
Pramote Paengkoum
School of Animal Production Technology, Institute of Agricultural Technology,Suranaree University of Technology, Muang, Nakhon Ratchasima, 30000, Thailand
M. Wanapat
Department of Animal Science, Faculty of Agriculture, Khon Kaen University, Muang, Khon Kaen, 40002, Thailand
Pakistan Journal of Nutrition
Year: 2009 | Volume: 8 | Issue: 8 | Page No.: 1229-1234







ABSTRACT
The objective of this study was to evaluate the effect of varying levels of sunflower seed meal on feed intake, digestibility and rumen fermentation of goats fed corn silage as roughages. Eight growing crossed Thai native x Anglo-Nubian goats between 8-10 months of age and pre-trial average body weight of 14.2±1.7 kg were allotted into two groups on the basis of body weight in a randomized double 4 x 4 Latin square design to investigate the utilization of concentrate mixtures of varying levels of Sunflower Seed Meal (SSM). The four concentrate mixtures had SSM included at 0% (control), 7.3 % (SSM1), 14.5% (SSM2) and 22.0% (SSM3) in concentrate as replacing Soybean Meal (SBM) with SSM as 25% CP, 50% CP and 75% CP, respectively. On average, all parameters measured, including Dry Matter Intake (DMI), nutrients digestibility, ruminal ammonia-N (NH3-N), ruminal total Volatile Fatty Acid (total VFA), individual VFAs concentration (mM/L), microbial N supply and N retention increased (p<0.05) with the addition of SSM in concentrate up to 14.5% (SSM2) in concentrate, thereafter decreased (p<0.05) in goats fed SSM 22% (SSM3). The results indicated that replacing SBM with SSM as 50 %CP (SSM2), resulted in higher animal performance as a consequence of improved ruminal fermentation, microbial yield and N balance, but decreased (p<0.05) with 22.0% SSM diet. It is concluded that sunflower seed meal can successfully replace soybean meal 50% CP as a protein source in growing goats.
PDF Abstract XML References Citation
How to cite this article
Pramote Paengkoum and M. Wanapat, 2009. Utilization of Concentrate Supplements Containing Varying Levels of Sunflower
Seed Meal by Growing Goats Fed a Basal Diet of Corn Silages. Pakistan Journal of Nutrition, 8: 1229-1234.
DOI: 10.3923/pjn.2009.1229.1234
URL: https://scialert.net/abstract/?doi=pjn.2009.1229.1234
DOI: 10.3923/pjn.2009.1229.1234
URL: https://scialert.net/abstract/?doi=pjn.2009.1229.1234
INTRODUCTION
Most concentrates for goat are grain-based to increase their energy concentration, which typically improves gain efficiency and cost of gain. The use of Soybean Meal (SBM) as a source of protein in animal feed has been well established for many years. However, high prices and fluctuation in production have raised the interest in alternative protein sources for feeding ruminants. The increased world production of sunflower has increased interest in Sunflower Seed Meal (SFM) as a high quality by-products for ruminants (Nishino et al., 1980; Titi, 2003). The SFM, like SBM, is a high-protein supplement, contains 30-46% crude protein, 13-15% crude fiber, 9-12 MJ ME, Neutral Detergent Fiber (NDF) 47% and Ether Extract (EE) 1.5% (Nishino et al., 1980; Titi, 2003; Irshaid et al., 2003). Recently, SFM is becoming available in Thailand for ruminant nutrition and as a by-product of extraction of sunflower oil locally. SFM could be used as a substitute for SBM. As a protein supplement, SFM could replace SBM in rations of small ruminants with similar gain and feed efficiency (Richardson et al., 1981; Irshaid et al., 2003).
Therefore the objectives of this research were to study the effect of SFM on feed intake, digestibility of nutrients, microbial protein synthesis, nitrogen balance and on the performance of Thai native x Anglo-Nubian growing goats.
