
E. E. Iyare
Department of Physiology, College of Medicine, University of Nigeria, Enugu Campus, Enugu State, Nigeria
U. I. Nwagha
Department of Physiology/Obstetrics and Gynaecology, College of Medicine, University of Nigeria, Enugu Campus, Enugu State, Nigeria
Pakistan Journal of Nutrition
Year: 2009 | Volume: 8 | Issue: 11 | Page No.: 1760-1765







ABSTRACT
This study investigated whether consumption of aqueous extract of Hibiscus sabdariffa during the juvenile-pubertal period will affect the BMI and visceral organ weights at onset of puberty in rats. Weaned 21 days old rats, whose mothers were given food and water ad libitum during pregnancy and lactation, were divided randomly into three groups of nine rats each. Control group had tap water only while the extract groups had 0.6 g-extract/100 ml and 1.8 g-extract/100 ml as their drinking solution throughout the juvenile-pubertal period (PND 21-puberty onset). All groups had normal rat chow and their respective drinking solutions ad libitum. From PND 30 onwards, the rats were inspected daily for vaginal opening, which was used as the index for puberty onset. Rats in the extract groups (0.6 g/100 ml and 1.8 g/100 ml) drank less fluid and consumed less food compared with the control group at all periods of measurement. The weight of 0.6 g/100 ml was not different from the control whereas 1.8 g/100 ml was lower at PND 28, similar at PND 35 and higher at PND 42 and at puberty onset compared with the control. Puberty onset in the extract groups was delayed and body weights and lengths, BMI, visceral organs weights and weight indices were higher in the extract groups. It is concluded that consumption of aqueous extract of Hibiscus sabdariffa during the juvenile-pubertal period decreased fluid and food consumption, increased BMI and visceral organs’ weights and may predispose to obesity at puberty onset in rats.
PDF Abstract XML References Citation
How to cite this article
E. E. Iyare and U. I. Nwagha, 2009. Postweaning Consumption of Aqueous Extract of Hibiscus sabdariffa May Predispose Rats to Obesity. Pakistan Journal of Nutrition, 8: 1760-1765.
DOI: 10.3923/pjn.2009.1760.1765
URL: https://scialert.net/abstract/?doi=pjn.2009.1760.1765
DOI: 10.3923/pjn.2009.1760.1765
URL: https://scialert.net/abstract/?doi=pjn.2009.1760.1765
INTRODUCTION
It is well established that an animal’s reserves of adipose tissue can influence reproductive capability in two ways: by timing the onset of puberty and maintaining reproductive function during adulthood. Schneider and Wade (1989) demonstrated that fasting induced anoestrus occurs more rapidly in lean hamsters than in fat hamsters and rats maintained on a high fat diet was observed to come into first estrus earlier than do animals fed on a low fat diet (Frisch et al., 1975).
The metabolic pathway linking body fat stores with the neuroendocrine reproductive system was unclear until the discovery of the adipocyte-derived hormone, leptin. The finding that leptin circulates in plasma in proportion with body adiposity (Maffei et al., 1995) led to the theory that leptin acts as an “adipostat”, a humoral signal carrying information regarding energy reserves. The possibility of a link between leptin and reproduction became apparent when it was observed that a homozygous mutation in the leptin (ob) gene was responsible for the obesity syndrome in the obese (ob/ob) mouse (Zhang et al., 1994).
Several papers have testified to the reproductive shortcomings of ob/ob mice after the first published work on this mutant (Ingalls et al., 1950). These mice are infertile (Ingalls et al., 1950) and the infertility can be corrected in both sexes by the administration of leptin (Chehab et al., 1996; Mounzih et al., 1997), which stimulates the secretion of gonadotrophins and gonadal function (Barash et al., 1996). Treatment with recombinant leptin also advances puberty in normal female mice (Chehab et al., 1997) and rats (Cheung et al., 1997) and allows the onset of puberty in severely food-restricted rats (Gruaz et al., 1998).
It has been suggested therefore, that leptin is the signal that informs the brain that metabolic stores are adequate for the initiation of reproductive function (Chehab et al., 1997) and triggers puberty onset (Bronson and Manning, 1991; Campfield et al., 1996; Chehab et al., 1997) by interacting with the GnRH neuron (Lee, 1995) and causing changes in the frequency and magnitude of GnRH pulses which has been shown to herald the onset of puberty (Terasawa, 1995).
