
C. A. Echendu
Department of Human Nutrition and Dietetics, Michael Okpara University of Agriculture, Umudike, PMB 7267, Umuahia, Abia State, Nigeria
I. C. Obizoba
Department of Home Science, Nutrition and Dietetics, University of Nigeria, Nsukka, Nigeria
J. U. Anyika
Department of Human Nutrition and Dietetics, Michael Okpara University of Agriculture, Umudike, PMB 7267, Umuahia, Abia State, Nigeria
Pakistan Journal of Nutrition
Year: 2009 | Volume: 8 | Issue: 12 | Page No.: 1849-1854







ABSTRACT
This study evaluated the effects of germination on the chemical composition of groundbean seeds. The seeds were cleaned, soaked for 8h and germinated for 0-4 days. The samples were separately dried and milled to fine flours. The flour samples were analyzed for proximate, mineral and antinutrient composition using standard methods. Results showed that groundbean germinated for 72h (GGB72) had the highest proximate composition which was significantly different from the rest (p<0.05) except for groundbean germinated for 96h (GGB96) in fibre (1.22%) and CHO (67.40%) in groundbean germinated for 0h (ungerminated). Groundbean germinated for 48h (GGB48) proved to be more beneficial in most mineral content than the rest of the germination periods (p<0.05). Antinutrient content was highest in groundbean germinated for 48h (GGB48) except for tannin. The study showed that 72h of germination of groundbean (GGB72) proved to be the best for highest nutrient content while germination for 48h increased antinutrient content.
PDF Abstract XML References Citation
How to cite this article
C. A. Echendu, I. C. Obizoba and J. U. Anyika, 2009. Effects of Germination on Chemical Composition of Groundbean (Kerstingiella geocarpa harm) Seeds. Pakistan Journal of Nutrition, 8: 1849-1854.
DOI: 10.3923/pjn.2009.1849.1854
URL: https://scialert.net/abstract/?doi=pjn.2009.1849.1854
DOI: 10.3923/pjn.2009.1849.1854
URL: https://scialert.net/abstract/?doi=pjn.2009.1849.1854
INTRODUCTION
Groundbean (Kerstingiella geocarpa harm) is a lesser known and under-exploited grain legume crop. It is said to have originated in the savanna areas of West Africa and has a very restricted range of cultivation being confined to tropical Africa, particularly Nigeria, Mali, Burkina-Faso, Upper Volta, Niger, Benin and Togo (Kay, 1979; Obasi and Agbatse, 2003) where it is grown at subsistence level.
Groundbean is widely cultivated in both high rainfall and savanna areas in tropical Africa (NAS, 1979). It is grown successfully in areas with annual rainfall of between 500-600 mm. It grows satisfactorily on many types of soil and most types of tropical and sub-tropical climate. The crop reaches maturity 90-150 days after planting. There are many different forms of Kerstingiella groundnut although widely reported to be found in cultivated states. Hepper (1963) differentiated between a robust cultivated form, Kerstingiella geocarpa var geocarpa and a wild form Kerstingiella geocarpa var tisseranti. The Kerstingiella groundnut is frequently confused with the bambara groundnut, Voandzeia subterranea, but can be distinguished by its flowers which have a deeply divided calyx with narrow lobes.
Germination is one of the methods used in elimination of various anti-nutritional factors present in foods. It is a natural process in which dormant but viable seeds are induced to start growing into seedlings. This is the process by which amylase degrade starches into dextrin and maltose. Germination of seed is a simple process that does not require sunlight or soil and requires only short sprouting time. However, its yield is high (Chen, 1970). It is characterized by a changing array of enzymatic activities. Some of these activities increase dramatically from an initially low or even undetected state to a moderate level. Later these activities then decline to an activity lower than initially present. Another group of enzyme activities are found in dry seed at an intermediate level which remain constant throughout germination (Nnanna and Phillips, 1988). The enzymes convert the stored foods such as insoluble carbohydrates and proteins to soluble components (Enwere, 1998). Nout and Ngoddy (1997) reported that germination of seeds triggers the enzyme systems that cause breakdown of complex macromolecules of proteins, carbohydrates and lipids into simpler forms that are much easily assimilated. Proteins are broken down to peptides and amino acids by protease activity. Nitrogen is transferred to developing axis, carbohydrate to simpler sugars by amylase, phytic acid to inositol and phosphoric by phytase and breakdown of tannin-protein-enzyme-mineral complexes.
