
Ronny Lesmana
Physiology Molecular Laboratory, Biological Activity Division, Central Laboratory, Universitas Padjadjaran, Bandung, Indonesia
LiveDNA: 62.22876
Susianti
Physiology Molecular Laboratory, Biological Activity Division, Central Laboratory, Universitas Padjadjaran, Bandung, Indonesia
Todia Pediatama
Physiology Molecular Laboratory, Biological Activity Division, Central Laboratory, Universitas Padjadjaran, Bandung, Indonesia
Nova Sylviana
Physiology Molecular Laboratory, Biological Activity Division, Central Laboratory, Universitas Padjadjaran, Bandung, Indonesia
Yuni Susanti Pratiwi
Physiology Molecular Laboratory, Biological Activity Division, Central Laboratory, Universitas Padjadjaran, Bandung, Indonesia
Hanna Goenawan
Physiology Molecular Laboratory, Biological Activity Division, Central Laboratory, Universitas Padjadjaran, Bandung, Indonesia
LiveDNA: 62.22347
Unang Supratman
Departement of Chemistry, Faculty of Mathematics and Natural Sciences, Universitas Padjadjaran, Bandung, Indonesia
Pakistan Journal of Biological Sciences
Year: 2021 | Volume: 24 | Issue: 1 | Page No.: 35-41







ABSTRACT
Background and Objective: The natural bioactive compounds of Curcuma longa, known as curcuminoids, has been shown to exerts anticancer effects to diverse cancer cell line in vitro, including breast cancer cell line. These curcuminoids consist of curcumin (Cur), demethoxycurcumin (DMC) and bisdemethoxycurcumin (BDMC). Furthermore, there has never been a study to compare the extent of antiproliferative and apoptotic modulation potential between Cur, DMC and BDMC in the breast cancer cell, until now. In the present study, we explore the efficacy among Cur, DMC and BDMC to alters MCF-7 cell viability, which might lead to apoptotic modulation. Materials and Methods: This kind of study was performed in vitro whereby the cells were maintained in an appropriate medium and the anticancer effect of curcuminoids (Cur, DMC and BDMC) was measured by using resazurin-based PrestoBlue cell viability assay. Later, MCF-7 breast cancer cells were cultured in 12 wells plate added with different concentrations of Cur, DMC and BDMC for western blotting analysis. Statistical analysis was performed with GraphPad 8, One-way ANOVA and Student’s t-test. Results: The result showed that Cur, DMC and BDMC inhibiting the proliferation of MCF-7 cells. In the concentration dose of 31.25 μg mL–1, the cell viability in cells treated with Cur is 27%, DMC is 31.5% and BDMC is 46%. The IC50 dose of Cur, DMC and BDMC were 25.63, 29.94 and 36.91 μg mL–1. Conclusion: Cur is more effective in inhibiting proliferation and apoptotic modulation in MCF-7 cells compare to DMC and BDMC. It represents the potential of Cur, DMC and BDMC as adjunctive therapy in treating breast cancer.
PDF Abstract XML References Citation
Copyright: © 2021. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Ronny Lesmana, Susianti, Todia Pediatama, Nova Sylviana, Yuni Susanti Pratiwi, Hanna Goenawan and Unang Supratman, 2021. Active Compounds from Curcuma longa and Comparison of their Effectively Induced Apoptosis in MCF-7 Cell. Pakistan Journal of Biological Sciences, 24: 35-41.
DOI: 10.3923/pjbs.2021.35.41
URL: https://scialert.net/abstract/?doi=pjbs.2021.35.41
DOI: 10.3923/pjbs.2021.35.41
URL: https://scialert.net/abstract/?doi=pjbs.2021.35.41
INTRODUCTION
Breast Cancer (BC) is the most frequently diagnosed cancer and the leading cause of cancer death in females worldwide. In 2018, approximately 2.1 million BC cases were newly diagnosed around the world. The incidence rate and mortality rate of BC are gradually increasing in low and middle-income countries (LMIC)1. For example, in Indonesia, the results of the 2018 Basic Health Research (Riskesdas) by the Ministry of Health of the Republic of Indonesia show that the incidence rate increased from 1.4 per 1000 in 2013 to 1.8 per 1,000 people with an average mortality rate of 17 per 100,000 people in 20182,3.
