
Rowaida Khalil
Department of Botany, Faculty of Science, Alexandria University, Alexandria, Egypt
Yasser Elbahloul
Department of Botany, Faculty of Science, Alexandria University, Alexandria, Egypt
Fatima Djadouni
Department of Botany, Faculty of Science, Alexandria University, Alexandria, Egypt
Sanaa Omar
Department of Botany, Faculty of Science, Alexandria University, Alexandria, Egypt
Pakistan Journal of Nutrition
Year: 2009 | Volume: 8 | Issue: 3 | Page No.: 242-250







ABSTRACT
A Bacillus megaterium 19 strain isolated from a mixture of fermented vegetable wastes, produced a bacteriocin that displayed a wide spectrum antimicrobial activity against food-spoilage microorganisms and possessed a bactericidal mode of action. The bacteriocin activity against Salmonella typhimurium and Staphylococcus aureus attained its maximum value during the mid stationary phase. Maximal bacteriocin activity (93% growth reduction) was achieved using MRS broth, at initial pH of 5.5-6.5 and at 30°C incubation temperature. High levels of bacteriocin activity (up to 85% growth reduction) were detected under limited or reduced oxygen levels. The inhibitory activity increased in mMRS broth with 4% sucrose and (0.10%) beef extract. The bacteriocin was thermally stable over a wide temperature range up to 100°C for 15 min and retained its activity at acidic and neutral pH values, but not at alkaline pHs. Full bacteriocin activity was expressed after 30 days of storage at 4°C, for 15 min of exposure to UV light. The bacteriocin was amylase and lipase sensitive, however, treatment with proteolytic enzymes resulted in a remarkable stability. SDS-PAGE analysis of the partially purified bacteriocin revealed an apparent molecular weight ranging from 3.496 to 6.512 kDa. The strain may have a potential use as food biopreservative, because of its promising thermostable technological properties and broad antimicrobial spectrum.
PDF Abstract XML References Citation
How to cite this article
Rowaida Khalil, Yasser Elbahloul, Fatima Djadouni and Sanaa Omar, 2009. Isolation and Partial Characterization of a Bacteriocin Produced by a Newly Isolated Bacillus megaterium 19 Strain. Pakistan Journal of Nutrition, 8: 242-250.
DOI: 10.3923/pjn.2009.242.250
URL: https://scialert.net/abstract/?doi=pjn.2009.242.250
DOI: 10.3923/pjn.2009.242.250
URL: https://scialert.net/abstract/?doi=pjn.2009.242.250
INTRODUCTION
Alternate methods for controlling pathogenic bacteria including the production of antimicrobial peptides “bacteriocins” are now highly considered. Bacteriocins are compounds produced by bacteria that have a biologically active protein moiety and a bactericidal action (Line et al., 2008). Bacteriocins from gram-positive organisms, such as lactic acid bacteria (LAB), have attracted much attention and have been the subject of intensive investigation due to their extensive incorporation as biopreservatives ingredients into model foods particularly in the dairy industry (Diop et al., 2007) and also in human therapeutics (Martin-Visscher et al., 2008). Bacillus is an interesting genus to investigate since it produces a diverse array of antimicrobial peptides representing several different basic chemical structures (Bizani and Brandelli, 2002). The production of bacteriocins or bacteriocin-like substances has been already described for some bacilli such as B. subtilis, B. cereus, B. stearothermophilus and other Bacillus species. Some strains produce bacteriocins with a broad spectrum activity, including important pathogens such as Listeria monocytogenes and Streptococccus pyogenes (Cherif et al., 2001). Some were well characterized such as: lichenin produced by B. licheniformis 26-103 RA strain (Pattnaik et al., 2001) and megacin produced by B. megaterium (Lisboa et al., 2006). In spite of the considerable approaches taken by researchers to gather data on Bacillus bacteriocins, the importance and the industrial value of those bacteriocins has been largely underestimated and has attracted little attention compared to those produced by LAB. Hence, the present study primarily aimed at screening for bacteriocinogenic bacilli strains using agro-based wastes. A bacilli strain isolated from fermented vegetable wastes showed strong inhibitory activity, a finding that prompted us to evaluate the potential of the bacteriocin produced. The bacteriocin was identified and partially characterized. The results reported in the current study may be useful in revealing the possibility of potential use of the bacteriocin as an antimicrobial agent to prevent the proliferation of some pathogenic and food-spoilage microorganisms and to improve the microbial quality and safety of foods.
MATERIALS AND METHODS
Bacterial strains: The pathogenic indicator strains used in this work were provided by the laboratory of Bacteriology, Microbiology Department at the Faculty of Medicine, Alexandria University. They included both gram negative (Escherichia coli, Pseudomonas aeruginosa, Proteus vulgaris, Klebsiella pneumoniae, Salmonella typhimurium, Salmonella para-typhimurium A, Salmonella para-typhimurium B and Enterococcus faecalis) and gram positive strains (Streptococcus pyogenes and Staphylococcus aureus).