Sprouting seeds of most cereals and legumes have shown improvements in nutrients in human diet and compare well with their fresh counterparts if not better (Kakade and Evans, 1966; Kylen and McCready, 1975).
Studies have shown that although legumes are known for their high protein content, their utility is limited because of the low protein digestibility. A combination of sprouting and cooking resulted in an excellent digestibility coefficient (El-Hag et al., 1978; Ologhobo and Fetuga, 1986).
Germination resulted in greater retention of all minerals and B-complex vitamins compared to cooking treatment in chickpeas (El-Adawy, 2002). Germination increased amount of thiamin, riboflavin, niacin and ascorbic acid in both soybean and mung bean (Abdullah and Baldwin, 1984). Mineral values in sprouted beans increased with germination except with iron. Iron values decreased in sprouted seeds but its availability increased due to an increase in phytase activity during seed germination (Bates et al., 1977; Walker and Kochhar, 1982).
As germination takes place, the anti-nutritional factors are greatly decreased to insignificant levels or to nothing (Siddhuraju and Becker, 2001; El-Adawy, 2002; Ugwu and Oranye, 2006). The objective of this study therefore is to determine the effects of germination on the chemical composition of groundbean.
MATERIALS AND METHODS
Sources of materials: Groundbean (GB) is not widely cultivated in Nigeria as well as in Enugu State. It was bought (10 kg) from Nrobo market in Uzo-Uwani L.G.A. of Enugu State where it is commonly grown.
Preparation and treatment of samples: Groundbean seeds were sorted and cleaned to remove spoilt grains, stone and other foreign materials.
Pilot study (germination of GB): Cleaned GB seeds were soaked for 2, 4, 6, 8 and 10h in order to ascertain appropriate soaking period that would enhance dehulling and germination within 24h. Soaking for 8h was found to be most appropriate for both treatments.
Germinated GB: The 8h soaked seeds were separately spread on four wet jute bags, covered with muslin cloth and wet jute bags. Water was sprinkled on the seeds every 12 h until the end of germination periods (24, 48, 72 and 96h). The germinated seeds were picked carefully with the sprouts, washed, dehulled, oven dried at 50oC for 24h, milled and stored in name labelled polyethylene bags prior to analysis.
Data analysis: Data were analyzed using the computer programme statistical software package (SAS, 2003). Analysis of variance (ANOVA), Standard Error of the Mean (SEM) and Least Significant Difference (LSD) were used to separate the mean differences among samples (p<0.05).
RESULTS AND DISCUSSION
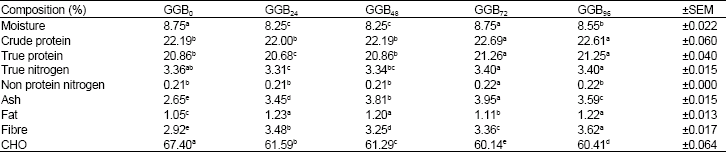
Table 1 presents proximate composition of ungerminated and germinated groundbean. The moisture content ranged from 8.25 in 24h to 8.75% in ungerminated (control) and 72h germination. Generally, the moisture content were comparable regardless of treatment (p>0.05).
The Crude Protein (CP) values ranged from 22.00 in 24h to 22.69% in 72h germination. Germination significantly caused increased CP values. The 72 and 96h germination periods had significantly higher values (22.69 and 22.61%) than others (p<0.05). The values for the control and 48h germination periods were similar (22.19%). The true protein values followed almost the same trend as in CP values and ranged from 20.68% in 24h to 21.26% in 72h germination. The GB 72h germination that had the highest CP also had the highest TP (22.69 and 21.26%). The True Protein Nitrogen (TPN) ranged from 3.31% in 24h to 3.40% in 72 and 96h. 24h TPN value was significantly lower than others (p<0.05). Germination increased the Non Protein Nitrogen (NPN) from 0.21% in the control to 0.22% in 96h germination. The NPN of the 72 and 96h germinated GB were higher (0.22%) as compared with those of the 24, 48h and the control (0.22 vs 0.21%).
