
Mostafa Abdel Halim El-Harairy
Department of Animal Production, Faculty of Agriculture, Mansoura University, 35516, Egypt
Sherif Maghawry Mohamed Shamiah
Animal Production Research Institute, Ministry of Agriculture, Egypt
Kandil Abdel Hai Ali Attia
Department of Evaluation of Natural Resources, Environmental Studies and Research Institute, University of El-Sadat City, Egypt
LiveDNA: 20.34950
Samaa Mohamed Yehia Mehrez
Department of Animal Production, Faculty of Agriculture, Mansoura University, 35516, Egypt
Pakistan Journal of Biological Sciences
Year: 2021 | Volume: 24 | Issue: 4 | Page No.: 500-506







ABSTRACT
Background and Objective: The sex pre-selection for offspring before conception is desirable demand especially for the breeding program of farm animals. This study aimed to evaluate the preconception treatment of monovalent and divalent ions on the primary sex ratio, ovarian structures and serum minerals levels in New Zealand white rabbit does. Materials and Methods: Nine New Zealand white rabbits doe (5.4±0.61 months of age and 2.4±0.35 kg of body weight) were used. Rabbits in the 1st group were given drinking water only (control). While the 2nd and 3rd groups were given 1% of (calcium and magnesium) and (sodium and potassium) in daily drinking water, respectively for 15 days before mating. The embryos of each group were individually collected after three days of mating for primary sex detection using SRY (Sex Determining Region Y) Polymerase Chain Reaction (PCR) assay. Mineral analyses for all studied animals were weekly detected in serum before and after mating. Results: The primary sex ratio for embryos of rabbits does receive (Na+K) produced more males (69.7%) while (Ca+Mg) administrated rabbits does produce more females (72.2%). The mineral treatment leads to a significant increase in the number of corpus luteum, total embryos, follicles bleeding and a significant decline in the count of large follicles. Also, there was no significant change in serum Na and Ca levels in the treated groups compared to the control. Conclusion: The preconception administration of Ca+Mg could produce more females while Na+K could produce more males without adverse side effects on serum minerals concentration.
PDF Abstract XML References Citation
Received: November 21, 2020;
Accepted: February 10, 2021;
Published: March 15, 2021
Copyright: © 2021. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Mostafa Abdel Halim El-Harairy, Sherif Maghawry Mohamed Shamiah, Kandil Abdel Hai Ali Attia and Samaa Mohamed Yehia Mehrez, 2021. Impact of Some Ions Administration on Sex Ratio and Ovarian Structures in Rabbits. Pakistan Journal of Biological Sciences, 24: 500-506.
DOI: 10.3923/pjbs.2021.500.506
URL: https://scialert.net/abstract/?doi=pjbs.2021.500.506
DOI: 10.3923/pjbs.2021.500.506
URL: https://scialert.net/abstract/?doi=pjbs.2021.500.506
INTRODUCTION
The percentage of males to females in animals and human are equal (50:50) according to Mendelian heredity laws, this depends on the sperm that fertilizes an egg1,2. The percentage of both sexes in an inhabitance at birth defined as a secondary sex ratio, while the percentage of males to females at the conception was defined as the primary sex ratio3. Many factors are impacting the percentage of male to female in mammal offspring such as the age of parents, physiology condition of maternal body4, mother’s birth season5 and fertility6. Besides, environmental pollution, like destructive earthquakes can alter the sex ratio7. The primary sex-ratio may be affected by deviating in the mortality, moving and ability of the X or Y gametes to combine with an egg8. There are a lot of ways for determining the gender of offspring, the simplest and low cost one of these methods is the diet adjustment9. Cation concentrations in diets affect the sex ratio in sheep10. The percentage of the four essential minerals like Na, K, Ca and Mg in food before pregnancy can facilitate the obtaining of the desirable sex 11. The mineral diet content can alter the pH level in the body, which changes the egg polarity or may influence the cervical mucus and the outer egg barrier permitting either (X) sperm or (Y) sperm to penetrate the ovum12-14. Sex pre-selection for offspring before conception to control the gender of progeny is desirable demand especially for the breeding program of farm animals. Sometimes there is a desire to increase the number of males to get meat or strength and sometimes it is the urge to get females to produce new individuals. In some cases, the principal medical indication for sex selection is to avoid giving progeny with a sex-linked genetic disorder.