Isolation of bacilli strains: Bacilli strains were isolated from agro-industrial wastes (including vegetables, fruits and fermented vegetable wastes). Samples were weighed as 1 g portions and thoroughly homogenized in sterile distilled water, serial dilutions were plated on LB agar (Difco Laboratories, Detroit, MI), plates were incubated at 30°C for 2-3 days.
Inoculum preparation and growth conditions: Inocula of the test strain (1% v/v) for all experiments consisted of cellular suspension from 12-18 h cultures on MRS (Biolife Italiana S.V.L) broth, adjusted to an absorbance (600 nm) of 1-1.2. For storage of stock cultures, cells were separated by centrifugation (8,000xg, 20 min at room temperature) and resuspended in 1 mL of fresh MRS liquid medium, from which 0.5 mL portion was transferred to 2 mL beads-containing cryogenic vials, freezed at -80°C in the presence of 15% (v/v) glycerol. Inocula of the indicator strains consisted of cellular suspension from MRS broth cultures that were monitored spectrophotometrically till an O.D.600 corresponding to 105 CFU/mL was reached (Naclerio et al., 1993).
Screening for bacteriocin-producing bacilli: Bacilli isolates were screened for bacteriocin production by the Agar Well Diffusion (AWD) assay (Lasta et al., 2008).
Identification of the bacteriocinogenic isolate: The pure isolate selected as a potential bacteriocin-producer was identified on the basis of its cultural, morphological, physiological and biochemical characteristics (Schillinger and Lücke, 1987), followed by partial 16S rRNA analysis (Maidak et al., 1999) conducted at the German culture collection of microorganisms and cell cultures DSMZ (Deutch Sammlung von Mikroorga-nismen und Zelkulturen GmbH).
Detection of bacteriocin activity during growth (Growth kinetics): MRS broth was inoculated with 2% (v/v) of an overnight preculture of the test strain and incubated at 30°C. Changes in O.D.600 were recorded every 3 h. The growth kinetics experiment (Vinderola et al., 2002) was employed with slight modification. The indicator strains (105 CFU/mL) were grown at 30°C in MRS broth in the presence of a concentrated cell-free supernatant (CFS) of the bacteriocinogenic strain. Optical Density Measurements (ODM) were recorded every 3 h for 12-18 h. The bacteriocin activity was expressed by the percentage of growth reduction of the indicator pathogenic strains and determined from the ratio between the optical densities of the treated cultures and untreated ones (the indicator strains without the CFS).
Effect of growth media, incubation temperature, initial pH and aeration on bacteriocin activity: Commercial media such as MRS (De man Rogosa Sharpe) broth, Brain Heart Infusion (BHI; Merck, Darmstadt, Germany) broth, M17 (Difco Laboratories, Detroit, MI) broth, whey and molasses (2% v/v) were tested for their suitability for bacteriocin activity using the ODM method. The test strain was inoculated into each test medium, grown aerobically at 2 selected temperatures (30 and 37°C). Cells were harvested by centrifugation and the resulting CFSs were mixed with the indicator strains and the bacteriocin antimicrobial activity was determined using the ODM method. To determine the effect of pH, aliquots of MRS broth were adjusted with 1N HCI or 1N NaOH to pH values of 4.5, 5.0, 5.5, 6.0, 6.5, 7.0 and 7.5 (Ougnbanwo et al., 2003a). Aliquots were autoclaved, inoculated with the bacteriocinogenic strain and aerobically incubated. Cultures were assayed for bacteriocin activity as described above. The effect of aeration conditions on bacteriocin activity was studied by varying the volume of the growth medium used. Aliquots of 10, 20, 40 and 50 mL of a modified version of MRS broth (mMRS) were inoculated with the test strain and assayed for bacteriocin activity as mentioned elsewhere.
Effect of sugar and nitrogen sources on bacteriocin activity: The strain was grown for 15, 18 and 24 h in mMRS broth supplemented with 2 and 4% of monosaccharide (galactose) and disaccharide (fructose, lactose, maltose, sucrose,) sugars, respectively, after replacing the medium glucose (Ogunbanwo et al., 2003b). The effect of nitrogen sources on bacteriocin activity was evaluated using (per lit): 4 g ammonium acetate, 4 g ammonium chloride, 4 g ammonium nitrate, 4 g ammonium sulfate, 4 g arginine, 10 g beef extract, 4 g sodium nitrate, 10 g tryptone 10 and 5 g yeast extract. The effect of nitrogen combinations were also studied: beef extract plus yeast extract, beef extract plus tryptone, beef extract plus ammonium chloride, beef extract plus arginine. Combinations were added as 1.5 and 0.5%, respectively.