The ash values for ungerminated GB increased with increase in germination period. The range was from 2.65% in the control to 3.95% in 72h germination. The 72h sample which had the highest ash differed from others (p<0.05).
Generally, both the control and the treated samples had low fat and the range was from 1.05% in the control to 1.23% in 24h germinated sample. Germination increased fat levels. The 24h sample had comparable value with those of 48 and 96h (p>0.05). The 72h had the least increase in fat.
Germination had varied fibre increases in GB. The range was from 2.92-3.62%. The 96h and 24h germination periods had the highest (3.62 and 3.48%, respectively). On the other hand, the 72h sample was slightly higher than that of 48h (3.36 vs 3.25%). Germination decreased carbohydrate. The range was from 67.4 in the control to 60.14% in 72h germination.
The untreated and the 72h germinated sample had comparable moisture content (8.75%) (Table 1). Generally, one can say that GB has comparable moisture regardless of treatment. This could be due to the effectiveness of the drying method used for the samples before moisture was determined. This is in contrast with the observation made by Obizoba (1991) that moisture level increased as sprouting times increased in pigeon pea. Low moisture content enhances keeping quality of flours (Echendu, 2004).
Germination has been shown to increase protein content of legumes (Kylen and McCready, 1975; Fordham et al., 1975). Germination increased protein content from 22.00 in 24h to 22.69% in 72h germination. These values are lower than 22.38-24.82% indicated previously (Chikwendu, 2003). The increases in crude protein, TP and TPN could be due to enzyme hydrolysis of the insoluble protein to soluble protein, which increased the protein availability. The increases which peaked at the third day of germination agreed with other works (Akpapunam and Achinewhu, 1985; Chikwendu, 2003) for cowpea (72h) and groundbean (48h), respectively. It has been reported that increase in protein during germination was due to the release of free amino acids after enzymatic hydrolysis for the synthesis of new protein (Bliss, 1975).
| Table 1: | Proximate composition of ungerminated and germinated groundbean |
 | |
Values are means of triplicate samples ±SEM; Means bearing different superscripts in the same column differed (p<0.05) | |
| GGB0 = 0h Germinated Groundbean GGB72 = 72h Germinated Groundbean | GGB24 = 24h Germinated Groundbean GGB96 = 96h Germinated Groundbean | GGB48 = 48h Germinated Groundbean |
The higher crude protein and lower true protein observed in all the samples was not a surprise. Crude protein contains more non protein N which reduces digestion, absorption and utilization. Lower TP value is much more digested, absorbed and utilized in both animals and humans (Ene-Obong and Obizoba, 1995). The similarity in non protein N for both 72 and 96h germinated flours (0.22%) or no difference between those of 24, 48h as well as the control (0.21%) led to high TPN which significantly increased the TP of the samples. The increase was mainly due to the increased activity of proteolytic enzymes which break down the protein molecules into simple units of amino acids as observed in pigeon pea (Obizoba, 1991) and in fenugreek seeds (El-Mahdy and El-Sebaily, 1982).
The ash values (Table 3) are lower than those reported for pigeon pea (Obizoba, 1991) and comparable to those reported for GB earlier (Chikwendu, 2003). The increases in ash from 24-96h germination period indicate that germination particularly 72h period is a good method of increasing ash content of GB. The increase in ash content was due to endogenous enzyme hydrolysis of complex organic compounds to release more nutrients leaving the antinutrients to leach into the germination medium (Chikwendu, 2003). The values for fat were generally low although germination caused slight increases. This is in contrast with the report of Obizoba (1991) where fat values for sprouted and unsprouted pigeon pea remained the same. The low fat levels for legume grains is because its major function is to maintain the integrity of cell wall and other forms of protection (Chikwendu, 2003). The slight increase might be due to non-conversion of free fatty acids to carbohydrates which may lead to increase in fat composition during germination (Afam-Anene and Onuoha, 2006).
