
Lamya A. Al-Keridis
Department of Biology, College of Sciences, Princess Nourah Bint Abdulrahman University, 11474 Riyadh, Saudi Arabia
LiveDNA: 966.31394
Karem Ghoneim
Department of Zoology and Entomology, Faculty of Science, Al-Azhar University, Cairo, Egypt
Pakistan Journal of Biological Sciences
Year: 2021 | Volume: 24 | Issue: 1 | Page No.: 42-52







ABSTRACT
Background and Objective: The worldwide fly species, Muscina stabulans (Diptera: Muscidae) is known as 'false stable fly'. It has veterinary, forensic and medical importance. The present study aimed to examine the toxicity of novaluron (chitin synthesis inhibitor) via its effect on the growth and reproductive potential of M. stabulans. Materials and Methods: The early last (3rd) instar larvae and prepupae have been treated with novaluron using five doses: 5.0, 1.0, 0.5, 0.1 and 0.01 μg/larva. Student's t-test analysis has been used for data processing as well as refined by Bessel correction for significant differences among means. Results: Current study revealed that, after the treatment different mortalities of larva, pupa and adult have been estimated. LD50 values of novaluron were 0.018 and 0.057 μg/insect, respectively. Furthermore, the larval period was insignificantly shortened while the pupal duration has been significantly extended and the developmental rate has been slightly enhanced. On the other hand, the adult longevity of females was considerably reduced and the adult emergence was considerably blocked, regardless the dose. However, only after the treatment, some deformed pupae were observed and some adult deformities were observed. Both fecundity and fertility were drastically reduced and sterilizing activity on novaluron increased in a dose-dependent course, regardless the time of treatment. Conclusion: Novaluron can be used as an effective IGR in the integrated control program for this medically and veterinary serious fly.
PDF Abstract XML References Citation
Received: May 17, 2020;
Accepted: August 18, 2020;
Published: December 15, 2020
Copyright: © 2021. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Lamya A. Al-Keridis and Karem Ghoneim, 2021. Evaluation of the Toxic Effects of Novaluron on Muscina stabulans (Fallen) (Diptera: Muscidae). Pakistan Journal of Biological Sciences, 24: 42-52.
DOI: 10.3923/pjbs.2021.42.52
URL: https://scialert.net/abstract/?doi=pjbs.2021.42.52
DOI: 10.3923/pjbs.2021.42.52
URL: https://scialert.net/abstract/?doi=pjbs.2021.42.52
INTRODUCTION
In flies, family Muscidae includes a small genus Muscina which currently comprises 14 valid species and 30 additional species1. Muscina stabulans (Diptera: Muscidae) is commonly known as 'false stable fly'. It is worldwide fly species2 that mostly found at urban and suburban areas3. Also, maggots are frequently found in animal excrement and decaying vegetable materials as well as in poultry houses 4. Also, it is reported to inhabit latrines and household wastes and other filthy habitats5. The fly is reported to cause myiasis in humans and animals6. Mian et al.7 isolated Salmonella enteritidis from the fly adults in California and Yoshida et al.8 isolated the trypanosomatid flagellate Herpetomonas mariadeanei (Protozoa) from this fly species. In addition, M. stabulans larvae are reported among flies of the forensic importance2,9-11. Despite the potential of adults as a vector of pathogens, larvae prey on the immature stages of other dipterous species and subsequently limit their abundance12. Thus, these larvae can play a role in the control of some Dipter13.
The intensive and discriminate uses of many broad-spectrum conventional insecticides cause several serious problems, such as toxicological problems to humans, destruction of the biological enemies and the development of insect resistance toward different insecticides14-17. Therefore, alternative control agents have been searched recently to minimize insecticide hazards. Few years ago, a recent group of safe compounds had been synthesized. These compounds are known as Insect Growth Regulators (IGRs)18. IGRs are not directly toxic but act selectively on the growth, metamorphosis and/or reproduction of serious insect pests19,20 owing to their disruptive effects on the normal activity of endocrine system of insects21.
According to their modes of action, these Insect Growth Regulators (IGRs) were categorized in three groups: (i) Juvenile Hormone Analogues (JHAs), (ii) Ecdysteroids or ecdysone agonists and (iii) Chitin Synthesis Inhibitors (CSIs)22,23. CSIs were known as safe or low toxicity to the beneficial biota and non-target organisms with expected have no residual impact24 because they interfere with chitin biosynthesis and thus disrupt molting25.
In the late decades, some new benzoylphenylurea (BPU) compounds were synthesized, such as diflubenzuron, hexaflumuron, noviflumuron, lufenuron and novaluron26-28. Novaluron was reported to exhibit different effects on several dipterous species29-32. Also, its powerful suppression was recorded for lepidopterous larvae33-35 and whiteflies36,37. In addition, novaluron exhibited various effects on some coleopteran insects38-40. This compound has no or moderate impacts on parasitoids and other natural enemies33,36. Recently, novaluron was found to exhibit various effects on the pink bollworm Pectinophora gossypiella, such as reduced survival, retarded development, impaired metamorphosis41, disrupted adult performance, disturbed reproductive ability42 and declined main metabolites43. Therefore, the current study was conducted to assess the toxic effects of novaluron and its impacts on the development and the reproduction of M. stabulans.
MATERIALS AND METHODS
Experimental insect
Study area: A culture of susceptible strain of false stable fly Muscina stabulans (Fallen) (Diptera: Muscidae) was established in the laboratory of Entomology, Faculty of Science, Al-Azhar University, Cairo, Egypt. Controlled laboratory conditions (25±2°C, 55±5% R.H., photoperiod 12 L:12 D) were justified for a number of generations during the period 2017-2019. Larvae were fed on an artificial diet (200 g wheat bran, 100 g powdered milk, 3 g yeast and 200 mL tap water) based on the rearing technique of the house fly Musca domestica described by Busvine44. Feeding larvae were kept in breeding pans covered with muslin and fitted with rubber bands. After pupation (pupariation), pupae were gently collected and confined in Petri dishes. These Petri dishes were transferred into wooden cages (45×45×45 cm). Each cage was structurally provided with wire-gauze sides except the bottom and the front which was fitted with a small circular window attached with a cloth sleeve for routine daily work, such as feeding, handling and cleaning. After emergence, the adult flies were supplied with a milk diet and a piece of cotton soaked in 10-15% sugar solution.