The current study aimed to investigate the efficacy of putting monovalent ions (Na and K) and divalent ions (Ca and Mg) to the drinking water on the primary sex ratio, ovarian structure and serum mineral levels in New Zealand white rabbits does.
MATERIALS AND METHODS
Study area: This study was carried out at the laboratory of physiology and biotechnology, Department of animal production, Faculty of Agriculture, Mansoura University, Egypt, from October, 2018-May, 2019.
Animals: Nine New Zealand (NZW) white rabbit does with 5.4±0.61 months of age and 2.4±0.35 kg live body weight were used. For normal mating five fertile (NZW) buck’s rabbits, (7.5-8.1) month of age and (3.650-4.100) kg live body weight were used. All males and females were kept under optimum conditions of feeding and management, being separately housed in (40×50×60 cm) metal cages provided with food and water ad-libitum. Before the beginning, all animals were acclimatized for 5 days as an introductory period.
Experimental design: The rabbit does were divided into three groups, each one contains three animals. The first group was given water without any additives considered as control. The second group was given drinking water 1% of calcium and magnesium (Ca+Mg), while the third group was given drinking water of 1% of sodium and potassium (Na+K) for 15 days pre-mating. After 15 days as the treatment period, all does were naturally mated with fertile bucks (buck per 2 does within each group without treatment).
Mineral analysis: Blood samples were weekly collected from all doe pre and post-mating for measuring the concentrations of Ca, Mg, Na and K according to Horowitz and Latimer15.
Embryo collection: After 72 hrs post-coitus, all doe rabbits were sacrificed and the embryos were extracted from the reproductive tract by flushing with (phosphate-buffered saline supplemented with CaCl2 (DPBS, Sigma D5773), 0.002 gm mL–1 bovine serum albumin (BSA, Sigma A3311), 100 IU mL–1 penicillin and 100 μg mL–1 streptomycin) (RM) at room temperature (20-25°C). The extracted embryos were morphologically examined to detect the normal ones (morulae with both intact and regular mucin coat and zona pellucida and homogenous cell mass). Normal embryos from each doe were washed twice in fresh RM and kept at room temperature until use (10-15 min). Ovulation rate (number of luteinized follicles with ovulation stigmas), number of small, large and bleeding follicles and the number of normal and abnormal embryos were recorded as reproductive traits.
Determination of the sex of embryos
DNA extraction: Embryos were washed in PBS medium and placed in a 50 μL medium micro-drop inside a 90 mm diameter Petri dish, under an inverted microscope. Extraction of DNA from rabbit embryo using NP40/PK protocol according to Dehkordi et al.16. Cell lysis buffer was composed of 2.5 μL distilled water, 2.5 μL PCR buffer and 5 μL NP40/PK solution. The NP40/PK solution was composed of 48 μL distilled water, 1 μL NP40 (10%) and 1 μL PK (Proteinase K). Each embryo was then transferred in a microtube containing 10 μL of DNA lysis buffer. A mixture of embryo sample and cell lysis buffer were heated at 50°C for 60 min in a water bath and then heated at 96°C for 15 min in DNA thermocycler (Tc-512) to inactivate Proteinase K.
Polymerase Chain Reaction (PCR): The presence of SRY (sex-determining region Y gene) sequence is evidence, the sex is male. While in its absence, animal sex is female. Primers for PCR were designed based on sequence declare in Gene Bank (AF230075)17. Specific primers were designed with the program CLC main workbench. (SRY FOR 5'-AGC GGC CAG GAA CGG GTC AAG-3', SRY REV: 5'-CCT TCC GGC GAG GTC TGT ACT TG-3') (Bio Basic Inc., Sigma Company-Canada).