The fibre values were higher than those previously reported for GB (Chikwendu, 2003). The increases in fibre due to germination might be that the microflora enzymes hydrolyzed complex carbohydrate to release fibre which subsequently decreased carbohydrate. The decrease in carbohydrate due to germination might also be that the young seedlings might have used it for metabolism (Obizoba and Atu, 1993). Another possible reason might be due to leaching into germination medium. Obizoba and Egbuna (1992) observed a similar decrease due to both germination and fermentation.
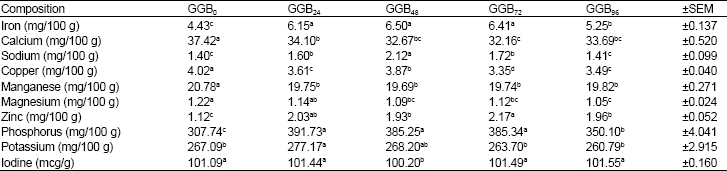
Table 2 shows the mineral composition of ungerminated and germinated groundbean. Germination significantly increased iron (Fe) in GB. However, the 96h germination had the least value (5.25 mg). The peak of increase was at 48h (6.50 mg). The control had significantly least value (4.43 mg) (p<0.05). Germination significantly decreased Ca values. The range was from 32.16 mg in 72h germination period to 37.42 mg in the control. The 24h germination had the highest (34.10 mg) as compared with the other periods (32.16-33.69 mg). However, the differences were insignificant (p>0.05). Germination had varied effects on Na. It increased it from 1.40-2.12 mg. The highest increase occurred at 48h followed by 72h (2.12 and 1.72 mg). The 96h sample had only 0.01 mg higher than that of the control (1.40 mg).
Copper values differed and the control had the highest (4.02 mg). Germination caused various decreases among the treatments. The highest decrease occurred at the 72h (3.35 mg) followed by that of 96h (3.49 mg). On the other hand, the 48h sample had a value (3.87 mg) which was higher than those of other treated samples. Manganese values followed the same trend as copper and ranged from 19.69 mg in 48h to 19.82 mg in 96h germination. The control had the highest value (20.78 mg), while the 96h sample was highest among the treatments (19.82 vs 19.75, 19.69 and 19.74 mg). Magnesium values were generally low for both the control and the treated samples. The control had 1.22 mg. The range for the treated samples was from 1.05 mg in 96h to 1.14 mg in 24h germination. Even though there were slight differences among the samples, the differences were not significant (p>0.05).
| Table 2: | Mineral composition of ungerminated and germinated groundbean |
 | |
Values are means of triplicate samples ±SEM; Means bearing different superscripts in the same column differed (p<0.05) | |
| GGB0 = 0h Germinated Groundbean GGB72 = 72h Germinated Groundbean | GGB24 = 24h Germinated Groundbean GGB96 = 96h Germinated Groundbean | GGB48 = 48h Germinated Groundbean |
Zinc values differed and ranged from 1.12-2.17 mg. The control had the least (1.12 mg). On the other hand, 24 and 72h samples had the highest and comparable values (2.03 and 2.17 mg). The 48 and 96h samples also had similar values (1.93 and 1.96 mg, respectively) (p>0.05). Phosphorus values varied for both germinated and ungerminated samples. The control had the least (307.74 mg). The values for the treated samples ranged from 350.10 in 96h to 391.73 mg in 24h germinated sample.
Germination had varied effects on potassium. The values ranged from 260.79 in 96h to 277.17 mg in 24h period. The 24 and 48h germination increased K from 267.09-277.17 and 268.20 mg, respectively. On the other hand, prolonging germination from 72-96h caused more decreases as compared with the control, 24 and 48h samples (263.70 and 260.79 vs 267.09, 277.17 and 268.20 mg, respectively). Iodine values differed and ranged from 101.09 in the control to 101.55 mcg in 96h germination. The 48h germination caused much more decrease in 12 when compared with control as well as other treatments (100.20 vs 101.09, 101.44, 101.49 and 101.55 mcg, respectively).