Novaluron administration: Novaluron (C17H9ClF8N2O4) was purchased from Sigma-Aldrich Chemicals. A sequence of five dose levels has been formulated using acetone for novaluron dilutions (5.0, 1.0, 0.5, 0.1 and 0.01 μg/larva). Fifty larvae in 10 replicates (5 larvae/replicate) of the early last (3rd) instar and a similar number of prepupae were topically treated, individually, with each dose using a Hamilton micro applicator (NHN 737). A similar number of replicates of early last instar larvae and prepupae were topically treated with 1 μL acetone only as controls. All treated and control replicates were confined in small tubes and kept under the previously mentioned laboratory conditions. Treated and control insects were observed every day for feeding of the larvae and recording all criteria of the study.
Toxicity of novaluron: Toxicity of novaluron was determined by observed mortality. Mortality of larvae, pupae and adults (treated and control) have been noted every day and corrected as stated by Abbott’s formula45 as follows:
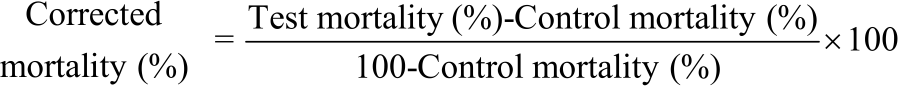 |
The values of LD50 for the tested compound, after treatment of larvae and prepupae, were calculated according to Finney46.
Developmental durations and rates: Developmental durations of larvae and pupae had been calculated (mean days±SD) using Dempster's equation47. The developmental rate was calculated according to Lotka48. Total longevity of adult female flies was also calculated in mean days±SD.
Metamorphosis and morphogenesis: Pupation rate has been calculated as % of the developed pupae and the adult emergence has been calculated in percentage as mentioned by Jimenez-Peydro et al.49 as follows:
 |
All of the possible aberrations of metamorphosis and morphogenesis were calculated in percentage.
Reproductive potential: For determination of the most important reproductive parameters, treated and control pupae were transferred into small cages (20×20×20 cm). Each treated female has been coupled with two normal males of the same age. Fecundity was calculated by the number of eggs per female. Fertility has been estimated by hatching the percentage of laid eggs. Sterility index has been evaluated as mentioned by Toppozada et al.50 as follows:
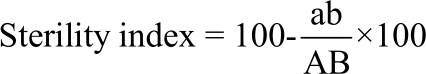 |
Where:
| a | = | Indicate the number of eggs laid per female in the treatment |
| b | = | Ratio of hatching in the treatment |
| A | = | Indicate the number of eggs laid per female in the controls |
| B | = | Percentage of hatching in the controls |
Data analysis: Data have been processed by the student's t-test and refined by Bessel correction51 for the test significant differences between means.
RESULTS
Toxic impact of novaluron against M. stabulans: Novaluron showed high toxicity that was expressed as mortality of larvae (maggots), pupae (puparia) and adult flies when early last (3rd) instar larvae have been treated as assorted in Table 1. Following topical treatment of prepupae, data of mortalities were arranged in Table 2. Last instar larvae treated with Novaluron showed high larval mortality in a dose-dependent course (10, 18, 28, 38 and 62% mortalities, at 0.01, 0.1. 0.5, 1.0 and 5.0 μg/larva, respectively, vs. 4% mortality among control larvae) as presented in Table 1. It was also observed that, pupae have completely died after the application of the highest two doses and the mortality of pupae was detected in a dose-dependent course, at other applied doses (22.2, 43.9 and 69.4% pupal mortality, at 0.01, 0.1 and 0.5 μg/larva, respectively, vs. control pupae mortality 2.1%).
Furthermore, pupae have completely died after the highest dose of Novaluron was applied for prepupae treatment as presented in Table 2. At other dose levels, the pupal mortality has been noted in a dose-dependent way (37.5, 47.6, 50.0 and 81.3% pupal mortality, at 0.01, 0.1, 0.5 and 1.0 μg/prepupa, respectively, vs. control pupae mortality, 4.1%). Depending on the data of the same table, no adult mortality could be recorded at the higher two doses of Novaluron because all pupae died. Following application of other three doses on last instar larvae, the adulticidal effect of Novaluron was exhibited parallel to the dose level, i.e., the mortality of adult has been noted in a dose-dependent manner (20.0, 47.8 and 63.6% adult mortality, at 0.01, 0.1 and 0.5 μg/larva, respectively, in comparison with 6.0% mortality of control adults). As clearly seen in Table 2, also, no adult mortality could be recorded after treatment of prepupae with the highest dose of novaluron because all pupae died. In addition, the adult mortality was recorded in a dose-dependent style (37.5, 47.6, 50.0 and 81.3% adult mortality, at 0.01, 0.1, 0.5 and 1.0 μg/prepupa, respectively, vs. control congeners mortality, 4.1%).
Depending on the corrected mortality percentages, Novaluron exerted a lethal potency against M. stabulans parallel to the dose level, regardless the time of treatment (Table 1 and 2). The calculated LD50 values of Novaluron against M. stabulans were 0.018 and 0.057 μg/insect for last instar larvae and prepupae, respectively. Therefore, the last instar larvae were more sensitive to the toxicity of Novaluron than prepupae.