Surgical, chemotherapy, radiotherapy and endocrine therapy are the mainstay management of BC. However, the treatment of BC is still inadequate, especially in LMIC, where the mortality rate is higher compared to high-income countries4. Limited access to cancer therapy is the main reason for this disparity5. Another reason is that most of the new BC cases presentation in LMIC is already in the late-stage of the disease. The problem of late-stage presentation of BC is not only that it has a relatively low survival rate, but it also requires more toxic and more expensive treatment that will further burden the patient. New anticancer drugs such as immunotherapies are expensive and out-of-reach to most of LMIC5. Therefore, a novel, practical and affordable therapeutic approaches that have fewer treatment-related toxicity like herbal medicine are needed to improve clinical management of BC in LMIC.
Curcuma longa is basically a plant from the Zingiberaceae family which is endemic in Southeast and South Asia. Traditionally, majority of people use the powder of Curcuma longa, known as turmeric, as medicine for a range of diseases6. The natural bioactive compounds of Curcuma longa, known as curcuminoids, has been shown to exerts anticancer effects to diverse cancer cell line in vitro, including breast cancer cell line7,8. These curcuminoids consist of curcumin (Cur), demethoxycurcumin (DMC) and bisdemethoxycurcumin (BDMC) (Fig. 1). Previous studies showed that Cur exhibited anticancer properties in breast cancer cell through a plethora of signalling pathways9,10. Unlike Cur, there has not been an extensive mechanistic study of both DMC and BDMC to affect breast cancer cell11,12. Furthermore, there has never been a study to compare the extent of antiproliferative and apoptotic modulation potential between Cur, DMC and BDMC in the breast cancer cell, until now. In the present study, we explore the efficacy among Cur, DMC and BDMC to alters MCF-7 cell viability, which might lead to apoptotic modulation.
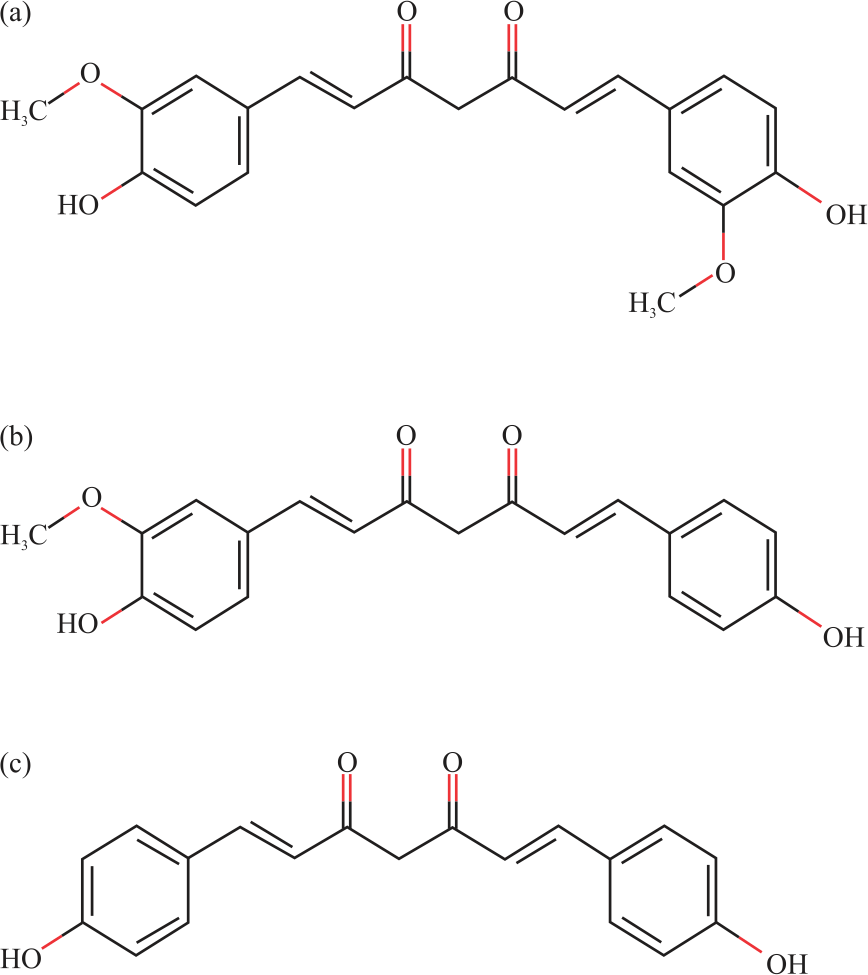 |
| Fig. 1: | The chemical structure of (a) Curcumin, (b) Demethoxycurcumin and (c) Bisdemethoxycurcumin |
MATERIALS AND METHODS
Study area: The study was carried out from April, 2019 to August, 2019 at the Central Laboratory of Universitas Padjadjaran, Jl. Raya Bandung Sumedang km 21, West Java, Indonesia, 45363.