Hibiscus sabdariffa (HS; Family: Malvaceae) is an annual, erect, bushy, herbaceous sub-shrub that grows up to 8 ft (2.4 m) tall, with smooth or nearly smooth, cylindrical and typically red stems. Other botanical features are well described by Ross (2003). A beverage produced from the thick, red and fleshy, cup-shaped calyces of the flower are consumed worldwide. Extracts are also used in folk medicine in the treatment of a variety of ailments including high blood pressure, liver diseases and fever (Dalziel, 1973; Wang et al., 2000; Ross, 2003). The red anthocyanin pigments in the calyces are also employed in food colouring (Esselen and Sammy, 1975).
We had earlier shown that maternal consumption of aqueous extract of HS during pregnancy and lactation increases weight gain and BMI at onset of puberty in female offsprings. It is not known whether postweaning exposure to aqueous extract of HS will increase body weight and BMI at onset of puberty. The present study was thus designed to investigate this and also to examine the possible mechanism underlying any effect observed.
MATERIALS AND METHODS
Animals and treatment: Eighteen in-bred virgin female Sprague-Dawley (SD) rats age between 10-12 weeks and weighing 125±5.5 g (mean±SEM) with two consecutive regular 4-day estrous cycles were used for this study. They were housed individually in cages under standard environmental conditions. The estrous cycles were monitored and males of proven fertility were introduced into the cages of the female rats that were expected to get into the estrus phase within 12 h to allow for mating. Day 1 of pregnancy was taken as the day spermatozoa were seen in the vaginal smear of the rats. From day 1 of pregnancy through postnatal day 20 (PND 20), animals were given food and water ad libitum. On PND 21 the young rats were weaned and divided randomly into three groups of nine rats each (A, B and C). Group A had tap water only while groups B and C had 0.6 g/100 ml and 1.8 g/100 ml HS respectively as their drinking solution throughout the juvenile-pubertal period (PND 21-puberty onset). All groups had normal rat chow and their respective drinking solutions ad libitum. From PND 30 onwards, the rats were inspected daily for vaginal opening, which was used as the index for puberty onset (Engelbregt et al., 2001). The age at vaginal opening (onset of puberty) was recorded. The body weight (g) and length (cm) were measured for the calculation of Body Mass Index (BMI). Thereafter, the animals were sacrificed by inhalation of chloroform and the uteri and ovaries, kidney, heart, liver, spleen and the adrenal glands were removed, trimmed of fat and weighed.
Extraction procedure: Mature dry dark-red calyces of HS were purchased in a local market in Enugu, Nigeria. It was authenticated by a resource staff at the Department of Botany, University of Nigeria, Nsukka, Nigeria. The extraction procedure used in our laboratory was as described previously (Iyare and Iyare, 2006a,b). Briefly, 30 g of the dry petals of HS was brewed in 400 ml of boiled tap water for 45 min. The resulting decoction was filtered.
The concentrations in the extract groups (0.6 g/100 ml and 1.8 g/100 ml) were derived as follows: 10 mls of filtrate was added to 48 mls of tap water to make approximately 0.6 g/100 ml tap water while 10 mls was added to 9 mls of tap water to make approximately 1.8 g/100 ml tap water.
Statistical analysis: For data comparison between the three groups, the one way analysis of variance (ANOVA) was used followed by a post-hoc Students Newman Keuls test. p<0.05 was taken as statistically significant.
RESULTS
Fluid and food intake: Rats in the HS groups (0.6 g/100 ml and 1.8 g/100 ml) drank less fluid compared with the control group at PND 28 (p<0.05 and p<0.005 respectively) and 35 (p<0.05 for both) (Table 1). At PND 42, the fluid intake in the HS groups was not significantly different from that of the control (p>0.05 for both HS groups). There was no difference in fluid intake between the two HS groups at PND 28, 35 and 42 (p>0.05 at each period of measurement).
The food intakes by the rats in the HS groups (0.6 g/100 ml and 1.8 g/100 ml) were significantly lower than that of the control dams at all periods of measurement (p<0.05 and p<0.00005 respectively at PND 28, p<0.05 and p<0.01 respectively at PND 35, p<0.05 for both at PND 42) (Table 1). There was no difference in food intake between the two HS groups except at PND 28 when the food intake in the high dose HS group (1.8 g/100 ml) was significantly lower than that of the low dose HS group (0.6 g/100 ml) (p<0.05).