Physical and biochemical characterization of the bacteriocin
Heat resistance and pH sensitivity: The thermal stability of the crude bacteriocin was assessed by exposing aliquots of the test strain CFS to different temperatures (0, 30, 40, 50, 60, 70, 80, 90, 100 and 121°C) for 15 min (Mota et al., 2004), then tested for antimicrobial activity. The effect of pH on the bacteriocin activity was tested by adjusting test strain CFS to pHs from 2-12 (at increment of one pH unit) with sterile 1 N NaOH or HCl (Albano et al., 2007). Samples were incubated at room temperature (25°C) for 1 h then assayed for antimicrobial activity.
Stability during storage and exposure to UV light: The crude bacteriocin was stored at -20 and 4°C for different intervals of time (30, 45 and 60 days). Sterile Petri dishes containing aliquots of 10 mL crude bacteriocin preparations were exposed to UV irradiation (Philips bulb, wave length 340 nm, 220-240 V, 50 Hz,) situated at a distance of 30 cm from the Petri dishes (Wanda and Bonita, 1991). Times of exposure to UV light ranged from 15-90 min, after which the bacteriocin activity was estimated (Ogunbanwo et al., 2003a).
Sensitivity to proteolytic and other enzymes: Proteolytic enzymes (Oxford laboratory reagents) including papain, pepsin, trypsin and non-proteolytic enzymes such as lipase and φamylase were dissolved in 0.05 M sodium phosphate (pH 7.0), 0.002 M HCl (pH 7), 40 mM Tris-HCl (pH 8.2), 0.1 M potassium phosphate (pH 6.0) and 0.1 M potassium phosphate (pH 7.0), respectively to a final concentration of 1 and 2 mg/mL. Enzyme solutions were sterilized by disposable bacterial filters (0.2 µm; Fischer chemicals, UK). Aliquots of filter sterilized CFSs of the test strain were mixed with each enzyme solution, incubated at 30°C for 2 h, subsequently heated in boiling water for 5 min to inactivate the enzymes (Bizani and Brandelli, 2002) and finally assayed for antimicrobial activity.
Effect of organic solvents on bacteriocin activity: The sensitivity of the bacteriocin to organic solvents such as acetone, chloroform, ethyl alcohol, hexane and methanol was investigated (Todorov et al., 2006). Freeze dried bacteriocin preparations from the producer strain were dissolved individually in each organic solvent at a final concentration of 10 mg/mL. Samples were incubated at 30°C for 1 h, then solvents were removed by evaporation. Dried residues from the organic phase were re-suspended in sterile MRS broth (Ten Brink et al., 1994) at a final concentration of 10 mg/mL and assayed for antimicrobial activity.
Mode of action on target cells: About 20 mL of filter sterilized CFS of the test strain was mixed with 100 mL of S. typhimurium culture (105 CFU/mL), incubated at 30°C for 12-18 h. Cells were harvested by centrifugation at 8,000xg at room temperature for 20 min, washed twice with sterile saline water and re-suspended in 10 mL (Albano et al., 2007) and was further processed according to Delmar et al. (2005) and observed with a JEOL JEM 1200ExII transmission electron microscope (JEOL, Tokyo) operating at 120 Kv.
Partial purification and molecular weight determination: The test strain was grown in MRS broth for 18 h at 30°C. Cells were harvested by centrifugation at 10,000xg for 20 min at 4°C, after which the bacteriocin was precipitated from the CFS with 45% saturated ammonium sulfate (Akyptis et al., 1998). The molecular weight of the bacteriocin was estimated according to the method of Sambrook and Russell (2001), using discontinuous SDS-PAGE performed using a double slab electrophoresis cell (Cleaver scientific Ltd). Molecular weight markers (Bio-RAD, Germany) ranging from 90-1.434 kDa were used. The gels were fixed and stained with Coomassie Brilliant Blue R 250 (Saarchem, Krugersdorp, South Africa). The apparent molecular mass of the sample was calculated by comparison with the mobility of the standard markers.
Statistical analysis: Data were expressed as mean±Standard deviation. Statistical significance was determined using the student’s t-test. p<0.05 was considered significant.
RESULTS
Screening and identification of bacteriocinogenic strains: About 30 isolates were obtained and screened for antimicrobial spectrum against gram-positive and gram-negative bacteria using the AWD method. The average diameter of the inhibition zones measured ranged from 0.5-5 mm in size (data not shown). Among the isolates, one strain (19) was isolated from fermented vegetable wastes, identified as B.megaterium by physiological and biochemical characteristics and further confirmed by 16S rRNA gene sequence analysis that showed 100% sequence identity with Bacillus megaterium DSM 32.
Growth curve and growth kinetics: Low bacteriocin activity against S. typhimurium was noted during the lag phase (almost 15% growth reduction). The maximal antimicrobial activity against the indicator strains S. typhimurium and S. aureus (86.54 and 86.53% growth reduction, respectively) was clearly demonstrated during the stationary phase and peaked at its mid after 15 h (Fig. 1).