Isolation of active compounds from Curcuma longa: The dried and powdered of Curcumin longa was extracted according to Sianturi et al.13 method with a modifications solvent using n-hexane, EtOAc, n-BuOH. The n-hexane dissolvable portion (26.3 g) was isolated by vacuum fluid chromatography, eluted with an angle of n-hexane-EtOAc (10:0-1:1), to give six subfractions (C01-C06). Subfraction C03 (250 mg) was chromatographed with CH2Cl2:CHCl3 (9.5:0.50), to give five subfractions (C03A-C03D). Subfraction C03C was isolated on preparative TLC, eluted with n-hexane:EtOAc (8.5:1.5), to give 1 (15.2 mg). Portion C and D were consolidated (1.80 g) and was chromatographed with a slope of n-hexane-EtOAc (10:1-1:10), to give seven subfractions (D01-D07). Subfraction D05 (340 mg) was chromatographed with an inclination of n-hexane-EtOAc (10:1-1:10) to bear the cost of four subfractions (D05A-D05D). Subfraction D05C was chromatographed with a slope of CHCl3-EtOAc (10:1-1:10) to give 2 (10.5 mg).
Cell culture: The breast cancer cell line, MCF-7 were purchased from the American Type Culture Collection (ATCC, MCF7-HTB22). The cells were maintained in DMEM supplemented with 10% FBS and 1% streptomycin-penicillin. Cells were incubated at 37oC, 5% CO2. The culture medium was changed every 48 h and the cells were subcultured when reached >80% confluency.
Cell viability: The cytotoxicity of curcuminoids was determined using resazurin-based PrestoBlue (Invitrogen, USA) cell viability assay. Cells were cultured in addition to 7.81, 15.62, 31.25, and 62.5 μg mL–1 of Cur, BDMC and DMC. Cells cultured without cisplatin served as the positive control and without any treatment act as a negative control. After 24 hrs, Prestoblue reagent was added directly and the cultured cells were incubated for 10 min in 37oC. Lastly, fluorescence was read by the scanner. The cell viability was then measured as a percentage of fluorescence in comparison to the negative control.
Western blotting: MCF-7 breast cancer cells were cultured in 12 wells plate added with different concentrations of Cur, BDMC and DMC. IC50 concentration and 1/10 of IC50 concentration dose were used. Cells added with cisplatin served as the positive control. Cells cultured without any treatment act as a negative control. Samples were resolved on 14% SDS-PAGE and then the protein was transferred to a nitrocellulose membrane. The primary antibodies of apoptotic marker LC3, p62, PARP, Caspase-3 and Caspase-9 were added for immunoblotting. β-actin protein expression served as the internal control for protein loading. Protein bands intensities were calculated using ImageJ software.
Statistical analysis: All statistical analysis was performed with GraphPad 8. One-way ANOVA was used to assess the statistical significance between groups. Student’s t-test was used to test the significance between two variables. A result of p<0.05 was considered a significant difference.
RESULTS
The methanolic remove from the dried stembark of Curcuma longa was focused and extricated progressively with n-hexane, ethyl acetic acid derivation and n-butanol. The n-hexane showed the nearness of triterpenoid mixes. By utilizing the triterpenoid test to control partitions, the n-hexane division was isolated by a blend of section chromatography on silica gel and preparative TLC on silica gel GF254 to bear the cost of two dammarane-type triterpenoids.
Curcumin is more effective than demethoxycurcumin and bisdemethoxycurcumin: The result from the cell viability assay showed that the proliferation of MCF-7 was inhibited by Cur, DMC and BDMC. All of the curcuminoids significantly reduced the cell viability in a dose-dependent response (Fig. 2). Then, we compared the efficacy of curcuminoid in comparison to the positive control. The IC50 dose of cisplatin is comparable with the curcuminoids dose of 31.25 μg mL–1. Cisplatin as positive control has 43% cell viability of the negative control.
