mainly starch-20% amylase and 70% amylopectin-having lost the soluble carbohydrates (i.e. glucose and sugar) during processing.
The nutritional value (protein, fat and mineral contents) of gari is said to be low (Oketiku and Oyenuga, 1970). Continual dependence on gari (and other cassava related foods) as staple food may lead to protein and vitamin deficiencies (Grace, 1977).
A number of illnesses have been attributed to high and continuous consumption of cassava based diets. The cause of these illnesses is essentially the presence of toxic material in cassava-namely cynogenic glycosides and the coumarins. The cyanide content of the gari flour produced in Nigeria is about 10.6-22.1mg/kg (Nartey, 1980). Long-term consumption of cassava containing high levels of cynogenic glycocides has been associated with tropical ataxic neuropathy, spastic paraparesis and in areas with low iodine intake, development of hypothyroidism, goitre and cretinism (Osuntokun, 1981; Delang and Ermaus, 1971; Delang et al., 1971; JECFA, 1993; Abiye et al., 1998). Acute cassava poisoning-sometimes leading to the death of whole families has been reported after the consumption of inadequately processed cassava (Osuntokun, 1981; Cliff and Coutinho, 1995).
The mitochondria are a very important organelle in the animal cell. The primary role of the mitochondria is the conversion of ADP and inorganic phosphate into ATP in a reaction requiring energy. However, from the physiological viewpoint, the mitochondria can be regarded as biochemical “machines” that recover the energy contained in foodstuffs (through the TCA cycle and the respiratory chain) and convert it by phosphorylation into the high energy bond of ATP. The ATP so formed is then exported for use in the rest of the cell.
There are more than 70 enzymes and coenzymes distributed in all the components of the mitochondria. However, most of these enzymes are also present in other sub-cellular organelles, with only a few present exclusively in the mitochondria. These few enzymes have been referred to as “Marker enzymes” of the mitochondria. Being unique to the mitochondria these enzymes are used for the identification of the mitochondrial fraction in any cell fractionation experiment. They are also used to detect possible contamination of other sub-cellular fractions by the mitochondrial fraction. Two of the marker enzymes of the mitochondria are succinate dehydrogenase and cytochrome c oxidase both of which are located in the inner mitochondrial membrane. The activities of both enzymes are affected by the presence of several toxins, including cyanide.
It has been reported that cyanide ion inhibits tissue cytochrome oxidase activity in rats, mice and rabbits with resulting anoxia (Way, 1984; USEPA, 1988). Conn (1973) and Debey (1975) have earlier demonstrated that cyanide causes inhibition of cytochrome oxidase due to its high affinity for Fe3+. Consequently, it blocks the electron transport through the cytochrome system and thus oxygen uptake by the tissues. Death usually occurs from tissue asphyxia notably affecting the central nervous system.
This paper reports the effect of gari based diet on the activities of mice liver succinate dehydrogenase and cytochrome oxidase.
MATERIALS AND METHODS
Animals: Twenty-six healthy albino mice weighing between 15-35g were acquired from the department of Veterinary Medicine, University of Nigeria, Nsukka. The animals were separated into two groups A and B, representing control and experimental, respectively. Each group contained 13 mice.
Feeds: Group A animals (control) were fed on maize based diet while group B animals (experimental) were fed on gari based diets for a period of 5 weeks. The feeds for the experimental animals comprised of gari (70%), Groundnut cake (20%) and crayfish (10%). For the control animals, the feeds comprised of maize (70%), groundnut cake (20%) and crayfish (10%). All the components of the feeds were bought from Nsukka market. The gari was first prepared in boiled water, dried and then made light and crisp again by grinding before use. The process yields what is known as “prepared gari”.
Animals from each group were however removed and the liver mitochondria isolated for analysis in the 2nd, 3rd, 4th and 5th weeks respectively.
Chemicals and reagents: All chemicals and reagents used were of high analytical grade. Bovine serum albumin (BSA) was obtained from Riedel-De Haem Company, Germany. Copper sulphate crystals were obtained from Merck, Germany. Adenosine diphosphate (ADP) was obtained from Sigma Chemicals Company Ltd, England. Sucrose from Hopkin and Williams, England; Sodium Succinate from British Drug House, England and Sodium hydroxide pellets from May and baker Ltd, England. The rest of the chemicals and reagents were obtained from BDH Chemicals Ltd, England.