Detection of antimicrobial spectrum: Results of the AWD method showed that the B. megaterium 19 CFS contained an antimicrobial compound with a wide spectrum that inhibited the growth of representatives of at least three indicator gram positive or gram-negative strains (data not shown). Inhibition zones of size 5 mm were detected.
Effect of growth media and incubation temperature on bacteriocin activity: The highest bacteriocin inhibitory effect was obtained in MRS broth after 12-18 h of incubation at 30°C. The activity was maintained after 21 h, where approximately 93% reduction of growth was recorded (Fig. 2). Good antimicrobial activity was recorded in the presence of 2% (w/v) molasses. BHI medium was not a suitable medium for bacteriocin production. Increasing the incubation temperature had a significant adverse effect on bacteriocin activity (data not shown).
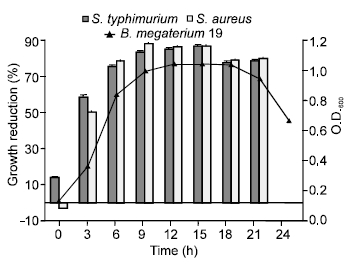 | |
| Fig. 1: | Growth kinetics of S. typhimurium (□) and S. aureus (■). (▲) is the growth of B. megaterium 19 in MRS broth, incubated aerobically at 30°C for 24 h. O.D.600 values are averages of triplicate measurements (n = 3). Error bars represent the standard deviation of mean values of % growth reduction of bacteriocin preparations to each indicator strain |
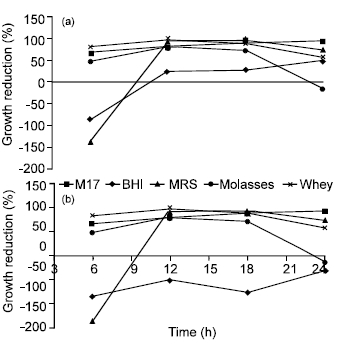 | |
| Fig. 2: | Influence of growth media on the activity of B. megaterium 19 bacteriocin against the indicator strains S. typhimurium (a) and S. aureus (b) when grown in MRS broth at 30°C, pH: 6.2-6.5. Results are expressed as percent of mean values of growth reduction (n = 3) ±standard deviations |
Effect of initial medium pH and aeration conditions on bacteriocin activity: The activity of the bacteriocin was incubation time and pH dependant. The activity was highly maintained in the acidic pH range of 4.5-5 (45-65% growth reduction). The highest antimicrobial activity (95% growth reduction) was demonstrated when MRS broth was adjusted to an initial pH of 5.5-7 (Fig. 3). At pH 7.5 and above, the bacteriocin was significantly (p>0.05) reduced after 12 h of incubation. The test strain produced bacteriocin under limited or reduced aeration. The activity increased with the increase in culture volume (decreasing the oxygen availability) from 20 mL up to 50 mL (data not shown).
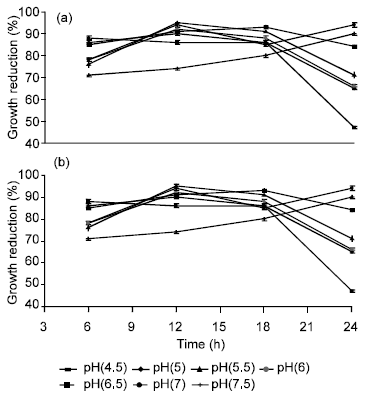 | |
| Fig. 3: | Activity of bacteriocin produced by B. megaterium 19 against (a) S. typhimurium and (b) S. aureus as influenced by the medium initial pH. Results are expressed as percent of mean values of growth reduction (n = 3) ±standard deviations |
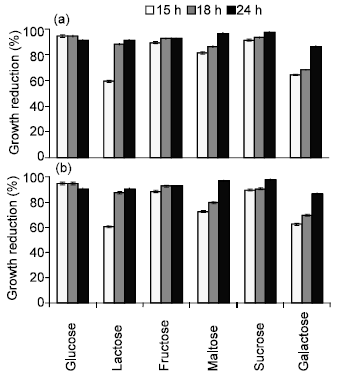 | |
| Fig. 4: | Effect of carbon sources on the activity of bacteriocin produced by B. megaterium 19 against (a) S. typhimurium and (b) S. aureus. Results are expressed as percent of mean values of growth reduction (n = 3) ± standard deviations |
Effect of carbon and nitrogen sources on bacteriocin activity: Optimal bacteriocin activity (97% growth reduction) was recognized after 24 h of incubation in mMRS supplemented with 4% sucrose, followed by glucose, maltose, fructose and finally lactose (Fig. 4). Galactose did not stimulate the bacteriocin production. The maximum inhibition to the growth of the indicator strains was demonstrated in mMRS broth supple-mented with beef extract (10 g/L) after 24 h of incubation, whereas a combination of beef extract and yeast extract resulted in 98% reduction of growth (data not shown).