| Table 1: Toxic effect (%) of Novaluron on M. stabulans after topical treatment of the early last instar larvae | ||||||
| Dose (μg/larva) | Larval mortality | Pupal mortality | Adult mortality | Total mortality | Corrected mortality | LD50 (μg/larva) |
| 5.0 | 62 | 100.0 | --- | 100 | 100.0 | |
| 1.0 | 38 | 100.0 | --- | 100 | 100.0 | |
| 0.5 | 28 | 69.4 | 63.6 | 92 | 90.9 | 0.018 |
| 0.1 | 18 | 43.9 | 47.8 | 76 | 72.7 | |
| 0.01 | 10 | 22.2 | 20.0 | 44 | 36.4 | |
| Control | 4 | 2.1 | 6.0 | 12 | --- | |
| -: No adult mortality could be calculated because no adult flies emerged | ||||||
Table 2: Toxic effect (%) of Novaluron on M. stabulans after topical treatment of the prepupae | |||||
| Dose (μg/prepupa) | Pupal mortality | Adult mortality | Total mortality | Corrected mortality | LD50 (μg/prepupa) |
| 5.0 | 100 | --- | 100 | 100.0 | 0.057 |
| 1.0 | 36 | 81.3 | 88 | 87.2 | |
| 0.5 | 24 | 50.0 | 62 | 59.6 | |
| 0.1 | 16 | 47.6 | 56 | 53.2 | |
| 0.01 | 4 | 37.5 | 40 | 36.2 | |
| Control | 2 | 4.1 | 06 | --- | |
| -: No adult mortality could be calculated because no adult flies emerged | |||||
| Table 3: Affected developmental durations (mean days±SD) of M. stabulans after topical application of Novaluron onto the early last instar larvae | |||||
Pupal stage | |||||
| Dose(μg/larva) | Larval duration | Duration | Developmental rate | Adult longevity | |
| 5.0 | 3.8±0.4a | --- | --- | --- | |
| 1.0 | 3.7±0.5a | --- | --- | --- | |
| 0.5 | 3.7±0.4a | 8.1±3.11b | 12.32 | 05.3±0.48d | |
| 0.1 | 3.8±0.2a | 8.0±1.84b | 12.50 | 08.3±0.94d | |
| 0.01 | 3.8±0.2a | 6.6±1.24a | 15.52 | 10.7±1.70d | |
| Control | 3.9±0.4 | 6.6±1.38 | 15.50 | 19.2±0.20 | |
| -: Some pupae died as morphologically normal pupae and other pupae died as deformed pupae, thus pupal duration could not be measured. Mean±SD followed with the same letter a: Insignificantly different (p>0.05), b: Significantly different (p<0.05), d: Very highly significantly different (p<0.001) | |||||
| Table 4: Affected developmental durations (mean days±SD) of M. stabulans after topical application of Novaluron onto the prepupae | |||||
Pupal stage | |||||
| Dose (μg/prepupa) | Duration | Developmental rate | Adult longevity | ||
| 5.0 | --- | --- | --- | ||
| 1.0 | 6.4±1.62a | 16.47 | 06.8±0.12d | ||
| 0.5 | 6. 5±1.74a | 16. 45 | 08.3±2.88d | ||
| 0.1 | 4.6±0.80b | 19.67 | 10.0±0.80d | ||
| 0.01 | 6.1±1.70a | 16.48 | 13.5±1.28d | ||
| Control | 6.6±4.60 | 16.40 | 19.7±1.50 | ||
| -: Some pupae died as morphologically normal pupae and other pupae died as deformed pupae, thus pupal duration could not be measured. Mean±SD followed with the same letter a: Insignificantly different (p>0.05), b: Significantly different (p<0.05), d: Very highly significantly different (p<0.001) | |||||
Effect of novaluron on the developmental durations and rates of M. stabulans: Data about the durations of different developmental stages of the last instar larvae treated with Novaluron were arranged in Table 3. Based on these data, the larval duration was insignificantly shortened, in no certain trend. In contrast, the pupal duration was remarkably prolonged at 0.5 and 0.1 μg/larva (8.1±3.11 and 8.0±1.84 days, respectively, vs. 6.6±1.38 days of control pupae). Also, the developmental rate of treated pupae had been seriously suppressed at these two doses. The adult female longevity was considerably shortened (5.3±0.48, 8.3±0.94 and 10.7±1.7 days at 0.5, 0.1 and 0.01 μg/larva respectively, in comparison to 19.2±0.2 days of control adult females.
Information about the effect of application of Novaluron onto the prepupae development was shown in Table 4. According to these data, the pupal duration has been significantly or insignificantly shortened, based on the dose concentration (6.4±1.62, 6.5±1.74, 4.6±0.8 and 6.1±1.70 days, at 1.0, 0.5, 0.1 and 0.01 μg/prepupa, respectively, vs. 6.6±4.6 days of control pupae).
| Table 5: Metamorphic and morphogenic effects of Novaluron on M. stabulans after topical treatment of early last instar larvae | |||||
Pupal stage | Adult stage | ||||
| Dose (μg/larva) | Pupation (%) | Pupal deformities (%)* | Emergence (%) | Adult deformities (%)** | |
| 5.0 | 39.0 | 32.0 | 00.0 | --- | |
| 1.0 | 61.3 | 19.8 | 00.0 | --- | |
| 0.5 | 71.6 | 30.6 | 87.0 | 06.4 | |
| 0.1 | 81.8 | 17.6 | 69.2 | 10.6 | |
| 0.01 | 89.9 | 11.4 | 93.3 | 05.0 | |
| Control | 100 | 00.0 | 93.3 | 00.0 | |
| *: Deformed pupae perished without metamorphosis into adult flies, **: Deformed adults with curly wings, atrophied mouth parts and ill-developed legs. These adult females perished within few days without mating | |||||
| Table 6: Metamorphic and morphogenic effects of Novaluron on M. stabulans after topical treatment of prepupae | |||||
Pupal stage | Adult stage | ||||
| Dose (μg/prepupa) | Pupation (%) | Pupal deformities (%)* | Emergence (%) | Adult deformities (%)** | |
| 5.0 | 90.0 | 0 | 00.0 | --- | |
| 1.0 | 100 | 0 | 51.6 | 30.2 | |
| 0.5 | 100 | 0 | 76.7 | 22.3 | |
| 0.1 | 100 | 0 | 83.3 | 16.1 | |
| 0.01 | 100 | 0 | 86.7 | 26.0 | |
| Control | 100 | 0 | 93.3 | 00.0 | |
| *: Deformed pupae perished without metamorphosis into adult flies, **: Deformed adults with curly wings, atrophied mouth parts and ill-developed legs. These adult females perished within few days without mating | |||||
The developmental rate of treated pupae was slightly elevated, especially at 0.1 μg/prepupa. Data of Table 4 obviously revealed that the adult longevity was conspicuously shortened, in a dose-dependent style, following prepupae topical treatment (13.5±1.28, 10.0±0.80, 8.3±2.88 and 6.8±0.12 days, at 0.01, 0.1, 0.5 and 1.0 μg/prepupa, respectively, vs. days of adult females control, 19.7±1.5).
Effects of novaluron on metamorphosis of M. stabulans: Following Novaluron application on the last instar larvae with, data of metamorphosis and morphogenesis were arranged in Table 5. Following current application of Novaluron on the prepupae, data of metamorphosis and morphogenesis were assorted in Table 6. As clearly seen in Table 5, the pupation rate (%) was regressed in a dose-dependent manner (89.9, 81.8, 71.6, 61.3 and 39% pupation, at 0.01, 0.1, 0.5, 1.0 and 5.0 μg/larva, respectively, vs. 93.3% pupation of control congeners). Additionally, data of Table 6 showed no effect of Novaluron on the pupation rate after the treatment of prepupae.
In respect of the adult emergence, no adults emerged following application of the highest two doses for the treatment of 3rd instar larvae. The adult emergence was remarkably blocked at the other three doses (93.3, 69.2 and 87.0%, at 0.01, 0.1 and 0.5 μg/larva, respectively, vs. 93.3% emergence of control adults, Table 5). Following treatment of prepupae, the adult eclosion was completely blocked at the highest dose and increasingly blocked parallel to the increasing dose (86.7, 83.3, 76.7 and 51.6% emergence, at 0.01, 0.1, 0.5 and 1.0 μg/prepupa, respectively, vs. 93.3% emergence of control adults, Table 6).