Preparation of liver homogenate: The mice were sacrificed by stunning and their liver were removed and immediately placed in ice cold isolation medium, pH 7.4, consisting of 0.33M Sucrose, 0.025mM K-EDTA and 15mM tris-HCl. The liver samples were minced with a sharp pair of scissors. The blood was washes off and the pieces of liver homogenized in a suitable volume of isolation medium for about 3 minutes using an electrically operated Lab. Type homogenizer (Ultra-Turrax T25).
Isolation of liver mitochondria: The mitochondria were isolated using a modification of the method published by Guerra (1979). The liver homogenate was placed in centrifuge tubes and spun in a refrigerated centrifuge (Sigma 2K15) at 800g for 10 minutes. The supernatant was carefully collected and centrifuged at 8200g for 10 minutes. The resultant supernatant was discarded. The pellets were suspended in about 20ml of isolation medium and further centrifuged at 8200g for 10 minutes. The pellets were suspended in about 20ml of suspension medium consisting of 0.33M Sucrose and 6mM magnesium chloride (to replenish the magnesium ions in the mitochondria) and centrifuged at 8200g for 10 minutes. The final pellets were suspended in the suspension medium to a final volume of 5ml. All mitochondrial isolation procedures were performed at 0-4oC. Mitochondrial protein concentration was spectrophotometrically estimated with the biuret method using bovine serum albumin as standard.
Measurement of mitochondrial oxygen uptake: The mitochondrial oxygen uptake (respiratory control ratio, RCR) was measured according to the method due to Estabrook, et al. (1967). This method involves the polariographic measurement of the volume of oxygen consumed by the mitochondria in metabolic states 3 and 4 using a Model 53 biological oxygen monitor (YSI) carrying a YSI 5331 Oxygen probe. The meter was calibrated with 3ml of an air saturated basal reaction medium containing 150mM sucrose; 10mM K2PO4 buffer, pH 7.4; 5mM MgCl2; and 20mM KCl. After calibration, the meter reading was set at 100%; 0.3ml of mitochondrial suspension was added to the medium in the cell, and allowed to respire until a steady meter reading was maintained. Mitochondrial respiration was initiated by adding 25μL of substrate (Na-succinate) and the rate of oxygen consumption was measured (State 4). State 3 respiration was determined after adding 25μL ADP.
The respiratory control ratio (RCR) was calculated as the ratio between State 3 and State 4 respiration using 235 nmol O2/mol as the value for the solubility of oxygen at 25oC (Estabrook et al.,1967). The degree of coupling of oxidative phosphorylation (q) was calculated using the equation: q = 1-1/RCR (Rottenberg et al., 1985).
Enzyme assay
Succinate dehydrogenase activity: Succinate dehydrogenase activity was measured spectrophotometrically as a decrease in optical density at 400nm according to the method of Slater and Bonner (1952). At zero time, 0.2ml of suitably diluted mitochondrial suspension was added to the reaction mixture consisting of 0.3ml of each of KCN and K3Fe3(CN)6, 0.2ml of 0.2M Na-succinate and 2.0ml of 0.15M sodium phosphate buffer, pH 7.4 to make a total of 3.0ml. The O.D. was read at one minute intervals for 5 minutes. The specific activity was calculated as the rate of change in O.D/min/mg protein [molar extinction coefficient (ε) at 400nm (ε 400) = 1.0mM–1cm–1].
Cytochrome c oxidase activity: The enzyme was assayed by the method due to Simon (1958). The mitochondrial suspension (0.2ml) was pipetted into a cuvette containing 1.5ml of 0.1M potassium phosphate buffer, pH 7.1, 0.1ml of 0.2M sucrose solution and 0.2ml of 20mM ferrocytochrome C. The contents of the cuvette were mixed and the change in absorbance at 550nm was monitored at 30 seconds intervals for 5 minutes against a reagent blank. The specific Cytochrome c oxidase activity was calculated as change in O.D/min/mg protein [molar extinction coefficient (ε) at 550nm (ε550) = 19.2mM–1cm–1].