The similarity in Fe for the 24, 48 and 72h germination periods appears to show that GB could be germinated for any of the periods to improve Fe content against the control (4.43 mg) (Table 2). The increase might be that the hydrolytic enzymes released more free Fe from its organic complexes (Obizoba and Atii, 1994). Germination had adverse effect on Ca. The lower values for all the treated samples might be due to leaching, dehulling or the seedlings used it for metabolism (Obizoba and Atii, 1994). This agrees with the report of Chikwendu (2003) but contrary to that of Ganesh Kumar et al. (1978) in chick pea, green gram and cowpea. The slightly low value for Na indicates that GB may not necessarily be a rich source of Na. The lower Na content of GB qualifies it to be one of the foods for patients on restricted Na intake. However, germination increased Na content up to 72h period. This agrees with the work of Abdullah and Baldwin (1984) for soya bean and mungbeans. The decreases in Cu due to treatment might be that the young embryo used Cu for metabolism which is a commonly observed phenomenon (Chikwendu, 2003).
The higher Mn for the control might not be bioavailable as compared with the germinated samples because the Mn may not be released from the organic complexes. The comparable values for all the treated samples (Table 2) suggest that germination of GB for any of the periods would produce GB flour rich in Mn. The low Mg value for both the control and the germinated samples might indicate that GB is not a good source of Mg. The lower value for the 96h sample (Table 2) indicates that it is not beneficial to germinate GB up to 96h to improve Mg nutriture. The slightly higher values for 24 and 72h samples indicate that these two periods had an edge over the other periods with respect to Zn content of GB. The increase in Zn due to germination is of interest. Zinc is recently recognized as the fourth world-wide deficient micronutrient. The increases might be that during germination, the increases in various enzymes hydrolyzed bonds between mineral-protein enzyme complexes to release free Zn (Obizoba and Amaechi, 1992).
The 24h sample had higher P value than 48 and 72h samples (391.73 vs 385.25 and 385.34 mg). The high P for the 24h sample though comparable with 48 and 72h samples appears to be the optimum period to germinate GB to obtain high P. It is known that during germination, the dormant phytase is activated to hydrolyze bonds between phytin protein-enzyme to release more free phosphorus. This is in agreement with another work (Obizoba and Amaechi, 1992). The decreases in K during 72 and 96h germination period might be due to increased need for nutrients by the growing plant metabolism (Table 2) (Obizoba and Atii, 1994). The slightly higher 12 value for 96h sample appears to suggest that it is the optimal period to germinate GB to increase its 1 2 content. The increase at 96h might be due to increased hydrolytic action of micro flora enzymes (Anyika, 2003).
Table 3 depicts the antinutrient composition of ungerminated and germinated groundbean. Tannins values were low for all samples regardless of treatments and ranged from 0.10-0.14 mg.
| Table 3: | Antinutrient composition of ungerminated and germinated groundbean |
 | |
Values are means of triplicate samples ±SEM; Means bearing different superscripts in the same column differed (p<0.05) | |
| GGB0 = 0h Germinated Groundbean GGB72 = 72h Germinated Groundbean | GGB24 = 24h Germinated Groundbean GGB96 = 96h Germinated Groundbean | GGB48 = 48h Germinated Groundbean |
The 24h sample had the highest decrease (0.10 mg) (p<0.05). The 48h sample had similar value with that of the control (0.14 mg). Phytate values ranged from 1.19 in 96h germination period to 1.44 mg in 48h period. The 24, 48 and 72h samples had higher and comparable values (1.42, 1.44 and 1.38 mg, respectively) than 96h germination. Germination increased phytate levels with the exception of 96h period when compared with the control (1.27 vs 1.19 mg).
Germination had varied effects on trypsin inhibitor. The 72 and 96h germination periods decreased it when compared with the control (2.22 vs 1.38 and 1.19 mg, respectively). The 24h sample had significantly higher TI value (p<0.05). Oxalate values followed the same trend as that of TI and ranged from 0.12-0.20 mg. The 24 and 48h samples had higher and comparable values (p>0.05). These values (19 and 20 mg) differed from that of the control (15 mg) as well as those of the 72 and 96h (0.14 and 0.12 mg, respectively). Cyanide values differed and ranged from 0.04-0.08 mg. The 48h germination increased cyanide most (0.08 mg). The 24 and 96h germination slightly increased cyanide when compared with the control and 72h germination values (0.05 and 0.06 mg vs 0.04 mg). These increases were not significantly different (p>0.05).