Physical and biochemical characterization
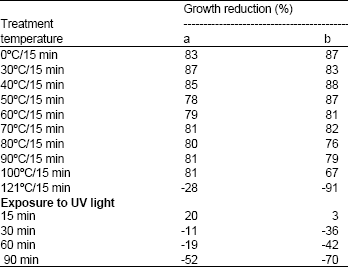
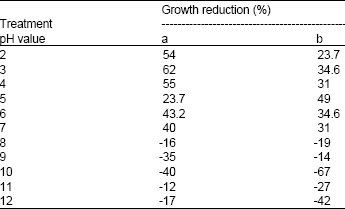
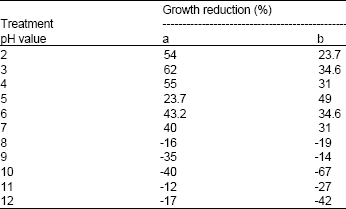
Thermal, UV light treatment, pH sensitivity and stability during storage: Results in Table 1 show that the inhibitory compound produced by B. megaterium 19 was heat stable upon exposure to 100°C for 15 min, but was sensitive to autoclaving temperature (121°C). The bacteriocin retained its activity after 15 min of exposure to UV light. The bacteriocin was also stable at pH 2-7 after 15 min, but the activity was totally lost at alkaline pH value of 8 and above (Table 2). The bacteriocin maintained high stability and showed maximal growth reduction (88%) after 30 days of storage at 4°C (Table 3). Exposing the bacteriocin to longer periods of storage (45 days) resulted in an insignificant decrease in activity by 10%; however the activity was restored after 90 days of storage and up to a maximum period of 12 months (data not shown). Hardly, any bacteriocin was produced at -20°C after 45 days and up to 90 days.
Sensitivity to enzymes and solvents: S. thermophilus 19 bacteriocin activity was insensitive to the two tested concentrations of papain (Table 4). The effect of pepsin and trypsin on bacteriocin activity at concentrations of 2 mg/mL was more pronounced compared to 1 mg/mL. Treatment with either concentrations of amylase inhibited the bacteriocin activity against S. aureus more than S. typhimurium. The reverse pattern was shown upon treatment of the bacteriocin with lipase. With an exception to hexane and ethanol, the bacteriocin was completely sensitive to solvents employed in the study (data not shown).
Mode of action: Figure 5a shows intact cells and typical rods of S. typhimurium before treatment with the bacteriocin preparation. Figure 5b illustrates the lysed ghost cells of S. typhimurium treated with the bacteriocin, where cells surfaces were damaged due to pore formation, in addition to an alteration in the cell morphology, confirming the bactericidal action of the bacteriocin..
| Table 1: | Effect of thermal and UV light treatment on bacteriocin activity against (a) S. typhimurium and (b) S. aureus. Results are expressed as % of mean values of growth reduction |
 | |
| Table 2: | Influence of pH on bacteriocin activity expressed as % of mean values of growth reduction to (a) S. typhimurium and (b) S. aureus |
 | |
| Table 3: | Effect of time and temperature of storage on bacteriocin activity produced by B. megaterium against the test indicator strains. Results are expressed as % of mean values of growth reduction (n = 3) |
 | |
| Table 4: | Effect of enzyme treatment and concentration on bacteriocin activity against (a) S. aureus and (b) S. typhimurium. Results are expressed as % of mean values of growth reduction (n=3) ± standard deviations |
 | |
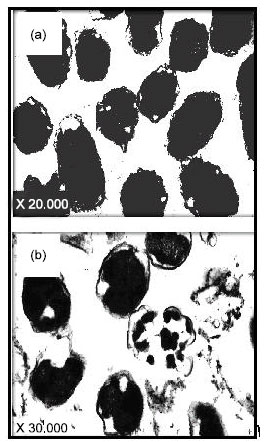 | |
| Fig. 5: | Transmission Electron Micrographs (TEM) of S. typhimurium (105 CFU/mL). Control cells were left untreated (a) or treated with bacteriocin preparation of B. megaterium 19 (b). Control cells remained as intact, smooth and undamaged rod shaped cells, while treated ones appeared as ghost cells with a deformed morphology and distorted cell surfaces |
Determination of bacteriocin molecular weight: The molecular weight of the partially purified bacteriocin was estimated to range from 3.496-6.512 kDa.