Effect of novaluron on morphogenesis of M. stabulans: Following Novaluron application for the treatment of last instar larvae, disruptive effect on the pupal morphogenesis was observed, since different percentages of deformed pupae were recorded, with no certain trend (11.4, 17.6, 30.6, 19.8 and 32.0% pupal deformities, at 0.01, 0.1, 0.5, 1.0 and 5.0 μg/larva, respectively, vs. 0% deformity of control pupae, Table 5). These deformed pupae perished without metamorphosis into adults. On the contrary, Novaluron failed to affect the pupal morphogenesis after topical treatment of prepupae, since no deformed pupae were observed (Table 6).
With regard to the affected adult morphogenesis, treated early last instar larvae with Novaluron showed some deformities of adults, in no certain trend (5.0, 10.6 and 6.4% deformed adults, at 0.01, 0.1 and 0.5 μg/larva, respectively, in comparison to 0% deformation of control adults, Table 5). Also, various percentages of malformed adults were recorded after topical treatment of prepupae with Novaluron, in no certain trend (26, 16.1, 22.3 and 30.2% deformed adults, at 0.01, 0.1, 0.5 and 1.0 μg/prepupa, respectively, in comparison to 0% deformation of control adults, Table 6). Irrespective of the time of treatment with Novaluron, the deformed adults could not sexually be differentiated (males or females). The major features of deformation had been seen in dwarf bodies with curly wings, atrophied mouthparts and ill-developed legs. All deformed adults perished within few days without mating.
| Table 7: Reducing effect of Novaluron on the reproductive potential of M. stabulans | ||||||
After treatment of last instar larvae | After treatment of prepupae | |||||
| Dose (μg/larva) | Fecundity (mean No. of egg±SD) | Fertility (hatching %) | Sterility index | Fecundity (mean No. of egg±SD) | Fertility (hatching %) | Sterility index |
| 5.0 | --- | --- | --- | --- | --- | --- |
| 1.0 | --- | --- | --- | 073.5±1.5d | 13.3 | 94.99 |
| 0.5 | 140.3±0.9d | 76.8 | 27.12 | 135.8±1.6d | 77.2 | 37.09 |
| 0.1 | 151.4±1.2d | 82.4 | 13.38 | 146.8±1.7d | 81.0 | 28.76 |
| 0.01 | 153.0±1.3d | 88.1 | 07.77 | 152.7±1.2d | 85.0 | 25.58 |
| Control | 192.3±1.8 | 98.5 | - | 194.8±2.5 | 100 | - |
| d: Very highly significantly different (p<0.001) | ||||||
Effect of novaluron on the reproductive capacity of M. stabulans: Important parameters of the reproductive capacity of M. stabulans have been presented in Table 7 as results of novaluron treatment on the early last instar larvae and prepupae. With regard to the fecundity (mean number of eggs/♀), novaluron exerted increasingly suppressive action on the oviposition females, regardless the time of treatment. In some detail, the female fecundity was drastically reduced after treatment of the early last instar larvae, in a dose-dependent fashion (153.0±1.3, 151.4±1.2 and 140.3±0.9 eggs/treated ♀, at 0.01, 0.1 and 0.5 μg/larva, respectively, compared to 192.3±1.8 eggs/control ♀). In a similar trend, fecundity was dramatically inhibited correspondingly to the dose level, after treatment of prepupae (152.7±1.2, 146.8±1.7, 135.8±1.6 and 073.5±1.5 eggs/treated ♀, at 0.01, 0.1, 0.5 and 1.0 μg/prepupa, respectively, in comparison to 194.8±2.5 eggs/control ♀).
In respect of the fertility (hatching% = hatchability), it was clearly indicated the inhibitory effect of Novaluron of the egg viability, regardless the time of treatment, since fertility was seriously reduced after treatment of last instar larvae, in a dose-dependent style (88.1, 82.4 and 76.8% reduction of fertility, at 0.01, 0.1 and 0.5 μg/larva, respectively, vs. 98.5% hatchability of control eggs) as noted in Table 7. Also, sterility index increased with the increasing dose level of Novaluron. Similarly, fertility was pronouncedly reduced after treatment of prepupae, in a dose-dependent manner (85.0, 81.0, 77.2 and 13.3% hatching of treated eggs, at 0.01, 0.1, 0.5 and 1.0 μg/prepupa, respectively, vs. 100% hatching of control eggs). In a comparable style, the sterility index increased (Table 7).
DISCUSSION
Findings from the present investigation indicated that the application of novaluron to the early last instar larvae of M. stabulans led to larval mortality in a dose-dependent manner.
Following application of the highest two doses against last instar larvae or treatment of prepupae with the highest dose, all developed pupae completely died. At other dose levels, pupal mortality has been noted in a dose-dependent style. Also, the mortality of adult has been noted in a dose-dependent manner after the treatment of larvae or prepupae with novaluron. Therefore, the present outcomes have been, to some extent, in accordance with some stated findings of toxicity of different IGRs against a number of insects, such as fenoxycarb against the parasitoid Phanerotoma ocularis52 and juvenoid pyriproxyfen against the desert locust Schistocerca gregaria53. Larvicidal and pupicidal impact of methoprene has been reported on the common house mosquito Culex pipiens54, the rice meal moth Corcyra cephalonica55, the yellow fever mosquito Aedes aegypti 56 and the flesh fly Sarcophaga ruficornis57. Also, toxicities of other IGRs were reported against different insects, such as kinoprene against C. pipiens58.
In the current investigation, larval mortalities of M. stabulans might be due to the prevention of larvae to split the old cuticle and expand the new one during moulting59. Additionally, the mortality of the larva might have resulted from continuous starvation60. In addition, pupal mortality might be directly due to failure of vital homeostatic mechanisms and/or some secondary factors, such as bleeding, suffocation and desiccation by imperfect exuvation61.
On the other hand, in insects, however, LC50 of an agent differs according to various factors, such as its lethal potency as well as its concentration, the sensitivity of the insect and its developmental stage or instar, method and time of treatment and the experimental conditions. In this context, the early last instar larvae of M. stabulans were more sensitive to novaluron than prepupae, in the present study. This finding coincides, to a great extent, with many works reporting that the early larval instars of different flies have been more sensitive than the later ones to some IGRs, such as Ceratitis62.