Mitochondrial protein determination: The protein content of the mitochondrial extract was spectrophotometrically assayed using bovine serum albumin as standard according to Lowry et al. (1951).
RESULTS AND DISCUSSION
There was an overall reduction of mitochondrial respiratory control ratio (RCR) in gari fed mice from 2.15 in week 2-1.96 in week 5. The RCR value for the control however increased from 2.25 in week 2-3.23 in week 5 (Fig. 1). RCR is recognized as the best indicator of the quality of the mitochondrial preparation and of the degree of coupling (or uncoupling) of oxidative phosphorylation. A plot of the degree of coupling of oxidative phosphorylation (q) against time in weeks (Fig. 2) shows that the experimental animals exhibited a lower degree of coupling at the end of the period of study. In both the control and experimental, introduction of ADP led to a marked increase in oxygen uptake. This was however, greater in the control than in the experimental as shown in Fig. 3.
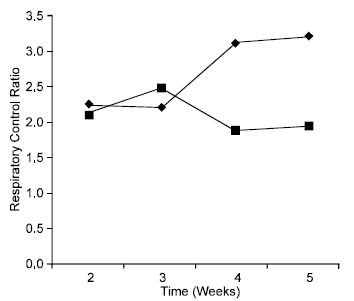 | |
| Fig. 1: | Liver Mitochondrial Respiratory Control Ratio (RCR) of Gari (■) and maize (♦) fed mice, respectively. |
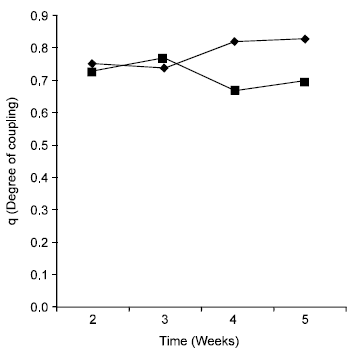 | |
| Fig. 2: | Degree of coupling of oxidative phosphorylation of Gari (■) and maize (♦) fed mice, respectively. |
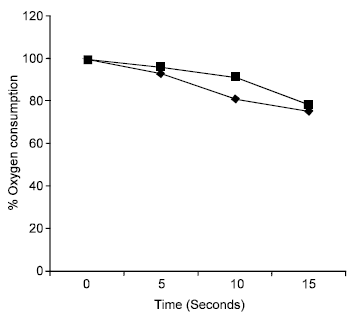 | |
| Fig. 3: | Mitochondrial Oxygen consumption of Gari (■) and maize (♦) fed mice, respectively. |
Figure 4 shows the effect of gari feeding on the specific activity of mitochondrial succinate dehydrogenase. There was a general fall in succinate dehydrogenase activity as a result of prolonged gari feeding. This fall was however, not consistent during the period of the study. In both the control and experimental the specific activity decreased as the feeding progressed but that of the control increased again in the 5th week. The mechanism of inhibition of succinate dehydrogenase is still not clear, but it has been suggested that the enzyme is irreversibly inactivated by cyanide. The inhibition of succinate dehydrogenase was further confirmed by the Km of the enzyme which was apparently higher in the gari fed animals. Figure 5 shows that the Km (Michaelis constant) of the enzyme increased from 0.029M (in wk.2) to 0.0775M (in wk.5) and 0.0188M (in wk. 2) to 0.055M (in wk. 4) for the experimental and control animals respectively.
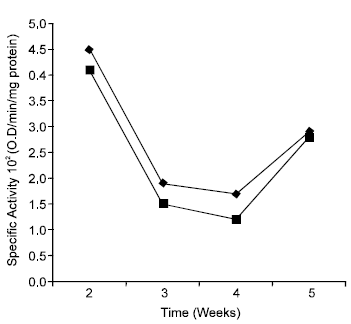 | |
| Fig. 4: | Specific activity of Succinate dehydrogenase of Gari (■) and maize (♦) fed mice, respectively. |
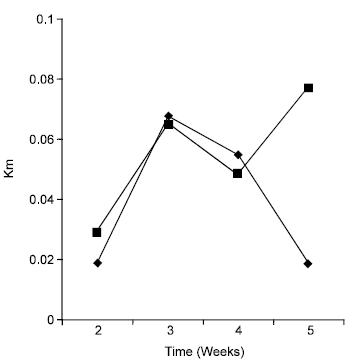 | |
| Fig. 5: | Km of succinate dehydrogenase of Gari (■) and maize (♦) fed mice, respectively. |
The Km for the control however reduced again in wk. 5.