Forward primer: About 91 μL of buffer was added for the forward primer to make a 100 μM stock solution.
Reserve primer: About 99 μL of buffer was added for the reverse primer. The concentration of primer was measured by spectrophotometer (Parken Elmar- USA) with Optical Density at 260-280 nm.
PCR master mixture contained 2.5 μL dH2O, 2.5 μL dNTPs (0.2 mM), 1.5 μL MgCL2, 1.25 μL forward primer, 1.25 μL reverse primer, 1μL Taq Polymerase, 15 μL DNA lysis from the previous step. After cell lysis, 15 μL of PCR master mix was added to the microtube in the previous step for PCR analysis. PCR amplify the DNA fragments, The PCR cycles were 38 and temperature grade was: 94°C 5 min., denaturation 94°C 1 min, annealing at 65°C 1 min, extension 72°C 1 min and last step 72°C 10 min. The products of PCR were seen in 2% agarose gel.
Preparation of agarose gel: Two gm agarose (Fermentus Company-USA) was dissolved in 100 mL TAE buffer (Trace Acetate EDTA buffer) cooked in the microwave for 2 min. The amplified products were then electrophoresed for 45 min (Electrophoresis device from biorade-USA volt 80v/40 ampere) on a 2% agarose gel, stained with ethidium bromide and evaluated using ultraviolet light. The embryos were scored if it's positive or negative for the Y chromosome. The positive is male, while the negative is female. SRY analysis depends on the detection of fragments of 294 base pair length in males. While in females the fragment, not present.
Statistical analysis: The statistical analysis was performed as a one-way analysis of variance to test the differences among 3 groups. The statistical analysis was performed by a software package of the General Linear Model procedures18. The significant differences were set at the 0.05 level according to Duncan19.
RESULTS
Primary sex ratio: The sex ratio of embryos recovered from rabbits does in different experimental groups 72 hrs post-mating and their sex was detected using PCR is shown in Table 1 and Fig. 1. The present results showed that doe rabbits treated with Ca+Mg produced more female embryos (72.2%) versus 56% in the control group. While the rabbit does treat with Na+K produced more males embryos (69.7%) versus 44% produced in the control group (Table 1).
Effect of mineral treatment on the ovulatory response: The results showed that the administration of Ca+Mg (G2) or Na+K (G3) lead to a significant (p<0.05) increase in the number of corpora lutea, bleeding follicles and total embryos. While the number of large follicles decreased significantly compared to those in the control group (p<0.05). However, the number of small follicles and total follicles showed an insignificant increase (Table 2). Also, the data presented in Table 2 showed a significant increase (p<0.05) in ovulation rates of rabbits does in both treated groups (G2 and G3) as compared to the control one (75.6 and 77.4% vs. 60.8%, respectively. However, the recovery rate showed insignificant change among experimental groups, being 89.3, 90.0 and 86.8% for rabbit does in G1, G2 and G3, respectively (Table 2).
Effect of mineral treatment on serum Ca, Mg and Na levels: The present results indicated that the concentration of Ca was ranged between 8.742-9.766 mg dL–1 in the control group and 8.30-11.90 mg dL–1 in Ca+Mg treated rabbits group without significant changes in both groups throughout the experiment period (Table 3). However, the results showed in Table 4 indicated that Mg concentration in the control group ranged from 4.9-5.4 mg dL–1 without significant change throughout the experiment. On the other hand, Ca+Mg treated animals showed a pronounced increase of Mg by about 31.9% (p<0.05) at the end of the treatment to reach 6.467 as compared to 4.900 mg dL–1 at the start of the treatment period.