In the present investigation on M. stabulans, prolonged pupal period and regressed developmental rate, following the treatment of last instar larvae with novaluron at different doses 0.5 and 0.1 μg/larva of, might be due to the disturbance of the neuroendocrine organs responsible for the synthesis and/or release of their hormones63. To explicate the shortened larval period after treatment of last instar larvae with novaluron and shortened pupal period after treatment of prepupae, this might be due to avoiding responses of these stages to novaluron as a xenobiotic agent.
In the current research, novaluron enforced the adult females of M. stabulans to survive short longevity. Current findings were, to some extent, in corroboration with many reports resulted in shortened longevity in different insects by several IGRs, such as S. litura by ecdysteroid RH-584964. Present findings, the shortened longevity of M. stabulans adult females, might be an indication to the interference of novaluron with the hormonal activity in adults, while a close relation between special hormones and adult longevity has been noted in other insects65,66.
After attainment of the sexual maturity in adult insects, different degenerative changes have been shown in some organs and tissues which could be collectively described as 'senility'. However, the disturbed adult longevity might be considered as a useful sign for the adult aging, i.e., shortened longevity might indicate the accelerated aging and vice versa. JH regulates the aging because of its ability for direct affects mechanisms of the somatic survival as reported by Yamamoto et al.67. Therefore, novaluron could influence the JH level and/or functions resulting in shortening of adult longevity, in the current investigations. Though, the particular mechanism of novaluron on the biochemical processes in adults still needs further investigation.
A dose-dependent regression of the pupation rate of M. stabulans, in the present study was recorded after treatment of last instar larvae with novaluron. In contrast, no affect was recorded on this rate after treatment of prepupae. These results were, to some extent, in agreement with some reported results in some insects by various IGRs which regressed the pupation rate68,21.
Depending on the present results the adult emergence of M. stabulans was completely prevented after treatment of early last instar larvae with the higher two doses of novaluron (5.0 and 1.0 μg/larva) and after treatment of prepupae with the highest dose. In addition, the adult emergence was remarkably blocked at the other doses by different percentages. These results were, to a great extent, in corroboration with some reported results since adult emergence in different insect species was partially or completely hindered after larval treatment with various LGRs, such as S. littoralis after treatment with novaluron35.
In this regard, it is critical to point out that the adult eclosion in insects is a vital process and regulated by the Eclosion Hormone (EH). Partial or complete disturbance of EH prohibit the adults to emerge69. The blocked adult emergence, in the present study, can be interpreted by a disturbing effect of novaluron on the adult EH release.
In the current study, a disruptive effect was exhibited by Novaluron on the pupal morphogenesis of M. stabulans, since different deformed pupae were observed after treatment of the last instar larvae. For interpretation of the anti-morphogenic activity of novaluron against pupae of M. stabulans, in the present study, it might exhibit suppressive effect on the chitin synthesis leading to prevent the normal deposition of new cuticle70.
In the current investigation, some adult deformations were observed after treatment of last instar larvae and prepupae of M. stabulans with novaluron. The production of anomalous M. stabulans adult flies might be interpreted by an adverse action of novaluron on the hormonal balance during the adult metamorphosis, since the disturbance of ecdysteroid titer leads to changed activity of the lysosomal enzyme.
In the present study the female fecundity of M. stabulans was drastically inhibited after novaluron application on last instar larvae or prepupae. In the current investigation, fecundity of M. stabulans was drastically prohibited which might resulted from the interference of novaluron with some reproductive developments, including the ovarian follicle growth and the egg maturation. In some detail, this can be explained by some reasons as follows:
| • | Some disorders in ovaries might be caused by novaluron71 |
| • | Novaluron might disrupt the synthesis of proteinaceous contents during the oogenesis72 |
| • | Novaluron might disturb the production and/or function of the gonadotropic hormone responsible for vitellogenesis (yolk precursors)73 |
| • | Eggs might develop normally in ovaries but they could not be lay, owing to the severely deformed ovipositor or to the reduced mechanical strength74 or their reabsorption before oviposition75 |
In the present study, the fertility of M. stabulans adult females was seriously reduced after treatment of last instar larvae or prepupae with novaluron. This result was in accordance with some noted outcomes of reduced fertility in different insects by some IGRs, such as L. dispar76 by methoxyfenozide.
In the current research, several recommendations might be given for explicating the fertility reduction in M. stabulans by novaluron, (1): Vitellogenins are necessarily required for the embryogenesis in the insect egg77. These vitellogenic metabolites are synthesized mainly by fat body during the developmental stages or inside the ovary itself78. Novaluron might disrupt the production and/or accumulation of these metabolites in adult females of M. stabulans, (2): Novaluron might disrupt the vitellogenin deposition into oocytes through the inhibition of gonadotropic hormone responsible for the egg maturation or/and exhibit a disruptive effect on the intracellular spaces in the follicular epithelium79, (3): From the treated M. stabulans immature stages, some residual amounts of novaluron penetrated into mothers and then into eggs. The residual material affected the cuticle synthesis resulting in weakened mouth parts of mature embryos to perforate the vitellin membrane for hatching42, (4): In the current study, reduction of M. stabulans fertility might be due to the negative influence at certain stages of novaluron on the developing embryos.
CONCLUSION
Depending on the present results, novaluron exhibited toxicity against larvae, pupae and adults of M. stabulans. Also, the tested compound exerted disruptive action on development, metamorphosis and morphogenesis of pupae and adults in addition to its reducing action on fecundity and sterilizing activity. Therefore, novaluron can be used as an effective IGR in the integrated control program for this medically and veterinary serious fly.
SIGNIFICANCE STATEMENT
This study discovered the tested compound, novaluron that can be beneficial for impairing the developmental and reproduction of M. stabulans. This study will help the researchers to uncover some critical areas of the control of the present medically dangerous fly that many researchers were not able to explore.
ACKNOWLEDGMENT
This research was funded by the Deanship of Scientific Research at Princess Nourah bint Abdulrahman University through the Fast-track Research Funding Program.
REFERENCES
- Grzywacz, A., M.J.R. Hall, T. Pape and K. Szpila, 2017. Muscidae (Diptera) of forensic importance-an identification key to third instar larvae of the western Palaearctic region and a catalogue of the muscid carrion community. Int. J. Legal Med., 131: 855-866.
CrossRefDirect Link - Patitucci, L.D., P.R. Mulieri, J.C. Mariluis and J.A. Schnack, 2010. The population ecology of Muscina stabulans (fallén) (diptera: muscidae), along an urban-rural gradient of buenos aires, Argentina Neotrop. Entomol., 39: 441-446.
CrossRefDirect Link - Smith, K.G.V., 1986. A Manual of Forensic Entomology. 1st Edn., British Museum (Natural History), London and Cornell University Press, Ithaca, Pages: 205.