MATERIALS AND METHODS
Animals and management: Eight growing crossed Thai native x Anglo-Nubian goats between 8-10 months of age and pre-trial average body weight of 14.2±1.7 kg were allotted into two groups on the basis of body weight in a randomized double 4 x 4 Latin square design to investigate the utilization of concentrate mixtures of varying levels of Sunflower Seed Meal (SSM). The four concentrate mixtures had SSM included at 0% (control), 7.3% (SSM1), 14.5% (SSM2) and 22.0% (SSM3) in concentrate as replacing Soybean Meal (SBM) with SSM as 25% CP, 50% CP and 75% CP, respectively. The goats were housed in individual pens and allowed 3 weeks to adapt to the experimental conditions. The goats were fed a basal diet containing corn silages supplemented with mineral and vitamin. The experimental diets were iso-nitrogenous (CP = 15%) and iso-energetic and based on corn silage, soybean meal, cassava pulp, molasses, urea, mineral and vitamins. Rations were formulated in accordance with NRC (1981) to achieve a medium activity and medium of growth rate. Drinking water was freely available to the animals. The dietary treatments and compositions are shown in Table 1.
Experimental procedure: The experiment consisted of three weeks of adaptation, following by four experimental periods. The duration of each period was 32 days, i.e., three weeks of adjustment followed by 12 days of measurements. The latter consisted of 2 days of adaptation to the metabolic crates, 7 days of digestibility and N balance studies, 2 days of rumen fluid and blood sampling. Samples of feed refusal, faeces and urine were collected before feeding morning to determine digestibility and N balance.
Sampling methods: A daily faeces of each goat was weighed and a 10% sub-sample collected and stored at -20oC. Samples were dried (60oC) and ground through 1 mm sieve and stored until analysis.
Daily urine output was collected into a plastic container (containing 25 ml of 10% H2SO4). Approximately 10% of the volume was sampled and stored at -20oC pending energy and N analysis. A separate urine sample was collected for determination of Purine Derivatives (PD). The urine sample was diluted 4 times follows (to prevent crystallization of uric acid during storage), filtered through Whatman cellulose membranes (25 mm, 0.2 microns) attached to a syringe, thereafter, frozen at -20oC for later analysis of PD content using High Performance Liquid Chromatography (HPLC) according to Balcells et al. (1992).
Rumen fluid samples from all goats were collected using a stomach tube at 3 h post-feeding during the digestibility trial. It was strained through 4 layers of cheese cloth and pH measured immediately using a pH meter (Mettler Toledo MP 125) fitted with a combined electrode. The rumen fluid was then acidified with H2SO4 (50%, v/v) and stored at -20oC for analyses of ammonia and VFA.
Blood samples were taken from the jugular vein at 3 h post-feeding and after rumen fluid were sampled. The blood samples were prior to plasma separation by centrifugation (3,000 x g for 15 min) and plasma stored at -20oC for urea analysis.
Chemical analysis and calculations: Feed samples were collected twice a week. Representative samples of feed and faeces collected during the digestibility trial were analyzed according to AOAC (1985) for DM, ash and CP and fiber components (Van Soest et al., 1991). Apparent digestibilities were calculated using equations of Schnieder and Flatt (1975).
Total VFA and molar proportions of acetic, propionic and butyric of rumen fluids were determined by Shimatzu GC-14 gas chromatography (Shimatzu, Japan) fitted with a Flame Ionization Detector (FID) and a packed column 5% Thermon-3000, Shincarbon A 60/80. Nitrogen was used as the carrier gas at 40 ml/min and the oven temperature was maintained at 220oC; injection and FID temperatures were fixed at 260oC. Plasma urea was determined by using a urea test kit (Sigma Diagnostics INFINITYTM BUN Reagent).