Postnatal weight: There was no significant difference in the weights of the rats at PND 0 (birth) and PND 21 (weaning) (Table 2). This was not surprising because these rats were still being nursed by their mothers that were given ad libitum food and water. There was no difference in postnatal weights between the low dose HS group (0.6 g/100 ml) and the control rats at PND 28, 35 and 42. The postnatal weight of the rats in the high dose HS group (1.8 g/100 ml) at PND 28 was significantly lower than that of the rats in both the control and the low dose HS groups (0.6 g/100 ml) (p<0.05), whereas at PND 35, it was not significantly different from both (p>0.05). At PND 42, the weight of the rats in the high dose HS group (1.8 g/100 ml) was significantly higher than those of the control and low dose HS groups (0.6 g/100 ml) (p<0.05).
Body parameters at vagina opening: The age at vagina opening in rats in the low dose HS group (0.6 g/100 ml) was significantly higher than those of the high dose HS (1.8 g/100 ml) (p<0.05) and control groups (p<0.0005) (Table 3). The age at vagina opening in rats in the high dose HS group (1.8 g/100 ml) was also significantly higher than that of the control rats (p<0.05).
| Table 1: | Effect of postweaning consumption of aqueous extract of Hibiscus sabdariffa on fluid and food intake |
 | |
N = 9 each. Values are expressed as M±SEM, * = p<0.05 compared with Control, P = p<0.05 compared with 1.8 g/100 ml | |
| Table 2: | Effect of consumption of aqueous extract of Hibiscus sabdariffa during the postweaning period on absolute body weight |
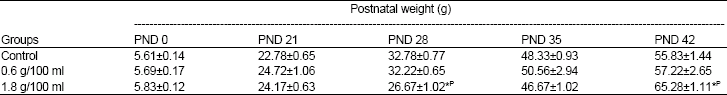 | |
N = 9 each. Values are expressed as M±SEM, * = p<0.05 compared with Control, P = p<0.05 compared with 0.6 g/100 ml | |
| Table 3: | Effect of consumption of aqueous extract of Hibiscus sabdariffa during the postweaning period on some body parameters at onset of puberty |
 | |
N = 9 each. Values are expressed as M±SEM, * = p<0.05 compared with Control, P = p<0.05 compared with 0.6 g/100 ml | |
The body weight at vagina opening in rats in the low dose HS group (0.6 g/100 ml) was also significantly higher than those of the high dose HS (1.8 g/100 ml) (p<0.01) and control groups (p<0.0005) (Table 3). The age at vagina opening in rats in the high dose HS group (1.8 g/100 ml) was also significantly higher than that of the control rats (p<0.01).
The body lengths of the rats in both groups of HS were significantly higher than that of the control rats (p<0.05) (Table 3). There was no statistical difference between the two HS groups (p>0.05).
The Body Mass Index (BMI) of the low dose HS group (0.6 g/100 ml) and the high dose HS (1.8 g/100 ml) HS group were significantly higher than that of the control group (p<0.01 and p<0.05 respectively) (Table 3). The BMI of the low dose HS group (0.6 g/100 ml) was also significantly higher than that of the high dose HS (1.8 g/100 ml) (p<0.05) group.
Reproductive organ weights: The weights of the ovaries in the rats in the two HS groups (0.6 g/100 ml and 1.8 g/100 ml) were significantly higher than that of the control (p<0.05 respectively) whereas there was no difference between the weights of the uteri in the two HS groups compared with the control (p>0.05) (Table 4). There was also no significant differences in the weight indices of the ovaries and uteri in the HS groups compared with the control group (p>0.05).
Visceral organ weights: The weights of the visceral organs (kidney, heart, liver, spleen and the adrenal glands) were all significantly higher in the two groups of HS rats compared with the control rats (Table 5). When these weights were corrected for body weights (i.e. when the weight index [organ weight/body weight] for each organ was calculated) (Table 6) all the weights of the organs (except the adrenal glands) in the high dose HS group (1.8 g/100 ml) were significantly higher than those of the control group (p<0.05 for kidney, p<0.01 for heart and p<0.001 for liver and spleen) whereas only the weights of the liver (p<0.05) and the spleen (p<0.001) were significantly higher than those of the control in the low dose HS group (0.6 g/ 100 ml).
