
Rollando Rollando
Department of Chemistry, Faculty of Mathematics and Natural Sciences, Brawijaya University, Malang 65145, Indonesia
LiveDNA: 62.33120
Warsito Warsito
Department of Chemistry, Faculty of Mathematics and Natural Sciences, Brawijaya University, Malang 65145, Indonesia
Masruri Masruri
Department of Chemistry, Faculty of Mathematics and Natural Sciences, Brawijaya University, Malang 65145, Indonesia
Widodo Widodo
Department of Biology, Faculty of Mathematics and Natural Sciences, Brawijaya University, Malang 65145, Indonesia
Pakistan Journal of Biological Sciences
Year: 2021 | Volume: 24 | Issue: 2 | Page No.: 172-181







ABSTRACT
Background and Objective: Pterygota alata (Roxb.) R.Br. bark extract has been studied to have cytotoxic activity on 4T1 cells. This study was conducted to determine the cytotoxic activity of several fractions of Pterygota alata (Roxb.) R.Br. bark against 4T1 breast cancer cells and to investigate the most active fractions on Bcl-2 and Bax expressions. Materials and Methods: The bark of Pterygota alata (Roxb.) R.Br. was extracted using 80% methanol and was fractionated into fractions of n-hexane, chloroform, ethyl acetate, n-butanol and insoluble n-butanol with liquid-liquid partition. Cytotoxic tests were performed using the MTT method and expressions of Bax and Bcl-2 on 4T1 breast cancer cells were detected with immunocytochemical staining. Identification of compounds in the most active fraction using GC-MS. Results: The results showed that the most active fraction was the insoluble fraction of n-butanol (IFB) with an IC50 of 15.14 μg mL–1. IFB also decreases the expression of Bcl-2 and increases the expression of Bax. Conclusion: It can be concluded that Pterygota alata (Roxb.) R.Br. bark has the potential to be developed for medical use, especially for breast cancer therapy.
PDF Abstract XML References Citation
Copyright: © 2021. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Rollando Rollando, Warsito Warsito, Masruri Masruri and Widodo Widodo, 2021. Pterygota alata (Roxb.) R.Br. Bark Fraction Induced Intrinsic Apoptotic Pathway in 4T1 Cells by Decreasing Bcl-2 and Inducing Bax Expression. Pakistan Journal of Biological Sciences, 24: 172-181.
DOI: 10.3923/pjbs.2021.172.181
URL: https://scialert.net/abstract/?doi=pjbs.2021.172.181
DOI: 10.3923/pjbs.2021.172.181
URL: https://scialert.net/abstract/?doi=pjbs.2021.172.181
INTRODUCTION
Breast cancer has a high impact on increasing mortality in women. The main reason is due to the lack of treatment options that are safer, more effective and do not have high side effects1. The prevalence of cancer in Indonesia has increased significantly from 1.4 per 1000 in 2013 to 1.79 per 1000 populations in 2018. The highest incidence rate for women is breast cancer, which is 42.1 per 100.000 population. The mortality rate of 17 per 100.000 population, followed by cervical cancer was 23.4 per 100.000 population with an average death rate of 13.9 per 100.000 population2. Globocan data states that in 2018 there were 18.1 million new cases with a death rate of 9.6 million deaths, where 1 in 6 women in the world experience cancer3. Genetic mutations are the leading cause of breast cancer (85%), specifically in the BRCA1 and BRCA2 genes4. Much evidence from research states that natural products used in traditional medicine systems have great potential to treat breast cancer5, this is due to the extensive chemical content and bioactive molecules that are curative compared to destructive chemotherapy drugs6. This causes many scientists to be interested in researching and discovering new compounds from natural products.
Pterygota alata is a plant that has a cytotoxic effect. Rollando et al.7 proved that the methanol fraction of P. alata-bark had a cytotoxic effect on T47D and MCF-7 cells. Besides, El-Sherei et al.8 reported that aqueous methanol extract of P. alata bark has an anti-hyperglycemic effect and anti-oxidative stress. Lin et al.9 reported that the leaves and bark of P. alata have 1-O-cinnamoyl 6-O-p-coumaroyl glucoside compounds. De Souza et al.10 reported 1-O cinnamoyl 6-O-p-coumaroyl glucoside capable of inhibiting the activation of nuclear factor (NF) κB, which has a crucial role in inflammation, oxidative processes and anti-apoptosis.