The purine derivatives allantoin, uric acid, hypoxanthine and xanthine were analyzed by reverse-phase High Performance Liquid Chromatography (HPLC), which consisted of a multi-solvent delivery system Model 600 E (Water, USA), an injector Model 712, a multi-wavelength detector Model 490E, set to 205 nm and a double 4.6 x 250 mm, C-18 reverse-phase column, according to the technique of Balcells et al. (1992). Production of microbial N was calculated using the equation of Chen et al. (1992).
Statistical analysis: Data were analyzed as a 4 x 4 Latin square design using the General Linear Model (GLM) procedure of the Statistical Analysis System Institute SAS (1988). Duncan’s New Multiple Range Test and Orthogonal Contrast Analysis (Steel and Torrie, 1980) were used to compare treatment means. Unless otherwise noted, significance was declared at p<0.05 and non-significance was declared at p>0.05.
RESULTS AND DISCUSSION
The CP, ash, NDF and ADF contents of Sunflower Seed Meal (SSM) on dry matter basis were 34.4%, 7.2%, 47.4 and 33.1%, respectively (Table 1). Crude protein of Soybean Meal (SBM), cassava pulp and corn silage were 44.7, 2.1 and 6.3%, respectively. The inclusion of SSM further raised the NDF and crude fiber content are shown in Table 2.
| Table 1: | Proximate chemical composition of feedstuffs |
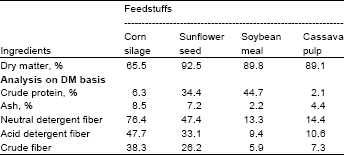 | |
| Table 2: | Percentage composition of concentrated mixtures and chemical compositions |
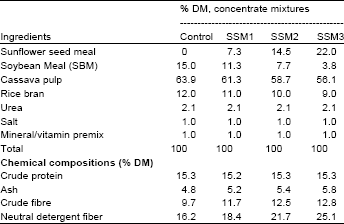 | |
| Control = SBM as a main source of protein in concentrate, SSM1 = replacing SBM with SSM as 25% CP, SSM2 = replacing SBM with SSM as 50% CP, SSM3 = replacing SBM with SSM as 75% CP | |
| Table 3: | Effect of varying levels of Sunflower Seed Meal (SSM) in concentrate on feed intake, nutrients digestibility and body weight change of goats |
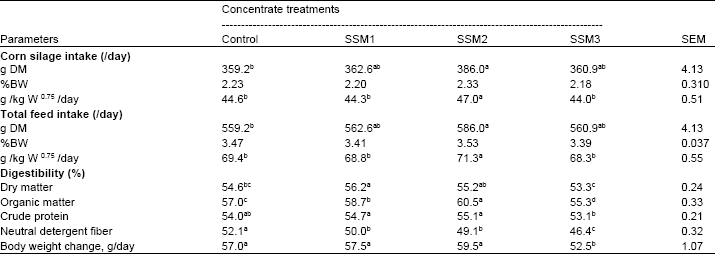 | |
| SEM = standard error of the mean. a,b,cMeans within row with different superscripts differ significantly (p<0.05). Control = SBM as a main source of protein in concentrate, SSM1 = replacing SBM with SSM as 25% CP, SSM2 = replacing SBM with SSM as 50% CP, SSM3 = replacing SBM with SSM as 75% CP | |
| Table 4: | Effect of varying levels of Sunflower Seed Meal (SSM) in concentrate on ruminal pH, ruminal NH3-N, Total Volatile Fatty Acid (TVFA), individual VFAs and Plasma Urea Nitrogen (PUN) of goats |
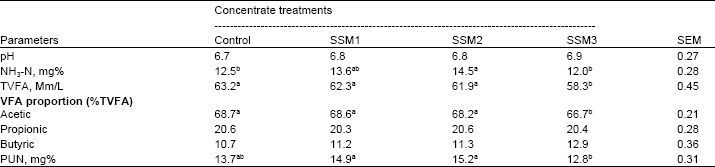 | |
| SEM = standard error of the mean. a,b,cMeans within row with different superscripts differ significantly (p<0.05). Control = SBM as a main source of protein in concentrate, SSM1 = replacing SBM with SSM as 25% CP, SSM2 = replacing SBM with SSM as 50% CP, SSM3 = replacing SBM with SSM as 75% CP | |