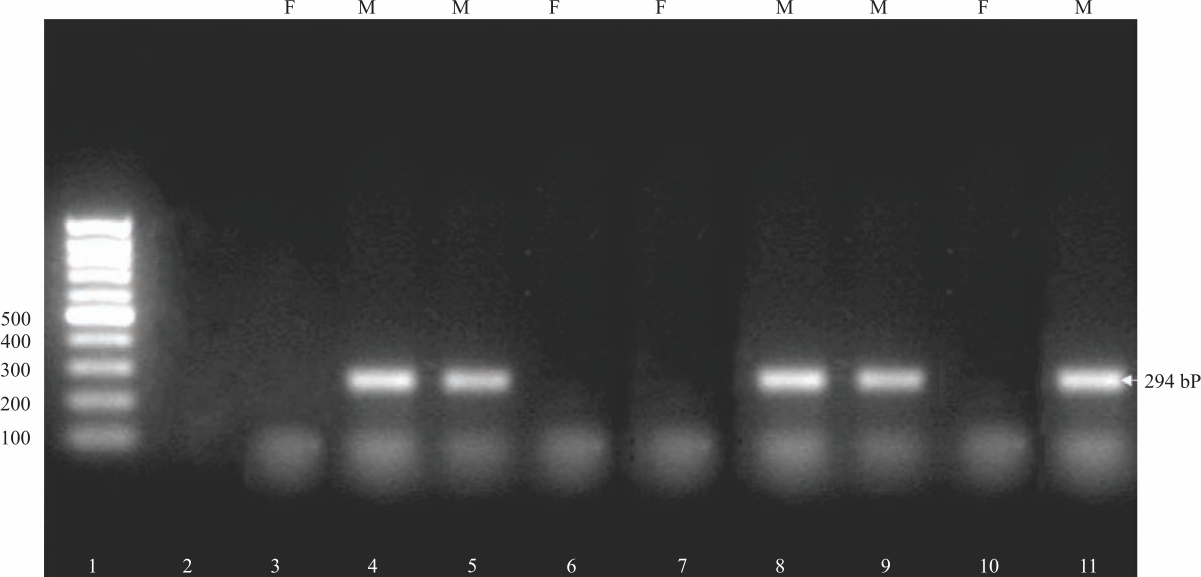 |
| Fig. 1: | Sex determination of rabbit embryos using SRY PCR assay Positive band at 294 bp is male. Lane 1 Molecular size marker, lane 2 is a negative control, lanes 4,5,8,9 and 11 are males (M). Lanes 3., 6, 7 and 10 are females (F) |
| Table 1: 1ry Sex ratio of recovered embryos from rabbit does (n = 3) in different experimental groups | |||||
| PCR sexing | |||||
| Male embryos | Female embryos | ||||
| Experimental group | Recovered embryos (N) | N | % | N | % |
| G1 (Control) | 25 | 11 | 44.0b | 14 | 56.0b |
| G2 (Ca+Mg) | 36 | 10 | 27.8c | 26 | 72.2a |
| G3 (Na+K) | 33 | 23 | 69.7a | 10 | 30.3c |
Upper letters (a, b and c) in the same column indicate a significant difference at p<0.05 | |||||
| Table 2: Effect of mineral treatment on ovarian structures and the ovulatory response of rabbits doe at different times of the treatment period (mean±SEM) | ||||
| Experimental group | ||||
| Item | G1 (Control) | G2 (Ca+Mg) | G3 (Na+K) | ±SEM |
| Average number of ovarian structures/doe | ||||
| Corpora lutea (CL) | 9.3c | 13.3a | 12.7b | 0.24 |
| Large follicles (LF) | 6.0a | 4.3b | 3.7b | 0.074 |
| Small follicles (SF) | 13.0 | 16.3 | 15.0 | 1.20 |
| Bleeding follicles (BF) | 0.33b | 1.0a | 1.30a | 0.04 |
| Total follicles (TF) | 19.3 | 21.6 | 20.0 | 1.31 |
| Total embryos (TE) | 8.3b | 12.0a | 11.0a | 0.31 |
| Ovulatory response (%) | ||||
| Ovulation rate (OR)* | 60.8b | 75.6a | 77.4a | 1.87 |
| Recovery rate (RR)** | 89.3 | 90.0 | 86.8 | 1.43 |
Upper letters (a, b and c) in the same row indicate a significant difference at p<0.05, * OR: Number of CL/number of LF+CLx100, **RR: Number of embryos/number of CLx100 | ||||
| Table 3: Calcium concentration in blood serum of control group and Ca+Mg treated rabbits during the treatment period (Mean±SEM) | ||||
| Concentration of Ca (mg dL–1) | ||||
| Experimental group | 0 time | 2 week | 4 week | ±SEM |
| G1 (Control) | 9.766 | 8.742 | 8.921 | 1.12 |
| G2 (Ca+Mg) | 10.1 | 8.30 | 11.90 | 1.87 |
| Table 4: Concentration of magnesium in blood serum of rabbits doe treated with Ca+Mg and control groups during the treatment period (±SEM) | ||||
| Concentration of Mg (mg dL–1) | ||||
| Experimental group | 0 time | 2 week | 4 week | ±SEM |
| G1 (Control) | 5.424 | 4.933 | 5.046 | 0.315 |
| G2 (Ca+Mg) | 4.900b | 4.800b | 6.467a | 0.344 |
| Upper letters (a, b and c) in the same row indicate a significant difference at p<0.05 | ||||
| Table 5: Concentration of sodium in blood serum of rabbits doe treat with Na+K and control groups during the treatment period (±SEM) | ||||
| Concentration of Na (mg dL–1) | ||||
| Experimental group | 0 time | 2 week | 4 week | ±SEM |
| G1 (Control) | 576.33 | 506.33 | 459.33 | 24.13 |
