
A. Moghtaderi
Department of Geology, College of Sciences, Payam Noore University, Evaz, Fars province, Iran
M. Taghavi
Department of Plant Protection, College of Agriculture, Shiraz University, Shiraz, Iran
R. Rezaei
Department of Plant Protection, College of Agriculture, Shiraz University, Shiraz, Iran
Pakistan Journal of Nutrition
Year: 2009 | Volume: 8 | Issue: 7 | Page No.: 1083-1092







ABSTRACT
Arid and semi-arid regions are characterized by sparse vegetation or absence of vegetation cover. The absence of a dense distribution of macrophytes (higher plants), much of arid and semi-arid surfaces are covered by microphytic communities of small non-vascular plants. An important group of organisms comprising soil crusts in such habitats are cyanobacteria. In this research the genus and species of cyanobacteria were detected at the Chadormalu desert, Yazd Province of Iran. The study showed that cyanobacteria (Microcoleous vaginatus, Nostoc.sp, Microcystis.sp, Ocillatoria.sp, Chroococcuss.sp, Chroococcidiopsis, Ocillatoria.sp, Chroococcuss.sp, Microcystis.sp) at Chadormalu Desert are comparable with other Deserts in cold and warm dry conditions.
PDF Abstract XML References Citation
How to cite this article
A. Moghtaderi, M. Taghavi and R. Rezaei, 2009. Cyanobacteria in Biological Soil Crust of Chadormalu Area, Bafq Region in Central Iran. Pakistan Journal of Nutrition, 8: 1083-1092.
DOI: 10.3923/pjn.2009.1083.1092
URL: https://scialert.net/abstract/?doi=pjn.2009.1083.1092
DOI: 10.3923/pjn.2009.1083.1092
URL: https://scialert.net/abstract/?doi=pjn.2009.1083.1092
INTRODUCTION
Arid and semi-arid regions are characterized by sparse vegetation or absence of vegetation cover. In the absence of a dense distribution of macrophytes (higher plants), much of arid and semi-arid surfaces are covered by microphytic communities of small non-vascular plants. These microphytic communities, containing mosses, lichens, algae, fungi, cyanobacteria, (bluegreen algae) and bacteria, in various combinations, form microphytic crust over and within a wide range of soil and rock substrates like limestones, chalk, dolomite, flint, sandstone, granite, sandy soil, shale stone, losses and dune sand (Danin et al., 1975; Springer West, 1990). An important group of organisms comprising soil crusts in such habitats are cyanobacteria (McGregor and Johnson, 1971; Belnap, 1990). They are usually the primary components of soil crust. Since cyanobacteria colonize the soil faster than the other microphytic communities, they usually represent an early stage in the soil crust succession. Cyanobacteria soil crust is dominant in regions of less than 100 mm of rain. They are well adapted for primary colonization of arid environments due to their extraordinary ability to survive desiccation and extreme temperatures (up to 70oC), high pH and salinity (Springer West, 1990). According to Karnieli et al. (1999), Konhauser (2006) and Waterbury (2006) cyanobacteria structure is similar to that of bacteria, but their photosynthetic mechanism resembles that of green algae. Cyanobacteria have the common chlorophyll a but also phycoblilin pigments. The lack of complex parts, make it possible for them to occupy an ecological niche in the desert. The cyanobacteria Soil crust consisting mostly of Microcoleous vaginatus (dominant species), Scytonema, Schizothrix, Calothrix, Chroococcidiopsis, Nostoc and Phorimidium (Danin et al., 1989; Danin, 1991; Lange et al., 1992). These organisms contribute to soil stability (Danin, 1991), soil build up (Shield and Drouet, 1962), soil fertility (Zobeek and Fryrear, 1986) and to the soil water regime (Verrecchia et al., 1995). Cyanobacteria Soil Crust (CSC) has been reported from the Middle East, the African Sahel and Sahara, North and South America, Central Asia and Australia, Northern Victoria Land, Mc Murdo Dry Valleys and Ice Shelf, Antarctica and India (e.g. Springer West, 1990; Pinker and Karnieli, 1995; Karnieli and Tsoar, 1995; Cavacini, 2001; Pichel et al., 2001; Torre et al., 2003; Rios et al., 2004; Tirkey and Adhikary, 2005; Wierzchos et al., 2006). Karnieli and Tsoar (1995), demonstrated that the lack of CSC in Sinai (Egypt) due largely to man’s activities. Pinker and Karnieli (1995) suggested that anthropogenic activities, which prevent the accumulation of crust or destroy an exciting, crust, rather than the overgrazing mechanism.