Tannins values were low but 48h germination appeared to have no edge over the control. The increase in phytate due to germination except for the 96h sample was not a surprise. Many workers have observed increases in phytate in germinated or fermented green leafy vegetables (Udofia, 2005; Mefoh, 2008). The increase might be due to hydrolysis of some phytate related compounds which could have some beneficial effects as anti oxidants (Graf and Eaton, 1993). Again, 96h might be the optimum period for high phytase activity during GB germination (Igbedioh et al., 1994; El-Adawy, 2002). The higher T.I. value for the 24h sample (4.55 mg) suggests that the higher the germination period, the lower the TI concentration (Table 3). The values for 72 and 96h periods were 1.38 and 1.19 mg, respectively. These are higher than those reported elsewhere (Chikwendu, 2003). Trypsin inhibitor activities affect protein digestibility negatively. Oxalate levels decreased as germination period advanced. It is interesting to know that oxalate was higher during the 24 and 48h periods (0.19 and 0.20 mg). The lower oxalate (0.11 mg) for the 96h sample strongly suggests that the optimum period to reduce oxalate in GB is 96h. The lower cyanide values (0.04-0.08 mg) in GB strongly suggest that GB contains low cyanide. This also suggests that people that depend on GB as their source of nutrient have less risk of ill health consequences based on cyanide. Generally, it seems that germination for longer periods (72 and 96h) were more effective in reducing antinutrients.
Conclusion: The study showed that 72h of Germination of Groundbean (GGB72) proved to be the best for highest nutrient content while germination for 48h increased antinutrient content.
REFERENCES
- Fordham, J.R., C.C. Wells and L.H. Chen, 1975. Sprouting of seeds and nutrient composition of seeds and sprouts. J. Food Sci., 40: 552-556.
Direct Link - Kumar, K.G., L.V. Venkataraman, T.U. Jaya and K.S. Krishnamurthy, 1978. Cooking characteristics of some germinated legumes. Changes in phytins, Ca++, Mg++ and pectins. J. Food Sci., 43: 85-88.
CrossRefDirect Link - Graf, E. and J.W. Eaton, 1993. Suppression of colonic cancer by dietary phytic acid. Nutr. Cancer, 19: 11-19.
Direct Link - Igbedioh, S.O., K.T. Olugbemi and M.A. Akpapunam, 1994. Effects of processing methods on phytic acid level and some constituents in bambara groundnut (Vigna subterranea) and pigeon pea (Cajanus cajan). Food Chem., 50: 147-151.
CrossRef - Kylen, A.M. and R.M. McCready, 1975. Nutrients in seeds and sprouts of alfalfa, lentils, mung beans and soybeans. J. Food Sci., 40: 1008-1009.
Direct Link - Nout, M.J.R. and P.O. Ngoddy, 1997. Technological aspects of preparing affordable fermented complementary foods. Food Control, 8: 279-287.
CrossRefDirect Link - Obasi, M.O. and A. Agbatse, 2003. Evaluation of nutritive value and some functional properties of Kerstings groundnut seeds for optimum utilization as a food and feed resource. E. Afr. Agric. For. J., 68: 173-181.
Direct Link - Obizoba, I.C. and N.A. Amaechi, 1993. The effect of processing methods on the chemical composition of baobab (Adansonia digitata L) pulp and seed. Ecol. Food Nutr., 29: 199-205.
CrossRefDirect Link - Obizoba, C.I. and V.J. Atii, 1994. Evaluation of the effect of processing techniques on the nutrient and antinutrient contents of pearl millet (Pennisetum glaucum) seeds. Plant Food Human Nutr., 45: 23-34.
CrossRefDirect Link - Obizoba, I.C. and H.I. Egbuna, 1992. Effect of germination and fermentation on the nutritional quality of bambara nut (Voandzeia subterranea L. Thouars) and its product (milk). Plants Food Hum. Nutr., 42: 13-23.
CrossRefDirect Link - Ugwu, F.M. and N.A. Oranye, 2006. Effects of some processing methods on the toxic components of African breadfruit (Treculia africana). Afr. J. Biotechnol., 5: 2329-2333.
Direct Link