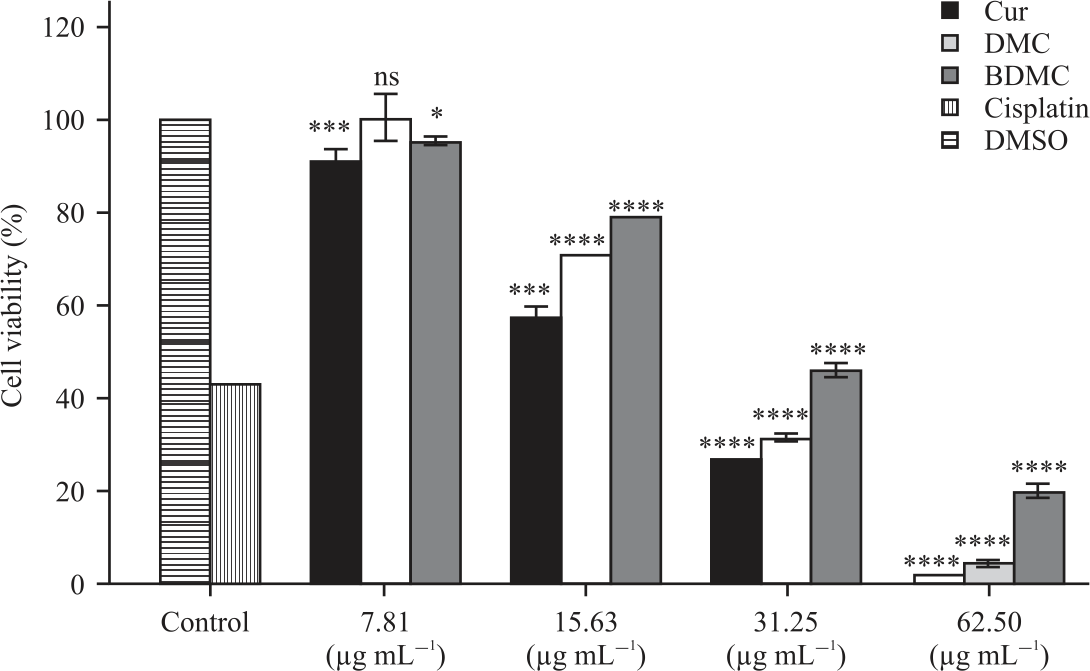 |
| Fig. 2: | The cell viability percentage of MCF-7 of curcumin, demethoxycurcumin and bisdemethoxycurcumin Negative control and the positive control in this study are DMSO 3% and cisplatin. Data was presented as average mean with standard deviation. p<0.05 was considered as significant, *p<0.05 and ****p<0.01 |
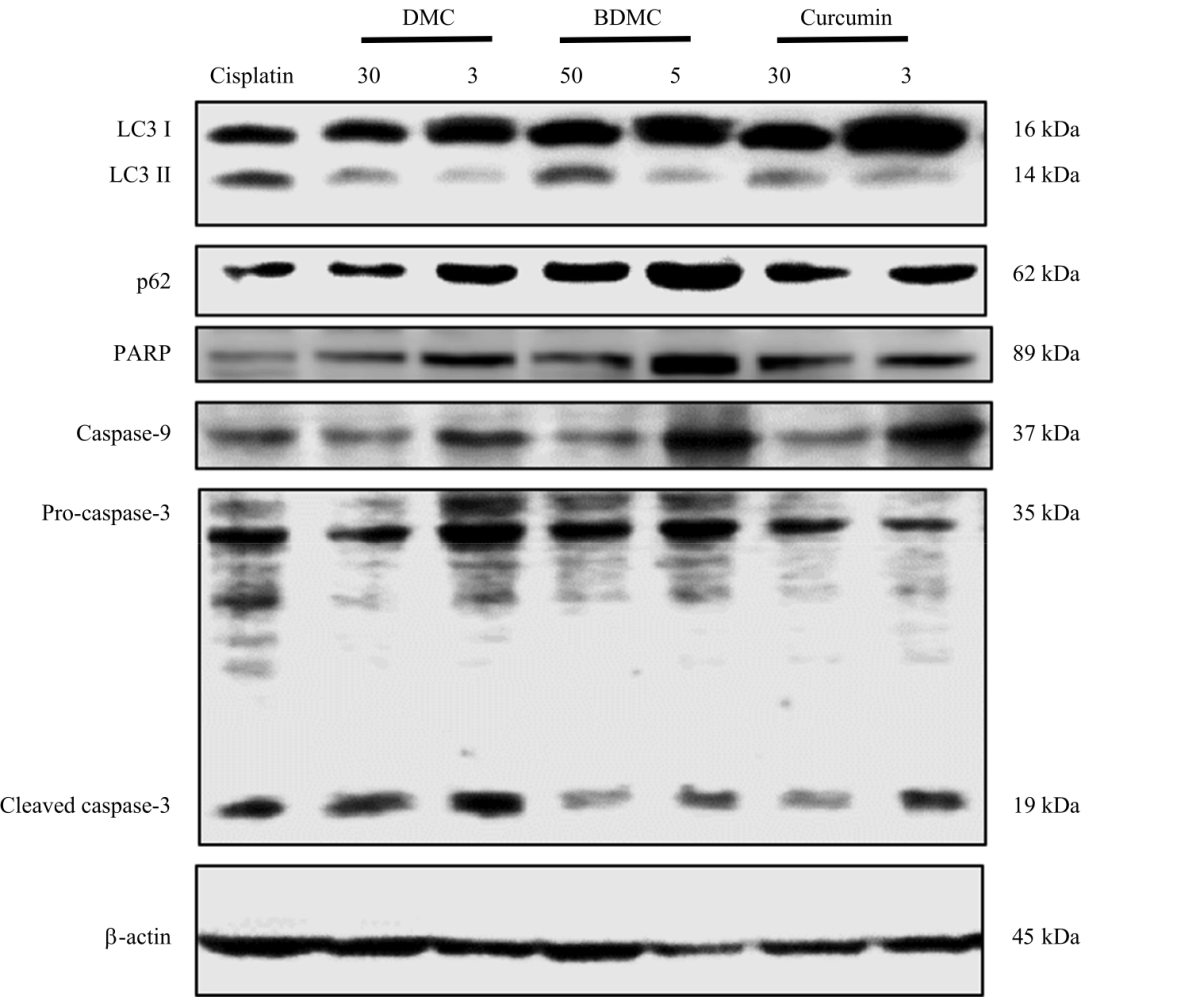 |
| Fig. 3: | Representative western blot membrane of autophagy markers (LC3II and p62), apoptosis signalling marker (PARP, Caspase 3 and Caspase 9) by curcumin, demethoxycurcumin and bisdemethoxycurcumin β-actin were used as housekeeping gene |
In the concentration dose of 31.25 μg mL–1, the cell viability in cells treated with Cur is 27%, DMC is 31.5% and BDMC is 46%. Cur and DMC in the concentration of 31.25 μg mL–1 were significantly more efficient to inhibit proliferation than cisplatin. There was no significant difference in cell viability between cells treated with BDMC in 31.25 μg mL–1 or cisplatin. We also calculated the IC50 dose of Cur, DMC and BDMC using this result. The IC50 dose of Cur, DMC and BDMC was 25.63, 29.94 and 36.91 μg mL–1. This shows that the degree of antiproliferative effect in the three curcuminoids are Cur>DMC>BDMC.
Comparison of autophagy marker LC3-I, LC3-II and p62 in curcumin, demethoxycurcumin and bisdemethoxycurcumin: To compare the effect of autophagy and apoptosis after treated with different curcuminoids, we observed the changes in protein expression of p62, LC3-I, LC3-II, PARP, Caspase-3 and Caspase-9 using western blot. The cells were treated