West (1990) proposed that microbiotic crusts develop on sandy soil and 45% clay and silt sandy Negev (Israel) and Sinai (Egypt) desert. This author noted that microbiotic cover is smoother where associated with relatively high bulk densities, but attributes this characteristic to precipitation and temperature regims. Pichel et al. (2001), distinguished cyanobacterias (e.g. M. vaginatous, Schizothrix spp., Phorimidium spp., Scytonema sp., Nostoc sp.) at sandy silt, gypsum and shale soils at Colorado Desert. They suggested that Microcoleous vaginatous, dominant in most samples. Wierzchos et al. (2006) showed that halite evaporate rocks from the driest part of the Atacama Desert (Chile) are colonized by Endolithic cyanobacteria. This colonization occur just a few millimeters beneath the rock surface, occupying spaces among salt crystals.
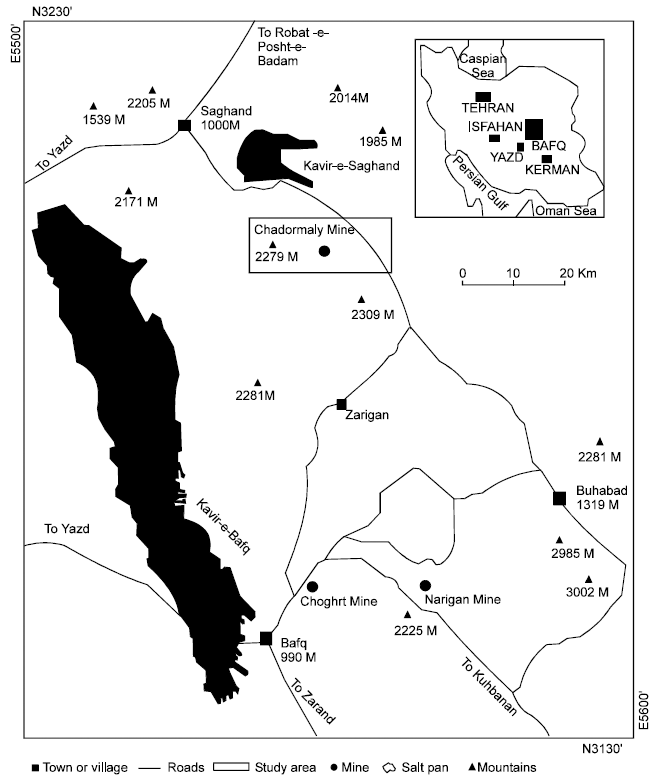 | |
| Fig. 1a: | Location of Chadormalu desert area in central Iran (Modified from Forster and Jafarzadeh, 1994) |
Their Work revealed that these communities are composed of extremely resistant Chroococcidiopsis morphospecies of cyanobacteria and associated heterotrophic bacteria. Cavacini (2001) reported 22 cyanobacterias taxa on soil from northern Victoria Land (Antarctica). Cyanobacterias were presented in 80% of the samples collected. Torre et al. (2003) showed that, microorganisms colonize the pore spaces of exposed rocks at McMurdo Dry Valleys, Antarctica. These cryptoendolithic communities are included lichens and cyanobacteria (e.g. Phorimidium) communities. Rios et al. (2004) showed Microstructural characterization of cyanobacterial mates from the McMurdo Ice Shelf, Antarctica. These authors used a broad suite of complementary techniques including optical and fluorescence microscopy, confocal scanning laser microscopy, scanning electron microscopy, microanalytical X-ray energy dispersive spectroscopy. This study proposed that Oscillatorialean taxa were the most abundant taxa and appeared to be intermixed with fine-size deposits of epicellular silica and calcium carbonate.