DISCUSSION
This study describes the partial characterization of a bacteriocin produced by a B. megaterium 19 strain isolated from fermented vegetable wastes. Choice was based on its broad antimicrobial spectrum against more than three genera including S. aureus, S. typhimurium and other pathogenic bacteria. The ODM was preferred over the AWD assay in characterization experiments as aggregation, non-diffusible bacteriocins, medium composition, protease inactivation and concentration may influence the sensitivity of the AWD assay (Lewus et al., 1991), leading to false negative results. Maximum bacteriocin activity was marked at the stationary growth phase, which suggests that the antimicrobial peptide is a secondary metabolite (Lisboa et al., 2006), a character confirmed by its low molecular weight (Abada, 2008).
 | |
| Fig. 6: | SDS-PAGE electrophoresis of the partially purified bacteriocin of B. megaterium 19. Lane a: Coomassie Brilliant blue-stained gel with small and large molecular weights of standard markers, lane b: Single band of partially-purified bacteriocin. Markers from top to bottom included: Bovine Serum albumin (E. coli), ovalbumin (chicken egg), carbonic anhydrase (bovine erythrocytes), ß-lactoglobilin (bovine milk), lyzozyme (chicken egg white), a-lactalbumin, aprotinin, insulin drain oxidized and bacitracin. Sizes on the left are indicated in kDa |
In this study, we reported the effect of several cultural conditions on the bacteriocin activity. Growth temperature played an important role and was often correlated with bacteriocin production (Todorov and Dicks, 2006). The activity of bacteriocin was much higher at 30°C than at 37°C, which was in good agreement with results by Bizani and Brandelli (2002). The high bacteriocin activity using MRS broth and low activity levels in BHI broth, molasses and whey suggests that specific nutrients are required for bacteriocin production (De Kwaadsteniet et al., 2005). Contradictory results to our findings were those reported by Lisboa et al. (2006) on bacteriocin produced by B. amyloliquefaciens, where maximal activity was evaluated at 37°C in BHI broth. The maximum values of bacteriocin activity were observed by preparations obtained from B. megaterium 19 grown in MRS broth at initial pH of 5.5, 5 and 6.5. This result was in consistence with those reported by Todorov et al. (2006) on L. lactis subsp. lactis bacHV219. Our bacteriocin was active under limited or reduced oxygen in the medium as indicated by Verluyten et al. (2003). Detectable inhibitory levels of the bacteriocin were observed in mMRS broth supplemented with sucrose, which may represent the first report of sucrose stimulation to bacteriocin production by a member of genus Bacillus. In contrast, most workers demonstrated high bacteriocin yields in association with the presence of glucose in the medium and not sucrose (Aasen et al., 2000).
Todorov et al. (2006) reported that yeast extract is the most effective organic nitrogen compound for bacHV219 production. This finding was not in accordance with our results, where beef extract was the nitrogen source that highly accelerated the production of B. megaterium 19 bacteriocin. The interesting feature of heat stability at 100°C for 15 min, supports the fact that it might constitute an advantage in view of its potential use as a food additive, in processes like pasteurization, drying, refrigeration and freezing. The thermal stability of our bacteriocin was markedly lost upon autoclaving, probably due to the lengthy time of exposure to heat and pressure. Accordingly, Vinod et al. (2006) described that the activity of the bacteriocin produced by Lactobacillus CA44 was completely lost at 121°C for 15 min. A similar result was reported for thuricin 7 produced by B. thuringiensis BMG1.7 (Cherif et al., 2001). Our pH results were consistent with those reported by Karaoglu et al. (2003), where the bacteriocin characterized showed an antimicrobial activity at the acidic pH more than the basic pH. Exposing the bacteriocin to prolonged periods of storage (up to 90 days) at 4°C resulted in an insignificant decrease in activity, which indicates that cold temperatures may be the appropriate preservation technique. B. megaterium 19 bacteriocin was stable after 15 min exposure to UV light and was completely destroyed after 90 min. These results were not surprising and confirmed the protein status of the bacteriocin, where the ultraviolet light may modify and/or change the ring structure of peptides, or affect the protein function (Ogunbanwo et al., 2003a). B. megaterium 19 bacteriocin was amylase and lipase sensitive, compared to other proteolytic enzymes (trypsin, pepsin and papain) used in this study, suggesting the presence of glycosylated and lipid moieties in the bacteriocin. The proteolytic enzymes failed to modify the antimicrobial activity of the bacteriocin, which is not unusual (Korenblum et al., 2005) and might be due to the presence of unusual amino acids in the bacteriocin structure, or cyclic N-and/ or C- terminally blocked peptides. Ethanol and hexane did not inhibit the activity of the bacteriocin, which might confirm the presence of lipid moieties in the bacteriocin structure. The bactericidal mode of action of our bacteriocin determined by the TEM may probably be due to pore formation (Montville et al., 1995) as typically reported for LAB bacteriocins (De Kwaadsteniet et al., 2006). The fraction containing the antimicrobial activity was analyzed and a single protein band with a range of molecular mass of about 3.496-6.512 kDa was identified. This result was in agreement with those obtained from the SDS-PAGE assays of some other bacteriocins produced by P. acidilactici and B. cereus 8/10 where molecular weights were estimated to range from 3.5-6.5 and 4-6 kDa, respectively (Albano et al., 2007). Svetoch et al. (2005) reported close results, where the characterized bacteriocin of B. circulans had an approximate molecular weight of 3.5 kDa.