The activity of Cytochrome C oxidase showed a decrease between the 2nd and 4th weeks of the experiment in the gari fed animals, but increased again in the 5th week (Fig. 6). The lowest activity in both the experimental and control animals, respectively, was observed in the 4th week.
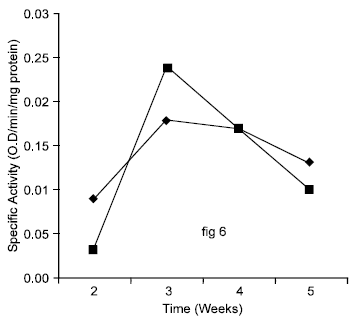 | |
| Fig. 6: | Specific Activity of Cytochrome c oxidase of Gari (■) and maize (♦) fed mice, respectively. |
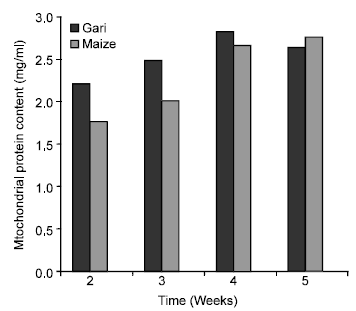 | |
| Fig. 7: | Estimation of protein content of Liver Mitochondria of Gari and maize fed mice, respectively. |
Investigations have shown that cyanide is a potent inhibitor of cytochrome C oxidase, an enzyme that functions at the terminal end of the electron transport chain (Solomonson, 1981; Leduc et al., 1982; Delhumeau et al., 1994). The results of this study further supports this claim. Chilaka et al. (1985) reported a 43% inhibition of the activity of NADPH-cytochrome c reductase in gari fed mice.
Figure 7 shows the effect of gari based diet on liver mitochondrial protein concentration. The liver protein concentration of gari fed mice were generally lower than that of the corn fed ones, except in the week 5 of the experiment.
In conclusion, this study suggests that cyanide affects some important enzymes of the mitochondrial electron transport system namely, succinate dehydrogenase and cytochrome c oxidase, both of which are used as markers of the organelle. It also showed that the rate of oxygen consumption is lowered upon prolonged gari feeding.
REFERENCES
- Abuye, C., U. Kelbessa and S. Wolde-Gebriel, 1998. Health effects of cassava consumption in South Ethiopia. East Afr. Med. J., 75: 166-170.
PubMed - Chilaka, F.C., E.O. Anosike and O. Obidoa, 1985. Effect of high and prolonged gari diets on some microsomal enzymes activities of rat liver. Plant Foods Human Nutr., 35: 159-164.
CrossRefDirect Link - Delang, F. and A.M. Ermans, 1971. Role of a dietary goitrogen in the etiology of endemic goiter on Idjwi Island. Am. J. Clin. Nutr., 24: 1354-1360.
Direct Link - JECFA, 1993. Cyanogenic glycosides: Toxicological evaluation of certain food additions and naturally occurring toxicants. Geneva, World Health Organization, 39th Meeting of the Joint FAO/WHO Expert Committee on Food Additives (WHO Food Additives series 30). http://bases.bireme.br/cgi-bin/wxislind.exe/iah/online/?IsisScript=iah/iah.xis&src=google&base=WHOLIS&lang=p&nextAction
- Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Simon, E.W., 1958. The effect of digitonin on cytochrome C oxidase of plant mitochondria. Biochem. J., 69: 68-74.
Direct Link - Slater, E.C. and W.O. Bonner, 1952. The effect of fluoride on the succinate dehydrgenase system. Biochem. J., 52: 185-196.
Direct Link - Way, J.L., 1984. Cyanide intoxication and its mechanism of antagonism. Annu. Rev. Pharmacol. Toxicol., 24: 451-481.
Direct Link