with Cur, DMC and BDMC in high concentration (30 μg mL–1 for Cur, 30 μg mL–1 for DMC, 50 μg mL–1 for BDMC) and low concentration (3 μg mL–1 for Cur, 3 μg mL–1 for DMC, 5 μg mL–1 for BDMC) for 24 hrs (Fig. 3). The protein expression of p62 was downregulated in high dose DMC, BDMC and Cur. The lowest expression of p62 is in the high dose DMC treated cell compared to the other curcuminoids. Interestingly, low dose DMC, BDMC was shown to have a higher expression of p62 compared to the negative control.
Autophagy marker of LC3-II expression in high dose DMC, BDMC and Cur was decreased compared to the negative control. High dose DMC has the lowest expression of LC3-II compared to high dose BDMC and Cur. In low dose concentration, only Cur has decreased LC3-II protein expression. The ratio of LC3-II/LC3-I in all the samples is less than 1, with the highest ratio of all the samples is the negative control.
Apoptosis marker cleaved PARP is upregulated in Cur, DMC and BDMC treated cells compared to the negative controls. Compared to the other study group, low dose DMC and BDMC has the highest expression of cleaved PARP.
The expression of apoptotic marker Cas 9 and Cas 3 was also measured to investigate whether the mechanism of apoptosis is mediated through the caspase pathway. In this study, the protein expression of cleaved Cas 9 is much lower than negative and positive control in high dose DMC, BDMC and Cur. Protein expression of cleaved Cas 3 is also downregulated in a high dose of DMC and BDMC, but not Cur. Cur has a similar Cas 3 expression with the Cisplatin treated cells.