Conclusion: The characterization of our bacteriocin revealed interesting properties that justifies it importance regarding food safety and may positively have an impact on its potential application in the biological control of pathogenic and spoilage microorganisms and hence food protection. The study showed evidence of the fact that the type of medium and medium components can influence bacteriocin activity, which may be taken into consideration when developing food products and modifying their consumption acceptability properties including texture and taste perception.
ACKNOWLEDGMENT
Mrs. Fatima Djadouni was the recipient of financial support provided by the Algerian government through in an exchange program between the University of Algeria and Alexandria University, Egypt.
REFERENCES
- Aasen, I.M., T. Moreto, T. Katla, L. Axelsson and I. Storro, 2000. Influence of complex nutrients, temperature and pH on bacteriocins production by Lactobacillus sake CCUG 42687. Applied Microbiol. Biotechnol., 53: 159-166.
CrossRefDirect Link - Abada, E.A., 2008. Isolation and characterization of a antimicrobial compound from Bacillus coagulans. Anim. Cells Syst., 12: 41-46.
Direct Link - Albano, H., S.D. Todorov, C.A. van Reenen, T. Hogg, L.M. Dicks and P. Teixeira, 2007. Characterization of 2 bacteriocins produced by Pediococcus acidilactici isolated from Alheira a fermented sausage traditionally produced in Portugal. Int. J. Food Microbiol., 116: 239-247.
Direct Link - Bizani, D. and A. Brandelli, 2002. Characterization of a bacteriocin produced by a newly isolated Bacillus sp. strain 8A. J. Applied Microbiol., 93: 512-519.
CrossRefPubMedDirect Link - Cherif, A., H. Ouzari, D. Daffonchio, H. Cherif and K.B. Slama et al., 2001. Thuricin 7: A novel bacteriocin produced by Bacillus thuringiensis BMG1.7, a new strain isolated from soil. Lett. Applied Microbiol., 32: 243-247.
CrossRefPubMedDirect Link - De Kwaadsteniet, M., S.D. Todorov, H. Knoetze and L.M. Diks, 2005. Characterization of a 3944 Da bacteriocin produced by Enterococcus mundtii ST15, with activity against gram-positive and gram-negative bacteria. Int. J. Food Microbiol., 105: 433-444.
CrossRefPubMedDirect Link - De Kwaadsteniet, M., T. Fraser, C.A. van Reenen and L.M.T. Dicks, 2006. Bacteriocin T8, a novel class IIa sec-dependent bacteriocin produced by Enterococcus faecium T8, isolated from vaginal secretions of children infected with human immunodeficiency virus. Applied Environ. Microbiol., 72: 4761-4766.
CrossRefDirect Link - Delmar, B., S. Amanda, J. Motta, R. Morrissy, A. Terra, A. Souto and A. Brandelli, 2005. Antibacterial activity of cerein 8A, a bacteriocin-like peptide produced by Bacillus cereus. Int. Microbiol., 8: 125-131.
PubMedDirect Link - Diop, M.B., R. Dubois-Dauphin, E. Tine, A. Ngom, J. Destain and P. Thonart, 2007. Bacteriocin producers from traditional food products. Biotechnol. Agron. Soc. Environ., 11: 275-281.
Direct Link - Karaoglu, A.S., F. Aydin, S.S. Kilic and A.O. Kilic, 2003. Antimicrobial activity and characteristics of bacteriocins produced by vaginal lactobacilli. Turk. J. Med. Sci., 33: 7-13.
Direct Link - Korenblum, E., I. von der Weid, A.L.S. Santos, A.S. Rosado and G.V. Sebastian et al., 2005. Production of antimicrobial substances by Bacillus subtilis LFE-1, B. firmus H2O-1 and B. licheniformis T6-5 isolated from an oil reservoir in Brazil. J. Applied Microbiol., 98: 667-675.
Direct Link - Lasta, S., Z. Fajloun, H. Darbon, P. Mansuelle and N. Andreotti et al., 2008. Chemical synthesis and characterization of J46 peptide, an atypical class IIa bacteriocin from Lactococcus lactis subsp. cremoris J46 strain. J. Antibiotics, 61: 89-93.
PubMed - Lewus, C.B., A. Kaiser and T.J. Montville, 1991. Characterization of bacteriocins produced by lactic acid bacteria isolated from meat. Applied Environ. Microbiol., 8: 1683-1688.
Direct Link - Line, J.E., E.A. Svetoch, B.V. Eruslanov, V.V. Perelygin and E.V. Mitsevich et al., 2008. Isolation and purification of enterocin E-760 with broad antimicrobial activity against gram-positive and gram-negative bacteria. Antimicrob. Agents Chemother., 52: 1094-1100.