Direct Link - Laos, F., L. Semenas and V. Labud, 2004. Factors related to the attraction of flies at a biosolids composting facility (Bariloche, Argentina). Sci. Total Environ., 328: 33-40.
CrossRefDirect Link - Zumpt, F., 1965. Myiasis in Man and Animals in the Old World: A textbook for Physicians, Veterinarians and Zoologists. Butterworths Publishing Company, London, Pages: 267.
CrossRefDirect Link - Mian, L.S., H. Maag and J.V. Tacal, 2002. Isolation of Salmonella from muscoid flies at commercial animal establishments in San Bernardino County, California. J. Vector Ecol., 27: 82-85.
Direct Link - Yoshida, N., E. Freymüller and F.G. Wallace, 2007. Herpetomonas mariadeanei sp. n. (Protozoa, Trypanosomatidae) from Muscina stabulans (Falléen, 1816) (Diptera, Muscidae). J. Protozool., 25: 421-425.
CrossRefDirect Link - Schroeder, H., H. Klotzbach and K Püschel, 2003. Insects colonization of human corpses in warm and cold season. Forensic Sci. Int., 5: 372-374.
CrossRefDirect Link - Gaudry, E., 2010. The Insects Colonisation of Buried Remains. Springer, Dordrecht, New York, ISBN: 978-1-4020-9684-6, pp: 273-311.
CrossRefDirect Link - Gunn, A. and J. Bird, 2011. The ability of the blowflies Calliphora vomitoria (Linnaeus), Calliphora vicina (Rob-Desvoidy) and Lucilia sericata (Meigen) (Diptera: Calliphoridae) and the muscid flies Muscina stabulans (Fallén) and Muscina prolapsa (Harris) (Diptera: Muscidae) to colonise buried remains. Forensic Sci. Int., 207: 198-204.
CrossRefDirect Link - Legner, E.F. and E.J. Dietrick, 1989. Coexistence of predatory Muscina stabulans andophyra aenescens [Dipt.: Muscidae] with dipterous prey in poultry manure. Entomophaga, 34: 453-461.
CrossRefDirect Link - Duarte, J.L.P, R.F. Kruger and P.B. Ribeiro, 2013. Interaction between Musca domestica L. and its predator Muscina stabulans (Fallen) (Diptera, Muscidae): Effects of prey density and food source abundance. Revista Brasileira Entomologia, 57: 55-58.
CrossRefDirect Link - Rose, R.I., 2001. Pesticides and public health: Integrated methods of mosquito management. Emerg. Infect. Dis., 7: 17-23.
CrossRef - Davies, T.G.E., L.M. Field, P.N.R. Usherwood and M.S. Williamson, 2007. DDT, pyrethrins, pyrethroids and insect sodium channels. IUBMB Life, 59: 151-162.
CrossRefDirect Link - Costa L.G., G. Giordano, M.Guizzetti and A. Vitalone, 2005. Neurotoxicity of pesticides: A brief review. Front. Biosci., 13: 1240-1249.
CrossRefPubMedDirect Link - Mosallanejad, H. and G. Smagghe, 2009. Biochemical mechanisms of methoxyfenozide resistance in the cotton leafworm Spodoptera littoralis. Pest. Manag. Sci., 65: 732-736.
CrossRefDirect Link - Khan, I. and A. Qamar, 2012. Andalin, an insect growth regulator, as reproductive inhibitor for the red cotton stainer, Dysdercus koenigii (F.) (Hemiptera: Pyrrhocoridae). Acad. J. Entomol., 5: 113-121.
CrossRefDirect Link - Nicholas, A.H., W.G. Thwaite and R.N. Spooner-Hart, 1999. Arthropod abundance in an Australian apple orchard under mating disruption and supplementary insecticide treatments for codling moth, Cydia pomonella (L.) (Lepidoptera: Tortricidae). Aust. J. Entomol., 38: 23-29.
CrossRefDirect Link - Martins, F. and I.G. da Silva, 2004. Evaluation of the inhibiting activity of the diflubenzuron on the ecdysis of larvae of Aedes aegypti (Linnaeus, 1762) (Diptera, Culicidae). Rev. Soc. Bras. Med. Trop., 37: 135-138.
CrossRefDirect Link - Wang, Q.L. and T.X. Liu, 2016. Effects of three insect growth regulators on Encarsia formosa (hymenoptera: aphelinidae), an endoparasitoid of Bemisia tabaci (hemiptera: aleyrodidae). J. Econ. Entomol., 109: 2290-2297.
CrossRefDirect Link - Dhadialla, T.S., G.R. Carlson and D.P. Le, 1998. New insecticides with ecdysteroidal and juvenile hormone activity. Ann. Rev. Entomol., 43: 545-569.
CrossRefDirect Link - Oberlander, H. and D. Silhacek, 2000. Insect Growth Regulators. In: Alternatives to Pesticides in Stored-Product IPM, Subramanyam, B. and D.W. Hagstrum (Ed.)., Kluwer Academic Publishers, Boston, ISBN: 9781461543534, pp: 147-163.
CrossRefDirect Link - Talikoti, L.S., D. Sridevi and T. Ratnasudhakar, 2012. Relative toxicity of insect growth regulators against tobacco caterpillar, Spodoptera litura (Fabricius). J. Entomol. Res., 36: 31-34.
Direct Link - Mondal, K.A.M.S.H and S. Parween, 2001. Insect growth regulators and their potential in the management of stored product pests. Integrated Pest Manage. Rev., 5: 255-295.
CrossRef - Kim, K.S., B.J. Chung and H.K. Kim, 2000. DBI-3204: A new benzoylphenyl urea insecticide with a particular activity against whitefly. Proceedings of an international conference held at the Brighton Hilton Metropole Hotel, Brighton, UK, November 13-16, 2000 CABI, pp: 41-46.
Direct Link - Karr, L.L., J.J. Sheets, J.E. King and J.E. Dripps, 2004. Laboratory performance and pharmacokinetics of the benzoylphenylurea noviflumuron in eastern subterranean termites (Isoptera: Rhinotermitidae). J. Econ. Entomol., 97: 593-600.
CrossRefDirect Link - Ishaaya, I., A. Barazani, S. Kontsedalov and R.A. Horowitz, 2007. Insecticides with novel modes of action: Mechanism, selectivity and cross-resistance. Entomol. Res., 37: 148-152.
CrossRefDirect Link - Bouaziz, A., H. Boudjelida and N. Soltani, 2011. Toxicity and perturbation of the metabolite contents by a chitin synthesis inhibitor in the mosquito larvae of Culiseta longiareolata. Annals Biol. Res., 2: 134-142.