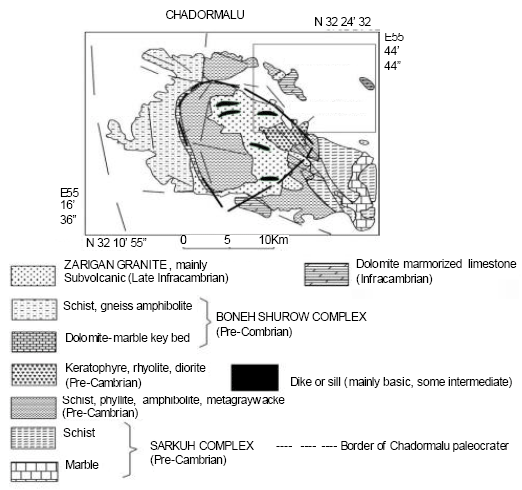 | |
| Fig. 1b: | Geological map of Chadormalu desert and Chadormalu iron oxide deposit (bordered area). |
Tirkey and Adhikary (2005), reported species of filamentous, sheath-forming cyanobacteria in the blackish-brown crusts on the upper millimeter of soils in different regions of India. Scytonema ocellatum, Scytonema chiastum, Plectectonema notatum, Lyngbya and Nostoc is dominant cyanobacteria. The main purpose of this research is to detect and compare and illustrate the genus and species of cyanobacteria at the Chadormalu desert, Yazd Province, Iran.
MATERIALS AND METHODS
The Chadormalu area (including the northern desert) is located in the Bafq metallogenic province in central Iran, about, 115 km southeast of Yazd city (55o 15'- 55o 45'E, 32o 15'-32o 25'N) (Fig. 1a, b). The extreme aridity of Chadormalu desert is due to zagros and Alborz mountain ranges in west and north, respectively which prohibit wet weather from reaching this area. Also, the salt desert (kavir) of Bafq and Saqand occupies west and north of Bafq region. The average annual rainfall is 55.7 mm and it only rains in the winter and early spring (January-April). Average minimum daily temperatures are -9.6oC in January and 25oC in May. The average maximum daily temperature is 18oC and 45oC in January and July, respectively.
Field study: As a first step 14 sampling station were selected in northern Chadormalu desert soil (e.g. sandy soil, silt, gypsum and shale) (Table 1). In each station, soil samples were collected (3 cm thick), retained in a plastic bags and then transported (in darkness, 25oC) over a period of days with no apparent loss of viability.
| Table 1: | Coordination and culturing results of cyanobacteria soil crust at Chadormalu desert area |
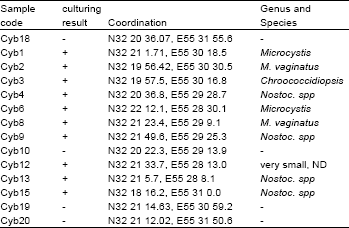 | |
| ND = Not determine | |
Isolation and purification: BG-11 media originally described by Hughes et al. (1958) and modified by Allen (1968) was used as culture media. Medium BG-11 has low phosphate content, is poorly buffered, and it has the following composition (in grams per liter of deionized distilled water): NaNO3, 1.5; K2HPO4, 0.04; MgSO4.7H2O, 0.075; CaCl2.2H2O, 0.036; Citric acid, 0.006; Ferric ammonium citrate, 0.006; EDTA (disodium magnesium salt), 0.001; Na2CO3, 0.02; Trace-metal mix A5, 1ml/lit. Trace-metal mix A5 is included: H3BO3, 2.86 g/liter; MnCl2.4H2O, 1.81 g/liter; ZnSO4.7H2O, 0.222 g/liter; Na2MoO4.2H2O, 0.39 g/liter; CuSO4.5H2O, 0.079 g/liter; Co(NO3) 2.6H2O, 0.0494 g/liter (after autoclaving and cooling, pH of medium was 7.1).