DISCUSSION
At onset of puberty, the rats in the two HS groups were generally bigger than those in the control group as they had significantly increased body weight, visceral organs weights, body length and body mass index. This observation may suggest that the onset of puberty in the rats in the two HS groups did not depend on the attainment of a certain degree of body weight or body mass index.
Although a clear departure from the critical weight hypothesis of Frisch et al. (1977) and Frisch (1996), this observation is in agreement with the reports of several other workers (Guzman et al., 2006; Engelbergt et al., 2002; 2000; Smith and Waddell, 2000) who found no relationship between body weight and onset of puberty. Whereas these authors observed a significantly decreased body weight at onset of puberty, we observed in the present study that the body weight of the rats that consumed aqueous HS postweaning were significantly higher than that of the control rats at onset of puberty.
| Table 4: | Effect of post-weaning consumption of aqueous extract of Hibiscus sabdariffa on ovarian and uterine weight onset of puberty |
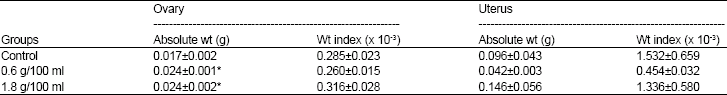 | |
N = 9 each. Values are expressed as M±SEM, * = p<0.05 compared with Control | |
| Table 5: | Effect of postweaning consumption of aqueous extract of Hibiscus sabdariffa on absolute weight of selected visceral organs at onset of puberty |
 | |
N = 9 each. Values are expressed as M±SEM, * = p<0.05 compared with Control, P = p<0.05 compared with 0.6 g/100 ml | |
| Table 6: | Effect of consumption of aqueous extract of Hibiscus sabdariffa during the postweaning period on weight index of selected visceral organs at onset of puberty |
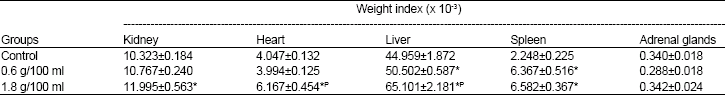 | |
N = 9 each. Values are expressed as M±SEM, * = p<0.05 compared with Control, P = p<0.05 compared with 0.6 g/100 ml | |
Since the food consumption by the rats that consumed the HS extract were lower than that of the control rats, the observed increased weight in the HS rats at onset of puberty may imply that the constituents of the HS extract may have influenced the increased weight gain in these rats. This is so because aqueous HS extract has been shown to contain substances like flavonoids, iron, vitamins A and C (Fuleki and Francis, 1968; Clydesdale, 1979; Duke and Francis, 1973; Morton, 1987; Dafallah and Al-Mustafa, 1996; Appel, 2003; Adigun et al., 2006) which have been shown to increase body weight gain (Jain et al., 2008; Wu et al., 2008; Christian et al., 2003; Hilakivi-Klarke et al., 1998; Ceesay et al., 1997). This observation is also in agreement with Guzman et al. (2006) who suggested that body weight is not the only factor that determines the onset of puberty.
Since puberty confers reproductive competence and one complete reproductive cycle of ovulation, conception, pregnancy and lactation is one of the most energy consuming activities a female mammal can undertake, particularly in species that bear multiple young (Engelbregt et al., 2001), the existence of a metabolic pathway linking nutrition with the neuroendocrine reproductive system ensures the appropriate coupling of the onset of reproductive activity with the period of adequate food availability so that scarce energy resources are not wasted on reproductive efforts that are unlikely to succeed.
Body weight, Body Mass Index (BMI) and plasma leptin levels have been reported to be highly positively correlated (Butzow et al., 1999; Maffei et al., 1995; Considine et al., 1996; Butte et al., 1997). In the present study, the observation that puberty onset in the HS rats was not related to BMI (a crude index of food availability), and thus circulating leptin level was unexpected. This is because leptin has been postulated to be the metabolic signal that informs the brain that energy reserves are adequate to support pregnancy (Bronson and Manning, 1991; Campfield et al., 1996; Chehab et al., 1997) thus triggering puberty by interacting with the GnRH neuron (Lee, 1995) and causing changes in the frequency and magnitude of GnRH pulses which has been shown to herald the onset of puberty (Terasawa, 1995).