Direct Link - Fontoura, N.G., D.F. Bellinato, D. Valle and J.B.P. Lima, 2012. The efficacy of a chitin synthesis inhibitor against field populations of organophosphate-resistant Aedes aegypti in Brazil. Mem. Inst. Oswaldo Cruz, 107: 387-395.
CrossRefDirect Link - Djeghader, N., H. Djeghader, A. Bouaziz and N. Soltani, 2013. Biological effects of a benzoylphenylurea derivative (Novaluron) on larvae of Culex pipiens (Diptera: Culicidae). Adv. Appl. Sci. Res., 4: 449-456.
Direct Link - Lohmeyer, K.H., J.M. Pound, K.M. Yeater and M.A. May, 2014. Efficacy of novaluron as a feed-through for control of immature horn flies, house flies and stable flies (Diptera: Muscidae) developing in cow manure. J. Med. Entomol., 51: 873-877.
CrossRefDirect Link - Ishaaya, I., S. Kontsedalov, D. Masirov and A.R. Horowitz, 2001. Bio-rational agents-mechanism, selectivity and importance in IPM programs for controlling agricultural pests. Med. Landbouww Rijksuniv Gent., 66: 363-374.
Direct Link - Murthy, K.S.R.K. and G.M. Ram, 2002. Studies on the efficacy of a new chitin synthesis inhibitor Rimon (novaluron 10 EC) on American bollworm Helicoverpa armigera Hubn. attacking cotton. In: Resources Management in Plant Protection During Twenty First Century, Vol. II, Hyderabad, India, pp: 165-168. https://www.cabdirect.org/cabdirect/abstract/20043048744
- Ishaaya, I., A.R. Horowitz, L. Tirry and A. Barazani, 2002. Novaluron (Rimon), a novel IGR: mechanism, selectivity and importance in IPM programs. Meded. Rijksuniv. Gent. Fak. Landbouwkd Toegep. Biol. Wet. 67: 617-626.
Direct Link - Ishaaya, I., S. Kontsedalov and A.R. Horowitz, 2003. Novaluron (Rimon), a novel IGR: Potency and cross-resistance. Arch. Insect Biochem. Physiol., 54: 157-164.
CrossRefDirect Link - Cutler, G.C., C.D. Scott-Dupree, J.H. Tolman and C.R. Harris, 2007. Field efficacy of novaluron for control of Colorado potato beetle (Coleoptera: Chrysomelidae) on potato. Crop Prot., 26: 760-767.
CrossRefDirect Link - Alyokhin, A., R. Guillemette and R. Choban, 2009. Stimulatory and suppressive effects of novaluron on the Colorado potato beetle reproduction. J. Econ. Entomol., 102: 2078-2083.
CrossRefDirect Link - Arthur, F.H. and E.A. Fontenot, 2012. Residual activity of methoprene and novaluron as surface treatments to manage the flour beetles, Tribolium castaneum and Tribolium confusum. J. Insect Sci., 12: 1-11.
CrossRefDirect Link - Ghoneim, K., H.A. Hassan, M.A. Tanani and N.A. Bakr, 2017. Toxic and disruptive effects of Novaluron, a chitin synthesis inhibitor, on development of the pink bollworm Pectinophora gossypiella (Saunders) (Lepidoptera: Gelechiidae). Int. J. Entomol. Res., 2: 36-47.
Direct Link - Hassan H.A., K. Ghoneim, M.A. Tanani and N.A. Bakr, 2017. Impaired adult performance and reproductive potential of the pink bollworm Pectinophora gossypiella (Saunders) (Lepidoptera: Gelechiidae) by the chitin synthesis inhibitor, Novaluron. J. Entomol. Zool. Stud., 5: 581-592.
Direct Link - Tanani, M.A., K. Ghoneim, H.A. Hassan and N.A. Bakr, 2017. Perturbation of main body metabolites in the pink bollworm Pectinophora gossypiella (Saunders) (Lepidoptera: Gelechiidae) by the chitin synthesis inhibitors novaluron and diofenolan. BioBulletin, 3: 8-21.
Direct Link - Busvin, J.R., 1962. A laboratory technique for measuring the susceptibility of house-flies and blowflies to insecticides. Lab. Pract., 11: 464-468.
Direct Link - Abbott, W.S., 1925. A method of computing the effectiveness of an insecticide. J. Econ. Entomol., 18: 265-267.
CrossRefDirect Link - Finney, D.J., 1971. Probit Analysis. 3rd Edn., Cambridge University Press, Cambridge, UK., ISBN-13: 9780521080415, Pages: 333.
Direct Link - Lotka, A.J., 1907. ART. XXII.--Studies on the mode of growth of material aggregates. Am. J. Sci., 24: 199-216.
CrossRefDirect Link - Jimenez-Peydro, R., C. Gimeno-Martos, J. Lopez-Ferrer, C. Serrano-Delgado and J. Moreno-Mari, 1995. Effects of the insect growth regulator cyromazine on the fecundity, fertility and offspring development of Mediterranean fruit fly, Ceratitis capitata Wied. (Dipt., Tephritidae). J. Applied Entomol., 119: 435-438.
CrossRefDirect Link - Tappozada, A., S. Abdallah and M.E. Eldefrawi, 1966. Chemosterilization of larvae and adults of the egyptian cotton leafworm, Prodenia litura, by Apholate, Metepa and Tepa. J. Econ. Entomol., 59: 1125-1128.
CrossRefDirect Link - Moroney, M.J., 1956. Facts from Figures. 3rd Edn., Pinguin Book Ltd., Harmondsworth, Middlesex, pp: 228.
Direct Link - Moreno, J., N. Hawlitzky and R. Jimenez, 1993. Effect of the juvenile hormone analogue fenoxycarb applied via the host on the parasitoid Phanerotoma (Phanerotoma) ocularis Kohl (Hym., Braconidae). J. Insect Physiol., 39: 183-186.
CrossRefDirect Link - Ghoneim, K.S., A.G. Al-Dali, R.F. Bakr, M.A. Tanani and A.S. Bream, 2007. Developmental and morphogenic responses of Schistocerca gregaria (Orthoptera: Acrididae) to the juvenoid pyriproxyfen (S-31183). Al-Azhar Bull. Sci., 18: 59-81.
CrossRefDirect Link - Gelbič I., J. Olejnı́ček and L. Grubhoffer, 2002. Effects of insect hormones on hemagglutination activity in two members of the Culex pipiens complex. Exp. Parasitol., 100: 75-79.