Table 3 shows the DMI, nutrient digestibility and body weight change of goats fed varying SSM in concentrates. Dry matter intake based on g/kg BW0.75, increased (p<0.05) with the addition of SSM in concentrate up to 14.5% (SSM2) in concentrate, thereafter decreased (p<0.05) in goats fed SSM 22% (SSM3). Moreover, DMI (g/day, g/kg BW0.75) in goats fed 14.5% SSM significantly higher (p<0.05) than goats fed the control (SBM). Organic matter, CP and NDF digestibility and body weight change increased (p<0.05) with the addition of SSM in concentrate up to 14.5% in concentrate, thereafter decreased (p<0.05) in goats fed SSM 22%. Body weight change of goats fed the control, 7.3% SSM and 14.5% SSM were not significantly (p>0.05) different in among treatments. This insignificant difference might be a result of the higher fiber content of the SSM and through less ability of digestion due to incomplete rumen functioning of those goats (Nishino et al., 1980; Titi, 2003; Mir et al., 2005).
Ruminal NH3-N, TVFA, acetic acid and PUN concentrations increased (p<0.05) with the addition of SSM in concentrate up to 14.5% in concentrate, thereafter decreased (p<0.05) in goats fed SSM 22% (Table 4). However, ruminal ph, propionic acid and butyric acid were not significantly (p>0.05) different in among treatements. Mould and Ørskov (1993); Wanapat et al. (2000) demonstrated that cellulose digestion is limited when ruminal pH reaches values below 6.0. Ruminal VFA concentrations and fermentation rates are correlated (Leng and Leonard, 1965; McDonald et al., 1995) and this relationship was also demonstrated in the present study (ruminal VFAs and digestibility). These results are in contrast to Koster et al. (1996) reported that TVFA increased dramatically in response to supplemental rumen degradable protein fed to beef cows.
The excretion of urinary Purine Derivatives (PD) and microbial N supply are shown in Table 5. The results showed that total PD excretion, allantoin, uric acid, hypoxanthine, xanthine, microbial N synthesis (g N/day) and efficiency of microbial N supply (g N/ kg OM Apparently Digested in the Rumen (OMDR) of goats fed the control, 7.3 and 14.5% SSM in concentrates were significantly higher (p<0.05) than in goats fed 22.0% SSM.
| Table 5: | Effect of varying levels of Sunflower Seed Meal (SSM) in concentrate on urinary perine derivatives and microbial N supply of goats |
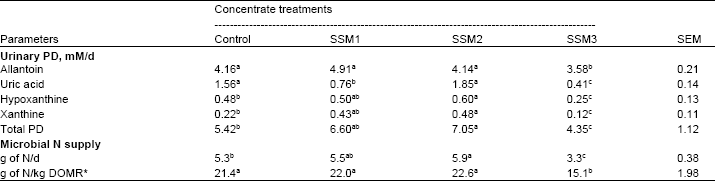 | |
| SEM = standard error of the mean. a,b,cMeans within row with different superscripts differ significantly (p<0.05). Control = SBM as a main source of protein in concentrate, SSM1 = replacing SBM with SSM as 25% CP, SSM2 = replacing SBM with SSM as 50% CP, SSM3 = replacing SBM with SSM as 75% CP | |
| Table 6: | Effect of varying levels of Sunflower Seed Meal (SSM) in concentrate on Nitrogen (N) balance of goats |
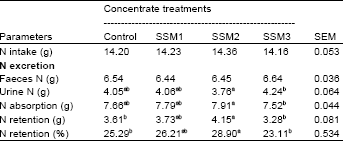 | |
| SEM = standard error of the mean. a,b,cMeans within row with different superscripts differ significantly (p<0.05). Control = SBM as a main source of protein in concentrate, SSM1 = replacing SBM with SSM as 25% CP, SSM2 = replacing SBM with SSM as 50% CP, SSM3 = replacing SBM with SSM as 75% CP | |
There were no effects of dietary treatment on N intake and faeces N. However, N absorption (g/day) and N retention (g/day, %) increased (p<0.05) with the addition of SSM in concentrate up to 14.5% in concentrate, thereafter decreased (p<0.05) in goats fed 22.0% SSM. However, urine N of goats fed 14.5% SSM was lower (p<0.05) than goats fed 22.0% SSM.