| G3 (Na+K) | 508.33 | 523.67 | 632.67 | 20.34 |
Besides, the data presented in Table 5 exhibited a gradual insignificant decrease in Na concentration in the control group during the treatment period. While there was an insignificant increase in those treated with Na+K.
DISCUSSION
In the present study, the male fetuses were identified after 72 hrs, post-mating by detection of 294 base pair length fragments using SRY PCR assay as a speed, sensitive and accurate tool whereas the absence of these fragments revealed to the female fetuses. Which cells of males contain X and Y chromosomes while females cell contain two X chromosomes. The Y-chromosome contains one gene named the SRY region (sex-determining region) that responsible for sex determination in mammals17,20. The determination of primary (1ry) sex ratio is associated with many factors such as parental age, environmental pollution and season21. Also, the mother's diet could impact the sex ratio of the progeny22,23. The present study showed that rabbits doe treated with Na+K produced more males (69.7%). While doe rabbits treated with Ca+Mg produced more female embryos (72.2%). These results are consistent with the previous studies which reported that administration of (Na and K) and (Ca and Mg) resulted in (56.80% male and 43.19% female), (39.20% male and 60.79% female) in rats and rabbits respectively9,24-26. Also, Chandraju et al.27 reported that rabbits take food contain (Na and K) gives more number of males than rabbits take normal food. Likewise, in humans, the diet has relative excess sodium and potassium ions would favor the birth of males, while diets with excess Ca and Mg ions would favor the birth of female28. The ability of an oocyte to attract either X or Y sperm depends on the balance between (Na and K) versus (Ca and Mg) that could alter the receptors of the oocyte wall. The attraction of oocyst to Y-sperm is associated with high Na and K intake than Ca and Mg11. The embryonic development of one sex more than the other may depend on changes in the reproductive tract PH and the ratio of ions in the ovaries and eggs as a result of the effect of Ca and Mg concentration29. While the change in pH of the follicular fluid from acidic to basic might have selectively attracted negatively charged X chromosome bearing spermatozoa30. Other studies reported that the pH of the vagina at the time of mating can affect the sex ratio, whereas sperm motility, life span and fertilizing capacity could be influenced by pH changes31. In this respect, Noorlander et al.23 showed that the diet constituents can promote ionic and/or hormonal changes that interact either through the cervical mucus with spermatozoa or at the oocyte level.