DISCUSSION
The active compounds in which have been isolated from Curcuma longa such as Curcumin, Demethoxycurcumin and Bisdemethoxycurcumin by using some chemicals, were found to be bioactive. Cur, DMC and BDMC have been shown in previous studies to inhibit the proliferation of MCF-712,14,15. The result from our study confirmed that the curcuminoids have a potent antiproliferative effect on MCF-7 and it affected the cell viability of MCF-7 in a dose-dependent manner. It was compared to the inhibitory effect of the three curcuminoids and discovered that Cur is more effective than both DMC and BDMC in inhibiting the proliferation of MCF-7. One study compared the effect of the three curcuminoids to MCF-7 and contrary to our result, DMC is more potent to inhibit the proliferation of MCF-7 than Cur and BDMC. It is believed that the phenolic hydroxyl groups, phenolic methoxyl group and the diketone moiety in the chemical structure of DMC are essential to the effectivity of antiproliferative activity compared to Cur and BDMC16. The discrepancy of the result of antiproliferative comparison between Cur, DMC and BDMC with the previous study could be attributed to the low number of replicates performed in the study, a duplicate for each group17.
Then, it was evaluated the autophagic activity in MCF-7 induced with curcuminoids by measuring the p62 and LC3 protein expression after being treated with curcuminoids. The amount of LC3-II protein that is converted from LC3-I is correlated with the number of formed autophagosomes and thus correlated with autophagic activity. High autophagic activity will result in low expression of p62 as the protein is degraded through autophagy18,19. In this study, p62 protein expression is found to be increased in MCF-7 treated with low dose DMC and BDMC. Accumulation of p62 protein could suggest that there is an inhibition of autophagy. High dose DMC, BDMC, Cur and low dose Cur have a downregulated p62 protein expression as expected. Surprisingly, the LC3-II protein expression is low in cells treated with a high dose of DMC, BDMC and Cur. In low dose DMC and BDMC treated cells, the LC3-II protein expression is comparable to the negative controls. However, in our results, the protein expression of LC3-I that is converted to LC3-II is low in the group with low p62 expression.
In contrast to our result, the previous study that investigated the synergistic activity of curcumin and berberine on MCF-7 cells has shown that even Cur alone can induce an increase in LC3-II and decrease in p62 protein expression20. Another study that investigated the combined effect of DMC and photodynamic therapy on MDA-MB-231 breast cancer cell line also shown that even DMC treated cells alone have decreased p62 and increase in LC3-II11. A previous study investigated the autophagy effect of curcuminoids in oral cancer cells, it was shown that BDMC, along with Cur and DMC promotes autophagy as the LC3-II expression is increased and p62 is decreased21. It seems possible that the low LC3-II protein expression in our results is because of the cells are being cultured in a more extended period with a higher concentration of curcuminoids22. Even though LC3-II regarded as the marker of autophagy, LC3-II can also be degraded itself through the activity of autophagy23. Thus, the LC3-II may be degraded in high dose curcuminoids because of the autophagic activity is more intense in this group.
Our result in the expression of caspase-3 and caspase-9 suggests that the mechanism of DMC, BDMC and Cur to induce apoptosis in MCF-7 cell is different from each other. The decreased expression of both cleaved Cas-9 and Cas-3 expression in high dose DMC and BDMC suggested that there is an inhibition in the caspase-dependent pathway apoptosis mechanism and the apoptosis might be mediated through the caspase-independent pathway (extrinsic pathway). On the other hand, Cur induces apoptosis through the activation of caspase-3. DMC has been shown to activates caspase-3 and caspase-9 to induce caspase-dependent apoptosis cascade in cutaneous squamous cell carcinoma, A431 and HaCaT cell line24. Cur has been shown to induces apoptosis through a caspase-9 dependent mechanism in MDA-MB-231 breast cancer cell line25. Other studies also showed that BDMC activates caspase-3 and caspase-9 in MCF-7 and NCI H460 lung cancer cell line15,26.
There is also a possibility that the incubation period of 24 hours treated with a high dose of curcuminoids this study is too long, or it can also be that the dose is too high. This can be seen by the high protein expression of apoptosis and autophagy marker in negative control and the low dose of curcuminoids treated cell. Most of the previous studies treated the cells in 15-50 μM of Cur, DMC, or BDMC for 24 hours.