Results of this study showed that the overall effect of dietary treatments were no significant differences on almost parameters of goats fed the control (0% SSM), 7.3% SSM and 14.5% SSM (replacing SBM with SSM as 0, 25 and 50% CP). These results agreed with earlier results indicated that SSM could replacing SBM as a protein source for ruminant feeding (Isobel et al., 1990; Schingeothe et al., 1996; Economides, 1998; Titi, 2003). Growth results of the control (SBM) and 7.3% SSM or 14.5% SSM agreed with other studies that showed no differences in these parameters (Economides and Koumas, 1999; Titi, 2003). On the other hand, earlier studies indicated that SSM proteins are more soluble under ruminal conditions than SBM proteins but used less efficiently due to less available carbohydrate and high fiber of SSM (Schingeothe et al., 1977; Economides, 1998; Irshaid et al., 2003).
Higher DMI and nutrient digestibility of goats fed replacing SBM with SSM 50% of CP than the control treatment assumed that no effect on palatability during feeding. These results agreed with other results reported similar findings (Stake et al., 1973; Economides and Koumas, 1999; Titi, 2003). Microbial N (g N/day) and the efficiency of microbial N supply [g N/kg OMDR] were significantly higher (p<0.05) in goats fed 14.5% SSM than 22.0% SSM treatments. The 21.4 TO 22.6 g N/kg OMDR obtained in this study for goats the control, 7.3% SSM and 14.5% SSM was approaching the values of 17-35 g N/kg OMDR suggested in the literature (ARC, 1984; Czerkawski, 1986; Sinclair et al., 1995; Paengkoum et al., 2006a, 2006b). Nitrogen excretions, particulary urine N increased with addition SSM. Kebreab et al. (2002) also reported similar finding and suggested that high urine N excretion has a higher adverse impact on environmental pollution. The above results thus support earlier reports that total N and particularly urine N excretions could be reduced through diet manipulation (Tamminga, 1996; Castillo et al., 2001; Kebreab et al., 2002).
Conclusions: There was an overall improvement in the performance of goats fed 7.3% SSM and 14.5% SSM as compared to the control diets. All the parameters measured indicated that although there was an overall improvement in DMI, nutrient digestibility, microbial protein synthesis and N utilization for goats fed additional SSM, there was no advantage of feeding them with more than replacing SBM with SSM as 50% CP. Results of the present study showed that approximate replacing SBM with SSM as about 50% CP to SBM as the expensive protein source could achieve an impressive about 60 g/d BW gain for growing goats. We postulated that addition of fibrolytic enzymes to the above diet (high SSM) would further enhance both of the efficiency of microbial synthesis and host animal performances.
ACKNOWLEDGEMENT
The authors acknowledges the Suranaree University of Technology (SUT) and The Thailand Research Fund (TRF) for financial and facilities support of this research.
REFERENCES
- Balcells, J., J.A. Guada and J.M. Peiro, 1992. Simultaneous determination of allantoin and oxypurines in biological fluids by high performance liquid chromatography. J. Chromatogr., 575: 153-157.