 | |
| Fig. 2: | (a-c) Microcoleous vaginatus in 4x, 10x and 40x magnification, respectively. (d-f) a Microcoleous vaginatus filamentous magnified, 40x and 100x respectively. (g) Cystine in Microcoleous vaginatus (100x). (h) Microcoleous vaginatus colony at BG-11 culture media (this figure has been captured by Canon TX1, in super macro format) |
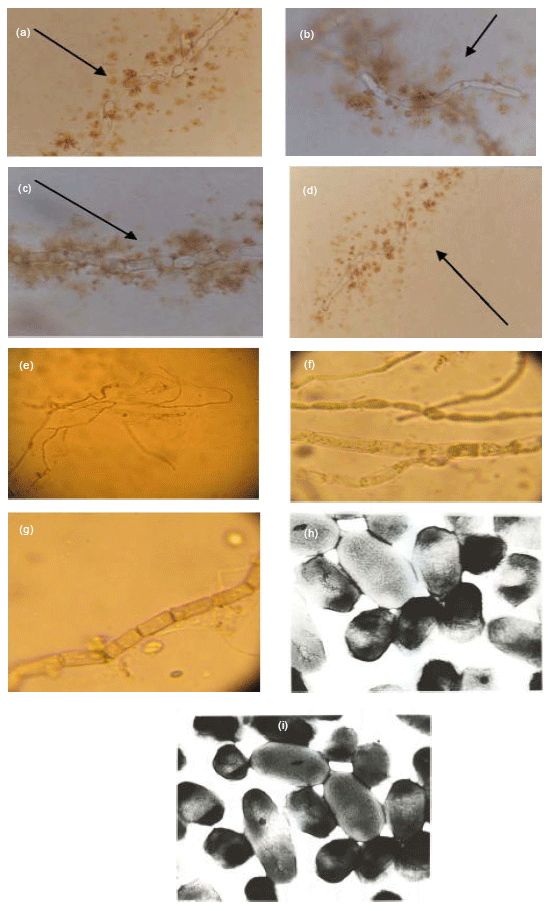 | |
| Fig. 3: | (a-d) a Nostoc filamentous with 10x, 40x, 60x, 25x magnifications (phase microscope). (e) and (f) Nostoc filamentous in 4x and 40x magnifications (TM). (g) unicellular at a Nostoc filamentous (40x). (h and (i) Microcystis colony under TEM (28000x) |
 | |
| Fig. 4: | (a-c) Ocillatoria.spp filamentous (4x, 10x and 40x); (a) TM photo (b) and (c) were prepared by phase microscope. (d) and (e) unicellular Chroococcuss.sp. (40x, TM and phase microscope, respectively). (f) and (g) Endolithic cyanobacteria (Chroococcidiopsis). (h) and (I) an artificial pond and green float Microcystis.spp colonies |
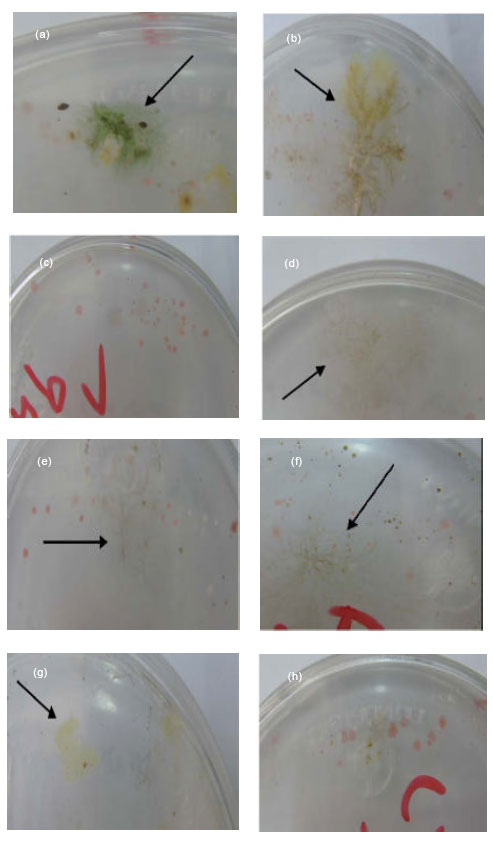 | |
| Fig. 5: | Fig. 5 (a-h):Cyanobacteria growth after 2 weeks (Ref to Fig. 4b, Fig. 5 and Table. 3). (a) Microcoleous vaginatus colony at station-8 (cyb8). (b) Nostoc.spp colony at station-4 (cyb4). (c) negative result at station-7 (cyb7). (d-f) Nostoc.spp colonies at stations 13 (cyb13), 9 (cyb9) and 15 (Cyb15) respectively. (g) Microcystis at station-1 (cyb1). (h) very small cyanobacteria colony |
Solid media were prepared by mixing, after cooling to 50oC, equal volumes of separately autoclaved double strength solutions of the mineral salts medium and either purified agar or agarose to give a final concentration of 0.6% (Castenholz, 1988; Shirai et al., 1989).