CrossRef - Lisboa, M.P., D. Bonatto, D. Bizani, J.A. Henriques and A. Brandelli, 2006. Characterization of a bacteriocin-like substance produced by Bacillus amyloliquifaciens isolated from the Brazilian atlantic forest. Int. Microbiol., 9: 111-118.
PubMed - Maidak, B.L., J.R. Cole, C.T. Parker, G.M. Jr. Garitty and N. Larsen et al., 1999. A new version of the RDP ribosomal database project. Nucl. Acids Res., 27: 171-173.
PubMedDirect Link - Martin-Visscher, L.A., M.J. van Belkum, S. Garneau-Tsodikova, R.M. Whittal, J. Zheng, L.M. McMullen and L. Vederas, 2008. Isolation and characterization of carnocyclin A, a novel circular bacteriocin produced by Carnobacterium maltaromaticum UAL307. Applied Environ. Microbiol., 74: 4756-4763.
CrossRefDirect Link - Montville, T.J., K. Winkowski and R.D. Lusdescher, 1995. Models and mechanisms for bacteriocin action and application. Int. Dairy J., 5: 797-814.
CrossRef - Naclerio, G., E. Ricca, M. Sacco and M. de Felice, 1993. Antimicrobial activity of a newly identified bacteriocin of Bacillus cereus. Applied Environ. Microbiol., 59: 4313-4316.
PubMed - Ogunbanwo, S.T., A.I. Sanni and A.A. Onilude, 2003. Characterization of bacteriocin produced by Lactobacillus plantarum F1 and Lactobacillus brevis OG1. Afr. J. Biotechnol., 2: 219-227.
Direct Link - Ogunbanwo, S.T., A.I. Sanni and A.A. Onilude, 2003. Influence of cultural conditions on the production of bacteriocin by Lactobacillus brevis OG1. Afr. J. Biotechnol., 27: 179-184.
Direct Link - Pattnaik, P., J.K. Kaushik, S. Grover and V.K. Batish, 2001. Purification and characterization of a bacteriocin-like compound (lichenin) produced anaerobically by Bacillus licheniformes isolated from water buffalo. J. Applied Microbiol., 91: 636-645.
PubMed - Schillinger, U. and F.K. Lucke, 1987. Identification of lactobacilli from meat and meat product. Food Microbiol., 4: 199-208.
CrossRef - Svetoch, E.A., N.J. Stern, B.V. Eruslanov, Y.N. Kovalev and L.I. Volodina et al., 2005. Isolation of Bacillus circulans and Paenibacillus polymyxa strains inhibitory to Campylobacter jejuni and characterization of associated bacteriocins. J. Food Prot., 681: 11-17.
Direct Link - Brink, B.T., M. Minekus, J.M.B.M. van der Vossen, R.J. Leer and J.H.J.H. Intveld, 1994. Antimicrobial activity of lactobacilli: Preliminary characterization and optimization of production of acidocin B, a novel bacteriocin produced by Lactobacillus acidophilus M46. J. Applied Bacteriol., 77: 140-148.
CrossRefPubMedDirect Link - Todorov, S.D., S.T. Danova, C.A. van Reenen, M. Meincken, G. Dinkova, I.V. Ivanova and L.M. Dicks, 2006. Characterization of bacteriocin HV219, produced by Lactococcus lactis subsp. lactis HV219 isolated from human vaginal secretions. J. Basic Microbiol., 46: 226-238.
PubMedDirect Link - Verluyten, V., W. Messens and L. de Vuyst, 2003. The curing agent sodium nitrite, used in the production of fermented sausages, is less inhibiting to the bacteriocin-producing meat starter culture Lactobacillus curvatus LTH 1174 under anaerobic conditions. Applied Environ. Microbiol., 69: 3833-3839.
Direct Link - Vinderola, C.G., G.A. Costa, S. Regenhardt and J.A. Reinheimer, 2002. Influence of compounds associated with fermented dairy products on the growth of lactic acid starter and probiotic bacteria. Int. Dairy J., 12: 579-589.
CrossRef - Vinderola, C.G., P. Mocchiutti and J.A. Reinheimer, 2002. Interactions among lactic acid starter and probiotic bacteria used for fermented dairy products. J. Dairy Sci., 85: 721-729.
PubMed - Joshi, V.K., S. Sharma and N.S Rana, 2006. Production, purification, stability and efficacy of bacteriocin from isolates of natural lactic acid fermentation of vegetables. Food Technol. Biotechnol., 44: 435-439.
Direct Link - Lyon, W.J. and B.A. Glatz, 1991. Partial purification and characterization of a bacteriocin produced by Propionibacterium thoenii. Applied Environ. Microbiol., 57: 701-706.
PubMed - Aktypis, A., G. Kalantzopulous, J.H.J. Huis in't Veld and B. ten Brink, 1998. Purification and characterization of thermophilin T, a novel bacterium produced by Streptococcus thermophilus ACA-DC 0040. J. Applied Microbiol., 84: 568-576.
CrossRefDirect Link