The delayed onset of puberty coupled with high body weight and body mass index in the rats that drank HS in this study may therefore suggest either a depression of the leptin signaling pathway that normally informs the brain that energy resources are adequate to support pregnancy thus triggering puberty or the increased weight gain was a direct consequence of the growth promoting effects of the various constituents of the HS extract. The increased visceral organs weights and specifically the organomegalies in the rats that drank the aqueous extract of HS may suggest underlying metabolic and other systemic diseases in these rats.
Conclusion: In conclusion, the present study suggests that postweaning nutritional perturbation may affect the phenotype later in life. Clearly, further work is needed in this area so as to identify the specific derangement(s) induced by consumption of aqueous HS during the postweaning period.
REFERENCES
- Adigun, M.O., O.D. Ogundipe, J.I. Anetor and A.O. Odetunde, 2006. Dose-dependent changes in some haematological parameters during short-term administration of Hibiscus sabdariffa Calyx aqueous extract (zobo) in Wistar albino rats. Afr. J. Med. Med. Sci., 35: 73-77.
PubMed - Barash, I.A., C.C. Cheung, D.S. Weigle, H. Ren and E.B. Kabigting et al., 1996. Leptin is a metabolic signal to the reproductive system. Endocrinology, 137: 3144-3147.
CrossRefPubMedDirect Link - Bronson, F.H. and J.M. Manning, 1991. The energetic regulation of ovulation: A realistic role for body fat. Biol. Reprod., 44: 945-950.
CrossRef - Butzow, T.L., J.M. Moilanen and M. Lehtovirta, 1999. Serum and follicular fluid leptin during in vitro fertilization: Relationship among leptin increase, body fat mass and reduced ovarian response. J. Clin. Endocrinol. Metab., 84: 3135-3139.
Direct Link - Butte, N.F., J.M. Hopkinson and M.A. Nicolson, 1997. Leptin in human reproduction: Serum leptin levels in pregnant and lactating women. J. Clin. Endocrinol. Metab., 82: 585-589.
Direct Link - Campfield, L.A., F.J. Smith and P. Burn, 1996. The OB protein (leptin) pathway-a link between adipose tissue mass and central neural networks. Hormone Metab. Res., 28: 619-632.
PubMed - Chehab, F.F., M.E. Lim and R. Lu, 1996. Correction of the sterility defect in homozygous obese female mice by treatment with the human recombinant leptin. Nat. Genet., 12: 318-320.
CrossRefPubMedDirect Link - Chehab, F.F., K. Mounzih, R. Lu and M.E. Lim, 1997. Early onset of reproductive function in normal female mice treated with leptin. Science, 275: 88-90.
CrossRef - Cheung, C.C., J.E. Thornton, J.L. Kuijper, D.S. Weigle, D.K. Clifton and R.A. Steine, 1997. Leptin is a metabolic gate for the onset of puberty in the female rat. Endocrinology, 138: 855-858.
Direct Link - Christian, P., S.K. Khatry, J. Katz, E.K. Pradhan and S.C. Le Clerq et al., 2003. Effects of alternative maternal micronutrient supplements on low birth weight in rural Nepal: Double blind randomised community trial. Br. Med. J., 326: 571-573.
Direct Link - Considine, R.V., M.K. Sinha, M.L. Heiman, A. Kriaucinas and T.W. Stephens et al., 1996. Serum immunoreactive leptin concentrations in normal weight and obese humans. N. Engl. J. Med., 334: 292-295.
Direct Link - Dafallah, A.A. and Z. Al-Mustafa, 1996. Investigation of the anti-inflammatory activity of Acacia nilotica and Hibiscus sabdariffa. Am. J. Chin. Med., 24: 263-269.
CrossRefPubMedDirect Link - Engelbregt, M.J.T., M.E.C.A.M. Houdijk, C. Popp-Snijders and H.A.D.V. de Waal, 2000. The effects of intrauterine growth retardation and postnatal undernutrition on onset of puberty in male and female rats. Pediatr. Res., 48: 803-807.
Direct Link - Engelbregt, M.J.T., M.M. van Weissenbruch, C. Popp-Snijders, P. Lips and H.A.D.V. de Waal, 2001. Body mass index, body composition and leptin at onset of puberty in male and female rats after intrauterine growth retardation and after early postnatal food restriction. Pediatr. Res., 50: 474-478.