Based on the findings, while preliminary, it could be hypothesized that the mechanism of Cur, DMC and BDMC to induce apoptosis might be through various pathways, the intrinsic and extrinsic pathway and might have a different order of anticancer effectivity. Nevertheless, this study did not include the marker for the extrinsic pathway or other pathways such as necroptosis. Even though the results undoubtedly be scrutinized, the conclusion that Cur, DMC and BDMC have an antiproliferative effect on MCF-7 cells is solid. It represents the potential of Cur, DMC and BDMC as adjunctive therapy in treating breast cancer. As apparent Cur expresses various compounds such as anti-inflammatory, anti-antigenic, anti-metastatic, anti-oxidant activities in animal experiments, so it could be a highly potential molecule in preventing the occurrence of cancer in humans. Basically, Cur produces antiproliferative activities in various kinds of cell lines in vitro and then utilize it through apoptosis modulation in breast cancer cells27. Previous research showed that curcumin fundamentally repressed the growth of breast cancer cell MCF-7 by initiated apoptosis in a dose- and time-dependent manner, accompanied by a decrement in cell viability of MCF- 7. Curcumin is commonly cytotoxic on a few colon cancer cell lines. It hindered the passage to cell cycle from G2 to M by restraining articulation of cdc2/cyclin B. The proapoptotic individuals from the Bcl-2 family, for example, Bax, were actuated and antiapoptotic qualities, for instance, Bcl-XL were repressed by curcumin27.
CONCLUSION
In this study, we confirmed that Cur is the more effective inhibiting proliferation and apoptotic modulation in MCF-7 cells compare to DMC and BDMC. It represents the potential of Cur, DMC and BDMC as adjunctive therapy in treating breast cancer.
SIGNIFICANCE STATEMENT
This study discovered the Cur, DMC and BDMC in Curcuma longa that can be beneficial for the development of adjunctive therapy in treating breast cancer. This study will help the researchers to uncover the critical areas of Curcuma longa compound bioactivity effectiveness that many researchers were not able to explore. Thus a new theory on the natural bioactive compound may be arrived at.
ACKNOWLEDGMENT
This work is funded by PDUPT No: 3854/Unc.6/2019 from Ministry Research, Technology and Higher Education to RL (2019).
REFERENCES
- Bray, F., J. Ferlay, I. Soerjomataram, R.L. Siegel, L.A. Torre and A. Jemal, 2018. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA: Cancer J. Clin., 68: 394-424.
CrossRefDirect Link - Diaz, R.A.N., N.N.T. Swandewi and K.D.P. Novianti, 2019. Malignancy determination breast cancer based on mammogram image with k-nearest neighbor. 2019 1st International Conference on Cybernetics and Intelligent System (ICORIS), 22-23 Aug, 2019, IEEE, pp: 233-237.
CrossRefDirect Link - Umami, A., S. Sudalhar, W. Tegar, P. Yudha, F. Istiqlal and F. Andan, 2020. Knowledge, barriers and motivation related to breast and cervical cancer screening among women in bojonegoro, East Java: A qualitative study. J. Health Promotion Behav., Vol. 5.
CrossRefDirect Link - Martei, Y.M., L.E. Pace, J.E. Brock and L.N. Shulman, 2018. Breast cancer in low- and middle-income countries. Clin. Lab. Med., 38: 161-173.
CrossRefDirect Link - Ruff, P., S. Al-Sukhun, C. Blanchard and L.N. Shulman, 2016. Access to cancer therapeutics in low- and middle-income countries. Am. Soc. Clin. Oncol. Educ. Book, 36: 58-65.
CrossRefDirect Link - Kocaadam, B. and N. Sanlier, 2017. Curcumin, an active component of turmeric (Curcuma longa) and its effects on health. Crit. Rev. Food Sci. Nutr., 57: 2889-2895.
CrossRefDirect Link - Hatamipour, M., M. Ramezani, S.A.S. Tabassi, T.P. Johnston, M. Ramezani and A. Sahebkar, 2018. Demethoxycurcumin: A naturally occurring curcumin analogue with antitumor properties. J. Cell Physiol., 233: 9247-9260.
CrossRefDirect Link - Ramezani, M., M. Hatamipour and A. Sahebkar, 2017. Promising anti-tumor properties of bisdemethoxycurcumin: A naturally occurring curcumin analogue. J. Cell Physiol., 233: 880-887.
CrossRefDirect Link - Choudhuri, T., S. Pal, M.L. Aggarwal, T. Das and G. Sa, 2002. Curcumin induces apoptosis in human breast cancer cells through p53-dependent Bax induction. FEBS Lett., 512: 334-340.
CrossRefPubMedDirect Link - Masuelli, L., M. Benvenuto, M. Fantini, L. Marzocchella and P. Sacchetti et al., 2013. Curcumin induces apoptosis in breast cancer cell lines and delays the growth of mammary tumors in neu transgenic mice. J. Biol. Regulators Homeostatic Agents, 27: 105-119.
CrossRefDirect Link - Lin, H.Y., J.N. Lin, J.W. Ma, N.S. Yang, C.T. Ho, S.C. Kuo and T.D. Way, 2015. Demethoxycurcumin induces autophagic and apoptotic responses on breast cancer cells in photodynamic therapy. J. Funct. Foods, 12: 439-449.