The present results showed an increase in Ca, Mg, Na levels after administration of drinking water supplemented with these minerals. These results run parallel with the data obtained by Edessy et al.28. However, these elevations were within the normal range of Ca, Mg and Na rabbit serum levels32,33. This means that the used dose of (Mg and Ca) was safe for rabbits.
The current study showed that increase in the number of corpora lutea, small follicles and total follicles in either (Ca and Mg) or (Na and K) treated groups. This is agreed with the previous studies which reported minerals to have a beneficial role in the resumption of follicular growth, ovulation and fertility through stimulation of the hypothalamus34-36. These results reflected the increase of the ovarian activity which Na intake was associated with increasing the follicle-stimulating hormone and luteinizing hormone. FSH is responsible for the development and maturation of ovarian follicles, increases estradiol hormone production37. As well as Kim et al.38 revealed that sodium and manganese may influence reproductive hormones and ovarian function in normally menstruating women. The present study showed that the recovery rate did not differ significantly among the studied groups, being 89.3, 90.0 and 86.8% for rabbits does in G1, G2 and G3, respectively. These results run parallel with results obtained by Kim et al.39. So we recommend adding the monovalent and divalent ions to the drinking water or altering the mineral percentage in the diets before conception for modulating the sex ratio in rabbits as a cheap, easy and safe method. These findings will help breeders manage their breeding, technicians and researchers to improve the reproductive performance of the farm animals. Further studies are required to determine the safe and optimal healthy doses of these minerals for sex preselection of the different farm animal species. Also, minerals administration could be used for human sex preselection.
CONCLUSION
The administration of (Na and K) increases the probability of male production while Ca and Mg administration increases the probability of female production. Also, the administration of these minerals increases the ovarian activity of the studied animals.
SIGNIFICANCE STATEMENT
This study showed the possible synergistic effect of administration of monovalent (Na and K) or divalent (Ca and Mg) ions for sex pre-selection of offspring before preconception. It is a simple and low-cost method to control the gender of progeny in New Zealand white rabbits does without adverse effects. This study will help the researcher and persons who are interested in genetic improvement and breeding farm animals to preselect the desirable sex of offspring according to their requirements.
REFERENCES
- Jongbloet, P.H., 2004. Over-ripeness ovopathy: A challenging hypothesis for sex ratio modulation. Human Reprod., 19: 769-774.
CrossRefDirect Link - Li, W.M., Y.P. Feng, R.X. Zhao, Y.Z. Fan and N.A. Affara et al., 2008. Sex ratio bias in early-dead embryos of chickens collected during the first week of incubation. Poult. Sci., 87: 2231-2233.
CrossRefPubMedDirect Link - Amadesi, A., A. Frana, L.M. Gandini, V. Bornaghi and K. Parati et al., 2015. Comparison between primary sex ratio in spermatozoa of bulls and secondary sex ratio in the deriving offspring. Theriogenology, 83: 199-205.
CrossRefDirect Link - Landete-Castillejos, T., A. Garcia, S. Langton, I. Inglis, L. Gallego and J. Garde, 2001. Opposing offspring sex ratio variations with increasing age and weight in mouflon mothers (Ovis musimon). Acta Vet. Hung., 49: 257-268.
CrossRefDirect Link - Nonaka, K., B. Desjardins, H. Charbonneau, J. Legare and T. Miura, 1999. Human sex ratio at birth and mother's birth season: Multivariate analysis. Human Biol., 71: 875-884.
Direct Link - Maestripieri, D., 2001. Female-biased maternal investment in rhesus macaques. Folia Primatol., 72: 44-47.