Apoptosis is a mechanism for programmed cell death to maintain the number of cells in the body or prevent the growth of cells that are not needed11. Apoptosis occurs via intrinsic and extrinsic pathways12. Intrinsic pathways are induced in response to DNA damage, glucocorticoids, ceramides and decreased growth factors that cause changes in the mitochondrial membrane13. In contrast, the extrinsic pathway is induced by ligand (TNF, FasL) with transmembrane death receptors, namely Fas and Tumor Necrosis Factor Receptor (TNFR-1)14. Changes in the potential of the mitochondrial membrane depend on the ratio between the antiapoptotic protein (Bcl-2) and proapoptotic protein (Bax)15. 4T1 cell lines are reported to have overexpression of Bax- and Bcl-216. Based on the results of this study, the study was conducted to determine the cytotoxic activity and expression of Bax and Bcl-2 from 4T1 cells treated with P. alata bark fraction.
MATERIALS AND METHODS
Study area: The study was carried out at Microbiology and Phytochemistry Lab, Department of Pharmacy, Ma Chung University from April-July, 2020.
Chemicals: Methanol, n-hexane, chloroform, ethyl acetate, n-butanol was purchased from Merck, Germany. 4T1 cells (American Type Culture Collection, Manassas, VA), were cultured in RPMI-1640 medium containing 10% (v/v) Fetal Bovine Serum (FBS) and 1% penicillin-streptomycin (Life Technologies, USA) at 37°C in 5% CO2 humidified incubator. Vero cells were courtesy from Parasitology Laboratory, Medical Faculty, Gadjah Mada University cultured in Dulbecco’s Modified Eagle Media (DMEM). Doxorubicin (DOX) and 3-(4,5-dimethylthiazol-zyl)-2,5-diphenyl tetrazolium bromide (MTT) were obtained from Sigma, USA. The 4T1 cells (ATCC® CRL-2539™) were incubated with mouse monoclonal anti-Bcl-2 (Dako, Carpinteria, USA) or rabbit polyclonal anti-Bax (Dako) antibody, biotinylated universal secondary antibody (Biocare, USA) and streptavidin HRP label (Biocare). Finally, the cells were added with the DAB substrate (Dako).
Plant collections and extractions: Pterygota alata collected from a specific area of Kupang City, East Nusa Tenggara, Indonesia. Plant identification using the library Flora of Java and identified by Taxonomist, namely Dr. Budi Sumarta. The voucher specimens were deposited in the Laboratory of Pharmacognosy, Ma Chung University Malang, Indonesia with specimen numbers as followed: Pterygota alata (Roxb.) R.Br (MCH/002/X). Extraction using the maceration method, 3.0 kg of dry bark soaked using 80% methanol for 24 hrs. The supernatant and pulp are separated, the supernatant is evaporated until a thick extract is obtained.
Liquid-liquid partitions: About 60.0 g of the extract is placed into the separating funnel and 600 mL of 80% methanol is added. Total 600 mL of n-hexane was added into the solution and vigorously shaken until dual-phase being formed. The hexane phase was collected and then the 80% methanol phase was added and was re-shaken with the chloroform in the same volume ratio until a separate phase was formed. The obtained n-hexane and chloroform phases were then evaporated under reduced pressure to collect n-hexane and chloroform fractions. The 80% methanol phase was added and re-shaken with an equal volume of ethyl acetate until two separate phases were formed. The ethyl acetate phase and 80% methanol phase were dried until ethyl acetate and 80% methanol fractions were obtained. The 80% dry methanol partition was added with n-butanol and stirred for 2 hrs to form an n-butanol partition and an insoluble n-butanol partition. The two partitions were evaporated to obtain n-butanol and insoluble n-butanol fractions. All fractions were used for in vitro cytotoxic tests.
In vitro cytotoxicity assay: Determine the cytotoxicity of on 4T1 and Vero cells; the MTT assay was employed. 4T1 is a triple-negative breast cancer cell line from Mus musculus17 which contains PEX9 and Vero cell is a non-tumorigenic cell from the kidney tissue of African green monkey18. The assay detects the reduction of MTT and reflects the normal functioning of mitochondria and hence cytotoxicity. Fraction solutions were prepared in the following concentrations: 10, 20, 40, 80, 160, 320, 640 and 1280 μg mL–1. Cisplatin was added to the cultures at the following concentrations: 0, 2,5, 5, 10, 20 and 40 μg in a final volume of 100 μL as a positive control using DMSO 0.1%. Cells (1×104/well) were seeded in 96-well at-bottomed plates and incubated with each extract at various concentrations for 24 hrs. Total 30 μL of MTT solution (5 mg mL–1 in PBS) was added to each well and the plate was incubated at 37°C for another 4 hrs. Then, the medium was discarded and 150 μL of DMSO was added to dissolve the formazan crystals. The test was carried out for 5 replications. The absorbance of each sample was read at 595 nm using a microplate reader. Results were expressed as a percentage of cell viability to untreated control cells (as 100%). Microsoft excel prepared the data calculation and the drug-dose dependent curve.