 | |
| Fig. 5 (i-o): | (i) Negative result at station-10 (cyb10). (j) Microcoleous vaginatus colony at station-2 (cyb2). (km) negative results at stations 11(cyb11), 5 (cyb5), 19 (cyb19). (n) and (O) Microcystis colony at station-6, artificial pond (Fig. 8h) and soil respectively |
Soil solution was prepared preparation in deionized distilled water and two stage serial dilution was performed on the (original) solution. The final soil solutions (final types) were cultured in the solid BG-11 mediums. During isolation and purification, cultures were incubated in a light-dark cycle with a 14-h light period and a 10 or 14 dark period for one to two weeks. The temperature of incubation was 20-30oC or room temperature (Castenholz, 1988, Rippka, 1988; Shirai et al., 1989; Waterbury, 2006).
 | |
| Fig. 6: | (a-d) manifestation of soil at stations 2, 8, 4 and 3 |
RESULTS
In this research, the cyanophytes in fully propagated colonies were identified in the solid BG-11 culture media after 1-2 weeks. Then, the cultures were examined by a phase-contrast microscope to determine genus and species. They mostly included: Microcoleous vaginatus (Fig. 2a-h), Nostoc.sp (Fig. 3a-g), Microcystis.sp (Fig. 3h-I), Ocillatoria.sp (Fig. 4a-d), Chroococcuss.sp (Fig. 4e-f) and Chroococcidiopsis (Fig. 4g-h). In the station 6, an artificial pond was found to contain Ocillatoria.sp, Chroococcuss.sp and Green float Microcystis.sp colonies (Fig. 4i-j). Fig. 5 a-o illustrates culture media after 2 weeks. Microcystis.sp colonies were changed from green to white color after 3weeks (Fig. 5l-o). This study shows that genus and species of cyanobacteria at Chadormalu Desert are comparable with other Deserts in cold and warm dry conditions (e.g. cyanobacteria taxa on soil from northern Victoria Land (Antarctica), Colorado plateau (North America) and Negev Desert (Middle East).
DISCUSSION
Biogenic crusts detections within world deserts are popular researches in microbiology and astrobiology studies (e.g. Wierzchos et al., 2006; Tirkey and Adhikary, 2005; Bhatnagar and Bhatnagar, 2005; Jafari et al., 2004; Rios et al., 2004; Cavacini, 2001; Pichel et al., 2001).
Wierzchos et al. (2006) discovered that Chroococcidiopsis can live at an endolithic environment which is an extremely dry and at the same time, saline microbial habitat. They suggested that photosynthetic microorganisms within dry evaporate rocks could be an important and previously unrecognized target for the search for life within our solar system. Tirkey and Adhikary (2005) concluded that the highly active upper layers of arid soils contain certain sheathed cyanobacteria that bind with soil particles forming a matrix protecting them from wind erosion. In addition, they are finely tuned in their physiology to the natural environmental conditions contributing organic matter and nitrogen through carbon and nitrogen fixation, thus increasing soil fertility. Bhatnagar and Bhatnagar (2005) illustrated a microbial diversity in desert ecosystems. They proposed that cyanobacteria contribute significantly to the biota of the hot arid regions in terms of primary productivity and nitrogen fixation. These authors showed that the most common genera are Microcoleous vaginatus and Nostoc.sp. Jafari et al. (2004) Compared some properties of crusted and uncrusted soils in Alagol Region in north of Golestan province, Northern Iran of Iran. They deduced that BSC (s) have important role in soil protection and chemistry in this area. Rios et al. (2004) suggested that the microscopic and microanalytical techniques provided a complementary suite of approaches for characterizing microbial mats. These researchers demonstrated that, application of a single method can result in a misleading estimate of the biodiversity and a more limited appreciation of the three dimensional organization of biogenic and nonbiogenic components within mats. Cavacini (2001) demonstrated that Filamentous cyanobacteria were greatly represented, specifically, by Phorimidium spp and Leptolyngbya frigida which were present in the majority of the samples examined on soil from northern Victoria Land. Pichel et al. (2001) indicated that soil characteristics may select for specific cyanobacteria. Gypsum crusts were most deviant from the rest, while sandy; silt and shale crusts were relatively more similar among themselves.
ACKNOWLEDGEMENT
The authors wish to thank the Plant Protection and Earth sciences Departments of Shiraz University. The research council of Pyam Noore University is also appreciated for supporting this research.