CrossRefDirect Link - Cagnacci, A., A. Renzi, S. Arangino, C. Alessandrini and A. Volpe, 2004. Influences of maternal weight on secondary sex ratio of human offspring. Hum. Rep., 19: 442-444.
CrossRef - Weir, J., 1962. Hereditary and environmental influences on the sex ratio of PHH and PHL mice. Genetics, 47: 881-897.
Direct Link - Oun, A.E., S. Bakry, S. Soltan, A. Taha and E. Kadry, 2016. Preconceptional minerals administration skewed sex ratio in rat offspring. Res. Obstet. Gynecol., 4: 11-15.
CrossRefDirect Link - Alhimaidi, A.R., A.A. Ammari, M.Q. Alghadi, M.Y. Al Saiady, R.A. Amran and A.A. Swelum, 2021. Sex preselection of sheep embryo by altering the minerals of maternal nutrition. Saudi J. Biol. Sci., 28: 680-684.
CrossRefDirect Link - Vahidi, A.R. and M.H. Sheikhha, 2007. Comparing the effects of sodium and potassium diet with calcium and magnesium diet on sex ratio of rats` offspring. Pak. J. Nutr., 6: 44-48.
CrossRefDirect Link - Stolkowski, J. and J. Lorrain, 1980. Preconceptional selection of fetal sex. Int. J. Gynecol. Obstet., 18: 440-443.
CrossRefDirect Link - Meikle, D.B. and M.W. Thornton, 1995. Premating and gestational effects of maternal nutrition on secondary sex ratio in house mice. Reproduction, 105: 193-196.
CrossRefDirect Link - Celik, K., S. Serbest, S. Vurur, A. Pala and K. Daglioglu, 2003. Experiments to investigate the factors that affect the rate of sex constitution. Pak. J. Nutr., 2: 238-241.
CrossRefDirect Link - AOAC, Horwitz, W. and G.W. Latimer, 2006. Official Methods of Analysis of AOAC International. 18th Edn., AOAC International, Gaithersburg, Maryland, ISBN-13: 9780935584776.
Direct Link - Dehkordi, F.F., A.M. Ahadi, A. Shirazi and M. Sadeghizade, 2009. Effect of lysis strategy in accuracy and repeatability of sex determination by single cell polymerase chain reaction method. J. Biol. Sci., 9: 78-82.
CrossRefDirect Link - Wallner, B., S. Huber and R. Achmann, 2001. Non-invasive PCR sexing of rabbits (Oryctolagus cuniculus) and hares (Lepus europaeus). Mamm. Biol.-Zeitschrift für Saugetierkunde, 66: 190-192.
Direct Link - SAS., 2004. The GLM Procedure. In: SAS/STAT® 9.1 User’s Guide. SAS Institute Inc., Cary, NC, USA, pp: 1731-1907.
Direct Link - Koopman, P., J. Gubbay, N. Vivian, P. Goodfellow and R. Lovell-Badge, 1991. Male development of chromosomally female mice transgenic for Sry. Nature, 351: 117-121.
CrossRefDirect Link - Mudie, N.Y., B.I. Gusev, L.M. Pivina, M.J. Schoemaker, O.N. Rijinkova, K.N. Apsalikov and A.J. Swerdlow, 2007. Sex ratio in the offspring of parents with chronic radiation exposure from nuclear testing in Kazakhstan. Radiat. Res., 168: 600-607.
CrossRefDirect Link - Rosenfeld, C.S. and R.M. Roberts, 2004. Maternal diet and other factors affecting offspring sex ratio: A Review. Bio. Rep., 71: 1063-1070.
CrossRefDirect Link - Noorlander, A.M., J.P.M. Geraedts and J.B.M. Melissen, 2010. Female gender pre-selection by maternal diet in combination with timing of sexual intercourse – a prospective study. Reprod. Biomed. Online, 21: 794-802.
CrossRefDirect Link - Shelan, S.S. and A.S. Alilawy, 2015. Embryonic sexing in rabbits by using special diet and change of vaginal pH. Intl. J. Sci. Res., 6: 2330-2333.