CrossRefDirect Link - Li, Y.B., J.L. Gao, Z.F. Zhong, P.M. Hoi, S.M.Y. Lee and Y.T. Wang, 2013. Bisdemethoxycurcumin suppresses MCF-7 cells proliferation by inducing ROS accumulation and modulating senescence-related pathways. Pharmacol. Rep., 65: 700-709.
CrossRefDirect Link - Sianturi, J., K. Farabi, T. Mayanti, D. Harneti and D. Darwati et al., 2016. Antioxidant constituents from the bark of Aglaia eximia (Meliaceae). Makara J. Sci., Vol. 20.
CrossRefDirect Link - Zhou, Q.M., X.F. Wang, X.J. Liu, H. Zhang, Y.Y. Lu and S.B. Su, 2011. Curcumin enhanced antiproliferative effect of mitomycin C in human breast cancer MCF-7 cells in vitro and in vivo. Acta Pharmacol. Sin., 32: 1402-1410.
CrossRefDirect Link - Mohankumar, K., S. Pajaniradje, S. Sridharan, V.K. Singh and L. Ronsard et al., 2014. Mechanism of apoptotic induction in human breast cancer cell, MCF-7, by an analog of curcumin in comparison with curcumin – An in vitro and in silico approach. Chemico-Biol. Interact., 210: 51-63.
CrossRefDirect Link - Sianipar, E.A., M. Louisa and S.I. Wanandi, 2018. Kurkumin meningkatkan sensitivitas sel kanker payudara terhadap tamoksifen melalui penghambatan ekspresi p-glikoprotein dan breast cancer resistance protein. Galenika J. Pharm., 4: 1-11.
CrossRefDirect Link - Klionsky, D.J., F.C. Abdalla, H. Abeliovich, R.T. Abraham and A. Acevedo-Arozena et al., 2012. Guidelines for the use and interpretation of assays for monitoring autophagy. Autophagy, 8: 445-544.
CrossRefDirect Link - Bjørkøy, G., T. Lamark, S. Pankiv, A. Øvervatn, A. Brech and T. Johansen, 2009. Chapter 12 monitoring autophagic degradation of p62/SQSTM1. Methods Enzymol. Autophagy Mamm. Sys., Part B 452: 181-197.
CrossRefDirect Link - Wang, K., C. Zhang, J. Bao, X. Jia and Y. Liang et al., 2016. Synergistic chemopreventive effects of curcumin and berberine on human breast cancer cells through induction of apoptosis and autophagic cell death. Sci. Rep., Vol. 6.
CrossRefDirect Link - Hsiao, Y.T., C.L. Kuo, F.S. Chueh, K.C. Liu, D.T. Bau and J.G. Chung, 2018. Curcuminoids induce reactive oxygen species and autophagy to enhance apoptosis in human oral cancer cells. Am. J. Chin. Med., 46: 1145-1168.
CrossRefDirect Link - Mizushima, N. and T. Yoshimori, 2007. How to interpret LC3 immunoblotting. Autophagy, 3: 542-545.
CrossRefDirect Link - Yoshii, S.R. and N. Mizushima, 2017. Monitoring and measuring autophagy. Int. J. Mol. Sci., 18: 1-13.
CrossRefDirect Link - Xin, Y., Q. Huang, P. Zhang, W.W. Guo, L.Z. Zhang and G. Jiang, 2017. Demethoxycurcumin in combination with ultraviolet radiation B induces apoptosis through the mitochondrial pathway and caspase activation in A431 and HaCaT cells. Tumour Biol.
CrossRefDirect Link - Gogada, R., M. Amadori, H. Zhang, A. Jones and A. Verone et al., 2011. Curcumin induces Apaf-1-dependent, p21-mediated caspase activation and apoptosis. Cell Cycle, 10: 4128-4137.
CrossRefDirect Link - Yang, S.T., A.C. Huang, N.Y. Tang, H.C. Liu and C.L. Liao et al., 2015. Bisdemethoxycurcumin-induced S phase arrest through the inhibition of cyclin A and E and induction of apoptosis via endoplasmic reticulum stress and mitochondria-dependent pathways in human lung cancer NCI H460 cells. Environ. Toxicol., 31: 1899-1908.
CrossRefDirect Link - Koohpar, Z.K., M. Entezari, A. Movafagh and M. Hashemi, 2015. Anticancer activity of curcumin on human breast adenocarcinoma: Role of Mcl-1 gene. Iran J. Cancer Preven., Vol. 8.
CrossRefDirect Link