REFERENCES
- Allen, M.M., 1968. Simple conditions for growth of unicellular blue-green algae on plates. J. Phycol., 4: 1-4.
CrossRefDirect Link - Bhatnagar, A. and M. Bhatnagar, 2005. Microbial diversity in desert ecosystems. Curr. Sci., 89: 91-100.
Direct Link - Cavacini, P., 2001. Soil algae from northern Victoria Land (Antarctica). Polar Biosci., 14: 45-60.
Direct Link - Forster, H. and A. Jafarzadeh, 1994. The Bafq mining district in central Iran: A highly mineralized Infracambrian volcanic field. Econ. Geol., 89: 1697-1721.
CrossRefDirect Link - Hughes, E.D., P.R. Gorham and A. Zehnder, 1958. Toxicity of a unialgal culture of Microcystis aeruginosa. Can. J. Microbiol., 4: 225-236.
CrossRefDirect Link - Jafari, M., A. Tavili, N. Zargham, G.A. Heshmati and M.A. Zare Chahouki et al., 2004. Comparing some properties of crusted and uncrusted soils in alagol region of Iran. Pak. J. Nutr., 3: 273-277.
CrossRefDirect Link - Karnieli, A., G.J. Kidron, C. Glaesser and E. Ben-Dor, 1999. Spectral characteristics of cyanobacteria soil crust in semiarid environment. Remote Sensing Environ., 69: 67-75.
Direct Link - Lange, O.L., G.J. Kidron, B. Budel, A. Meyer, E. Kilian and A. Abeliovich, 1992. Taxonomic composition and photosynthetic characteristics of the biologic soil crust covering sand dunes in western Negev Desert. Funct. Ecol., 6: 519-527.
Direct Link - McGregor, A.N. and D.E. Johnson, 1971. Capacity of desert algal crusts to fix atmospheric nitrogen. Soil Sci. Soc. Am. Proc., 35: 843-844.
CrossRefDirect Link - Pichel, F.G., A.L. Cortez and U. Nubel, 2001. Phylogenetic and morphological diversity of cyanobacteria in soil desert crusts from the Colorado Plateau. Applied Environ. Microbiol., 67: 1902-1910.
CrossRefDirect Link - Pinker, R.T. and A. Karnieli, 1995. Characteristic spectral reflectance of a semiarid environment. Int. J. Remote Sensing, 16: 1341-1363.
Direct Link - Rios, A., C. Ascaso, J. Wierzchos, E.F. Valiente and A. Quesada, 2004. Microstructural characterization of cyanobacterial mats from the McMurdo Ice Shelf, Antarctica. Applied Environ. Microbiol., 70: 569-580.
CrossRefDirect Link - Rippka, R., 1988. Isolation and purification of cyanobacteria. Methods Enzymol., 167: 3-27.
CrossRefPubMedDirect Link - Shield, L.M. and F. Drouet, 1962. Distributions of terrestrial algae within the Nevada test site. Am. J. Bot., 49: 547-554.
Direct Link - Shirai, M., K. Matumaru, A. Ohotake, Y. Takamura, T. Aida and M. Nakano, 1989. Development of a solid medium for growth and isolation of axenic Microcystis strains (Cyanobacteria). Applied Environ. Microbiol., 55: 2569-2571.
Direct Link - Tirkey, J. and S.P. Adhikary, 2005. Cyanobacteria in biological soil crusts of India. Curr. Sci., 89: 515-521.
Direct Link - De la Torre, J.R., B.M. Goebel, E.I. Friedmann and N.R. Pace, 2003. Microbial diversity of cryptoendolithic communities from the McMurdo Dry Valleys, Antarctica. Applied Environ. Microbiol., 69: 3858-3867.
CrossRefDirect Link - Wierzchos, J., C. Ascaso and C.P. McKay, 2006. Endolithic cyanobacteria in halite rocks from the hyperarid core of the Atacama Desert. Astrobiology, 6: 1-8.
PubMedDirect Link - Zobeek, T.M. and D.W. Fryrear, 1986. Chemical and physical characteristics of windblown sediments. II. Chemical characteristics and total soil and nutrient discharge. Trans. Am. Soc. Agric. Eng., 29: 1037-1041.
Direct Link