CrossRefDirect Link - Chandraju, S., A. Beirami and C.C. Kumar, 2013. Impact of sodium and potassium ions in identification of offspring gender in hamsters. Int. J. Pharm. Sci. Res., 4: 1529-1533.
Direct Link - Chandraju, S., A. Beirami and C.S.C. Kumar, 2012. Impact of calcium and magnesium ions in identification of offspring gender in high-Sugar rabbits. J. Chem. Pharm. Res., 4: 719-725.
Direct Link - Chandraju, S., A. Beirami and C.S.C. Kumar, 2014. Impact of sodium and potassium ions in identification of second generation of offspring gender in rabbits. Am. J. Phytomed. Clin. Ther., 2: 24-32.
Direct Link - Edessy, M., M.I. El-Rashedy, K. El-Batal, S. Ahmed, M. Badawy, A.E. Oun and A. Bendary, 2016. Pre-conceptional maternal diet and fetal sex pre-selection. Int. J. Curr. Res. Med. Sci., 3: 8-13.
CrossRefDirect Link - Farhadi, N., S. Khatamsaz, M. Shojaeifard, H. Kargar-Jahromi and Z. Khabbaz, 2014. Effects of administration of hydro-alcoholic extract of ginger (Zingiber officinale) on blood serum cations (Mg, Na+, K+, Ca2+) and the sex ratio of male and female newborn Wistar rats. J. Biol. Today's World, 3: 157-161.
CrossRefDirect Link - Grant, V.J. and L.W. Chamley, 2010. Can mammalian mothers influence the sex of their offspring peri-conceptually? Reproduction, 140: 425-433.
CrossRefDirect Link - Pratt, N.C., U.W. Huck and R.D. Lisk, 1987. Offspring sex ratio in hamsters is correlated with Vaginal pH at certain times of mating. Behav. Neural. Biol., 48: 310-316.
CrossRefDirect Link - Jones, R.T., 1975. Normal values for some biochemical constituents in rabbits. Lab. Anim., 9: 143-147.
PubMed - Hassan, S.A. and K.M. Hassan, 2009. Supplementation on some blood parameters of karadi lambs 1-effect of alkali-treated or untreated barley straw. Diyala J. Agric. Sci., 1: 1-12.
Direct Link - Brantmeier, S.A., R.R. Grummer and R.L. Ax, 1987. Concentrations of high density lipoproteins vary among follicular sizes in the bovine. J. Dairy. Sci., 70: 2145-2149.
CrossRefDirect Link - Murawski, M., G. Bydlon, K. Sawicka-Kapusta, E. Wierzchos and M. Zakrzewska et al., 2006. The effect of long term exposure to copper on physiological condition and reproduction of sheep. Reprod. Biol., 6: 201-206.
PubMedDirect Link - Ahuja, A.K. and D. Parmar, 2017. Role of minerals in reproductive health of dairy cattle: a review. Int. J. Livest. Res., 7: 16-26.
CrossRefDirect Link - Kim, B., E.S. Kang, M. Fava, D. Mischoulon and D. Soskin et al., 2013. Follicle-stimulating hormone (FSH), current suicidal ideation and attempt in female patients with major depressive disorder. Psychiatry Res., 210: 951-956.
CrossRefDirect Link - Kim, K., J. Wactawski-Wende, K.A. Michels, K.C. Schliep, T.C. Plowden, E.N. Chaljub and S.L. Mumford, 2018. Dietary minerals, reproductive hormone levels and sporadic anovulation: Associations in healthy women with regular menstrual cycles. Br. J. Nutr., 120: 81-89.
CrossRefDirect Link - Kim, M.J., H.J. Oh, J.E. Park, G.A. Kim, E.J. Park, G. Jang and B.C. Lee, 2012. Effects of mineral supplements on ovulation and maturation of dog oocytes. Theriogenology, 78: 110-115.
CrossRefDirect Link