PubMedDirect Link - Castillo, A.R., E. Kebreab, D.E. Beever, J.H. Barbi, J.D. Sutton, H.C. Kirby and J. France, 2001. The effect of protein supplementation on nitrogen utilization in lactating dairy cows fed grass silage diets. J. Anim. Sci., 79: 247-253.
PubMedDirect Link - Chen, X.B., Y.K. Chen, M.F. Franklin, E.R. Orskov and W.J. Shand, 1992. The effect of feed intake and body weight on purine derivative excretion and microbial protein supply in sheep. J. Anim. Sci., 70: 1534-1542.
Direct Link - Economides, S., 1998. The nutritive value of sunflower meal and its effect on replacing cereal straw in the diets of lactating ewes and goats. Livestock Prod. Sci., 55: 89-97.
CrossRefDirect Link - Isobel, C.V., R. Hill and R.C. Campling, 1990. A note on the use of rapeseed, sunflower and soybean meals as a protein sources in compound foods for milking cattle. Anim. Prod., 50: 541-543.
Direct Link - Kebreab, E., J. France, J.A. Mills, R. Allison and J. Dijkstra, 2002. A dynamic model of N metabolism in the lactating dairy cow and an assessment of impact of N excretion on the environment. J. Anim. Sci., 80: 248-259.
Direct Link - Koster, H.H., R.C. Cochran, E.C. Titgemeyer, E.S. Vanzant, D. Aldelgadir and G.S. Jean, 1996. Effect of increasing degradable protein intake on intake and digestion of low-quality, tall grass-prairie forage by beef cows. J. Anim. Sci., 74: 2473-2482.
Direct Link - Leng, R.A. and G.J. Leonard, 1965. Measurement of the rates of production of acetic, propionic and butyric acids in the rumen of sheep. Br. J. Nutr., 19: 469-484.
PubMedDirect Link - Mir, P.S., M. Ivan, G.L. Mears, C.M. Ross, T. Entz and Z. Mir, 2005. Effects of dietary protein and sunflower seed supplementation on physio-chemical characteristics of small intestinal digesta and plasma cholecystokinin concentrations in lambs. Small Rumin. Res., 58: 163-173.
CrossRef - Richardson, R.C., N.R. Ratcliff and J. Albin, 1981. SFM as a protein supplement for growing ruminants. J. Anim. Sci., 53: 557-563.
Direct Link - Schingeothe, J.D., A.J. Rook and F. Ludens, 1977. Evaluation of sunflower meal as a protein supplement for lactating cows. J. Dairy Sci., 60: 591-595.
Direct Link - Schingoethe, D.J., M.J. Brouk, K.D. Lightfield and R.J. Baer, 1996. Lactational responses of dairy cows fed unsaturated fat from extruded soybeans or sunflower seeds. J. Dairy Sci., 79: 1244-1249.
CrossRefPubMedDirect Link - Sinclair, L.A., P.C. Garnsworthy, J.R. Newbold and P.J. Battery, 1995. Effects of synchronizing the rate of dietary energy and nitrogen release in diets with a similar carbohydrate composition on rumen fermentation and microbial protein synthesis in sheep. J. Agric. Sci., 124: 463-472.
CrossRefDirect Link - Tamminga, S., 1996. A review on environmental impacts of nutritional strategies in ruminants. J. Anim. Sci., 74: 3112-3124.
Direct Link - Irshaid, R.H., M.Y. Harb and H.H. Titi, 2003. Replacing soybean meal with sunflower seed meal in the ration of Awassi ewes and lambs. Small Rumin. Res., 50: 109-116.
Direct Link - van Soest, P.J., J.B. Robertson and B.A. Lewis, 1991. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci., 74: 3583-3597.
CrossRefPubMedDirect Link - Wanapat, M., O. Pimpa, A. Petlum and C. Wachirapakorn, 2000. Participation scheme of smallholder dairy farmers in the NE, Thailand on improving feeding systems. Asian-Aus. J. Anim. Sci., 13: 830-836.
Direct Link