Direct Link - Engelbregt, M.J.T., M.M. van Weissenbruch, C. Popp-Snijders and H.A.D.V. de Waal, 2002. Delayed first cycle in intrauterine growth-retarded and postnatally undernourished female rats: Follicular growth and ovulation after stimulation with pregnant mare serum gonadotropin at first cycle. J. Endocrinol., 173: 279-304.
CrossRef - Frisch, R.E., D.M. Hegsted and K. Yoshinaga, 1975. Body weight and food intake at early estrus of rats on a high fat diet. Proc. Natl. Acad. Sci. USA., 72: 4172-4176.
Direct Link - Frisch, R.E., D.M. Hegsted and K. Yoshinaga, 1977. Carcass components at first estrus of rats on high-fat and low-fat diets: Body water, protein and fat. Proc. Natl. Acad. Sci. USA., 74: 379-383.
Direct Link - Gruaz, N.M., M. Lalaoui, D.D. Pierroz, P. Englaro, P.C. Sizonenko, W.F. Blum and M.L. Aubert, 1998. Chronic administration of leptin into the lateral ventricle induces sexual maturation in severely food-restricted female rats. J. Neuroendocrinol., 10: 627-633.
PubMedDirect Link - Guzman, C., R. Cabrera, M. Cardenas, F. Larrea, P.W. Nathanielsz and E. Zambrano, 2006. Protein restriction during foetal and neonatal development in the rat alters reproductive function and accelerates reproductive aging in female progeny. J. Physiol., 572: 97-108.
CrossRefDirect Link - Hilakivi-Clarke, L., E. Cho and R. Clarke, 1998. Maternal genistein exposure mimics the effects of estrogen on mammary gland development in female mouse offspring. Oncol. Rep., 5: 609-616.
Direct Link - Ingalls, A.M., M.M. Dickie and G.D. Snell, 1950. Obese, a new mutation in the house mouse. J. Heredity, 41: 317-318.
Direct Link - Iyare, E.E. and F.E. Iyare, 2006. Effect of in utero exposure to an aqueous extract of Hibiscus Sabdariffa on offspring's postnatal growth in sprague-dawley rats. Nig. J. Health Biomed. Sci., 5: 27-30.
Direct Link - Iyare, E.E. and F.E. Iyare, 2006. Lactational exposure to an aqueous extract of Hibiscus sabdariffa (Hs) accelerates offspring's early postnatal growth in sprague-dawley rats. J. Exp. Clin. Anat., 5: 6-9.
CrossRefDirect Link - Jain, S.K., R. Wise, K. Yanamandra, R. Dhanireddy and J.A. Jr. Bocchini, 2008. The effect of maternal and cord-blood vitamin C, vitamin E and lipid peroxide levels on newborn birth weight. Mol. Cell Biochem., 309: 217-221.
Direct Link - Maffei, M.J., J. Halaas, E. Ravussin, R.E. Pratley and G.M. Lee et al., 1995. Leptin levels in human and rodent: Measurement of plasma leptin and ob RNA in obese and weight-reduced subjects. Nat. Med., 1: 1155-1161.
CrossRefDirect Link - Mounzih, K., R. Lu and F.F. Chehab, 1997. Leptin treatment rescues the sterility of genetically obese ob/ob males. Endocrinology, 138: 1190-1193.
Direct Link - Schneider, J.E. and G.N. Wade, 1989. Availability of metabolic fuels controls estrous cyclicity of Syrian hamsters. Science, 244: 1326-1328.
CrossRefDirect Link - Smith, J.T. and B.J. Waddell, 2000. Increased fetal glucocorticoid exposure delays puberty onset in postnatal life. Endocrinolology, 141: 2422-2428.
Direct Link - Terasawa, E., 1995. Control of luteinizing hormone-releasing hormone pulse generation in nonhuman primates. Cell Mol. Neurobiol., 15: 141-164.
CrossRef - Wu, X., T. Iguchi, N. Itoh, K. Okamoto, T. Takagi, K. Tanaka and T. Nakanishi, 2008. Ascorbic acid transported by sodium-dependent vitamin C transporter 2 stimulates steroidogenesis in human choriocarcinoma cells. Endocrinol. J., 149: 73-83.
Direct Link - Zhang, Y., R. Proenca, M. Maffei, M. Barone, L. Leopold and J.M. Friedman, 1994. Positional cloning of the mouse obese gene and its human homologue. Nature, 372: 425-432.
CrossRefDirect Link