CrossRefDirect Link - Tripathi, P. and S.K. Tiwari, 2014. Potential of an insect growth regulator in the management of the rmoth Corcyra cephalonica stainton, 1866 (Lepidoptera: Pyralidae). Polish J. Entomol., 83: 79-97.
CrossRefDirect Link - Braga, I.A., C.B. Mello, A.A. Peixoto and D. Valle, 2005. Evaluation of methoprene effect on Aedes aegypti (Diptera: Culicidae) development in laboratory conditions. Mem. Inst. Oswaldo Cruz, 100: 435-440.
CrossRefDirect Link - Singh, Z., A. Singh, M. Kaur and T. Kaur, 2017. Assessment of barium carbonate toxicity on the developmental stages of Sarcophaga ruficornis (Diptera: Sarcophagidae). Int. J. Curr. Microbiol. Appl. Sci., 6: 485-494.
CrossRefDirect Link - Hamaidia, K., 2014. Laboratory evaluation of a biorational insecticide, kinoprene, against Culex pipiens larvae: effects on growth and development. Ann. Res. Rev. Biol., 4: 2263-2273.
CrossRefDirect Link - Linton, Y.M., A.J. Nisbet and A.J. Mordue, 1997. The effect of azadirachtin on the testes of the desert locust Schistocerca gregaria (Forskal). J. Insect Physiol., 43: 1077-1084.
CrossRef - Anger, K. and R.R. Dawirs, 1981. Influence of starvation on the larval development of Hyas araneus (Decapoda: Majidae). Helgol. Meeresuntersuchungen, 34: 287-311.
CrossRefDirect Link - Smagghe, G. and D. Degheele, 1994. The significance of pharmacokinetics and metabolism to the biological activity of RH-5992 (tebufenozide) in Spodoptera exempta, Spodoptera exigua and Leptinotarsa decemlineata. Pestcide Biochem. Physiol., 49: 224-234.
CrossRefDirect Link - Viñuela, E., F. Budia, J. Jacas, A. Adán, V. Marco and P.D. Estal, 1993. Differential larval age susceptibility of the medfly, Ceratitis capitata Wied. (Dipt., Tephritidae) to cyromazine. J. Appl. Entomol., 115: 355-362.
CrossRefDirect Link - Subrahmanyam, B., T. Muller and H. Rembold, 1989. Inhibition of turnover of neurosecretion by azadirachtin in Locusta migratoria. J. Insect Physiol., 35: 493-500.
CrossRefDirect Link - Seth, R.K., J.J. Kaur, D.K. Rao and S.E. Reynolds, 2004. Effects of larval exposure to sublethal concentrations of the ecdysteroid agonists RH-5849 and tebufenozide (RH-5992) on male reproductive physiology in Spodoptera litura. J. Insect Physiol., 50: 505-517.
CrossRefDirect Link - Carbone, M.A., K.W. Jordan, R.F. Lyman, S.T. Harbison and J. Leips et al., 2006. Phenotypic variation and natural selection at Catsup, a pleiotropic quantitative trait gene in Drosophila. Curr. Biol., 16: 912-919.
CrossRefDirect Link - Chamseddin, K.H., S.Q. Khan, M.L. Nguyen, M. Antosh and S.N.S. Morris et al., 2012. Takeout-dependent longevity is associated with altered juvenile hormone signaling. Mech. Ageing Dev., 133: 637-646.
CrossRefDirect Link - Yamamoto, R., H. Bai, A.G. Dolezal, G. Amdam and M. Tatar, 2013. Juvenile hormone regulation of Drosophila aging. BMC Biol., Vol. 11.
CrossRefDirect Link - Josephrajkumar, A., B. Subrahmanyam and S. Srinivasan, 1999. Plumbagin and azadirachtin deplete haemolymph ecdysteroid levels and alter the activity profiles of two lysosomal enzymes in the fat body of Helicoverpa armigera (Lepidoptera: Noctuidae). Eur. J. Entomol., 96: 347-353.
Direct Link - McNabb, S.L. and J.W. Truman, 2008. Light and peptidergic eclosion hormone neurons stimulate a rapid eclosion response that masks circadian emergence in Drosophila. J. Exp. Biol., 211: 2263-2274.
CrossRefDirect Link - Lucantoni, L., F. Giusti, M. Cristofaro, L. Pasqualini, F. Esposito, P. Lupetti and A. Habluetzel, 2006. Effects of a neem extract on blood feeding, oviposition and oocyte ultrastructure in Anopheles stephensi Liston (Diptera: Culicidae). Tissue Cell, 38: 361-371.
CrossRefDirect Link - Catchot, B., C.J. Anderson, J. Gore, R. Jackson, K. Rakshit, F. Musser and N. Krishnan, 2020. Novaluron prevents oogenesis and oviposition by inducing ultrastructural changes in ovarian tissue of young adult Lygus lineolaris. Lygus lineolaris Pest Manag. Sci., Vol. 76.
CrossRefDirect Link - Di Ilio, V., M. Cristofaro, D. Marchini, P. Nobe and R. Daliai, 1999. Effects of a neem compound on the fecundity and longevity of Ceratitis capitata (Diptera: Tephritidae). J. Econ. Entomol., 92: 76-82.
CrossRefDirect Link - Moreno, D.S., A.J. Martinez and M.S. Riviello, 1994. Cyromazine effects on the reproduction of Anastrephaludens (Diptera: Tephritidae) in the laboratory and in the field. J. Econ. Entomol., 87: 202-211.
CrossRefDirect Link - Zhou, F., G. Zhu, H. Zhao, Z. Wang and M. Xue et al., 2016. Sterilization effects of adult-targeted baits containing insect growth regulators on Delia antiqua. Sci. Rep., Vol. 6.
CrossRefDirect Link - Ouakid, M.L., Y. Adjami, W. Habbachi, R. Ghanem, H. Daas and A. Tahraoui, 2016. Insecticidal effect of halofenozide and methoxyphenoside in different stages of Lymantria dispar, an important fungal oak leafhopper. Turk. J. Agric. For., 17: 80-84.
Direct Link - Chapman, R.F., 1998. The Insects: Structure and Function. 1st Edn., Cambridge University Press, Cambridge.
Direct Link - Indrasith, L.S., T. Sasaki, T. Yaginuma and O. Yamashita, 1988. The occurrence of a premature form of egg-specific protein in vitellogenic follicles of Bombyx mori. J. Comp. Physiol. B, 158: 1-7.
CrossRefDirect Link - Davey, K.G. and D.R.B. Gordon, 1996. Fenoxycarb and thyroid hormones have JH‐like effects on the follicle cells of Locusta migratoria in vitro. Arch. Insect Biochem. Physiol., 32: 613-622.
CrossRefDirect Link