Immunocytochemistry: T41 cells were planted at a density of 5×104 cells/well. Upon reaching 90% confluent, the cells were treated with ¼IC50, ½IC50 and IC50 of the IFB for 24 hrs. After that, the cells were washed and fixed with methanol, then treated with 3.7% H2O2 in methanol for 10 min. Then, the cells were washed and treated with blocking serum for 15 min. The cells were added with a monoclonal anti-Bcl-2 mouse or rabbit polyclonal anti-Bax antibody for 60 min, a biotinylated universal secondary antibody for 20 min and streptavidin HRP label for 10 min. At the end of the process, the cells were added with DAB substrate (Dako) to visualize the antigen-antibody bound and counterstained with hematoxylin. The control group is treated with 1% ethanol as vehicle control. Expressions of Bcl-2 and Bax were observed as the brown color at the cytoplasm and membranes of 4T1 cells under the microscope. The test was carried out five times of repetition. The expressions were then semi quantified using ImageJ version 1.51w software.
GC-MS analysis metabolomic-based assay: The structural identification of the most active fraction was carried on GC-MS. For this 1 mg of the fraction was dissolved in 1 mL of dichloromethane. Rtx®-50 column (30×250×0.25 μm) was used as the stationary phase with helium as a carrier gas at a flow rate of 1 mL min–1 and a pressure of 8.80 psi. The GC oven temperature was kept at 70 and 300°C for initial and final temperatures, respectively. The temperature rate increase was set at 10°C min–1 for 15 min. Splitless injections were implemented with the liquid injection method in this experiment. A library search for the structural identification was carried out using Wiley 7 Nist05 library.
Data analysis: Statistical analysis was performed using a one-way analysis of variance (ANOVA method) followed by post hoc Tukey’s multiple comparisons test in SPSS 20 software. Differences were considered significant at the p<0.05 level.
RESULTS
In vitro cytotoxicity assay: The purpose of this test is to determine the cytotoxic effect of each fraction resulting from the separation. Besides, this test is used to evaluate the selectivity index of the fraction resulting from separation against normal cells. The in vitro cytotoxic test was carried out using the MTT method. The principle of the MTT method is the reduction of the yellow salt of tetrazolium MTT (3-(4.5-dimethylthiazol-2-il)-2,5-diphenyltetrazolium bromide) by the reductase system. First, a test of each fraction resulted from the separation of 4T1 and Vero cells was conducted. We used 4T1 cells as a model of highly metastatic triple-negative breast cancer and Vero as a model for normal cells. Fractions treatment showed a dose-dependent phenomenon in inducing 4T1 cell death. The percentage of viable 4T1 cells was decreased along with the increase of fractions concentration (Fig. 1). The cytotoxic test results showed that IFB was the most active fraction in inhibiting 4T1 cells with an IC50 (Table 1). IFB had no significant difference with cisplatin as a positive control (p>0.05). Another fraction showed weak activity against 4T1 cells with a value range of 88.24-748.24 μg mL–1.
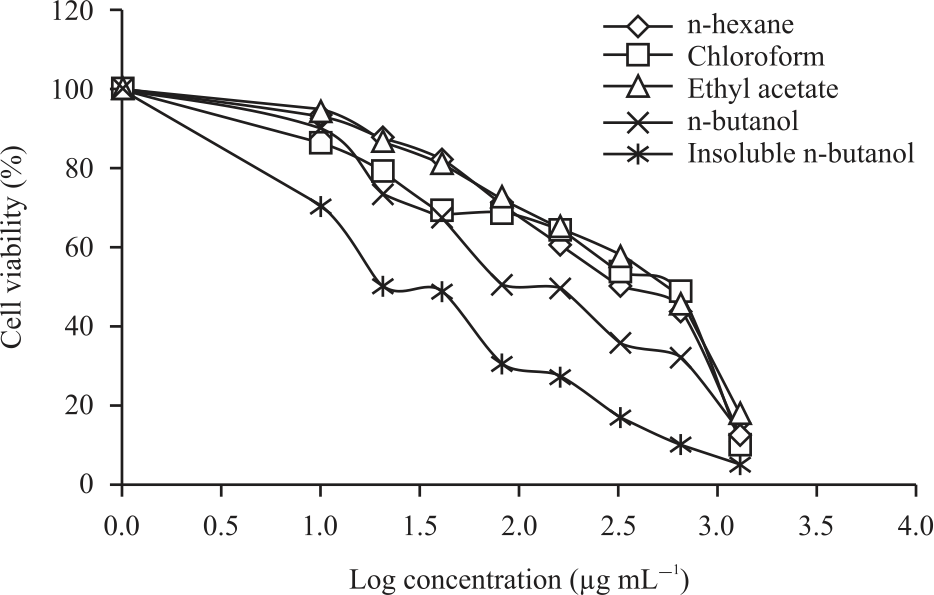 |
| Fig. 1: | Drug dose-dependent curves of five fractions against 4T1 cells |
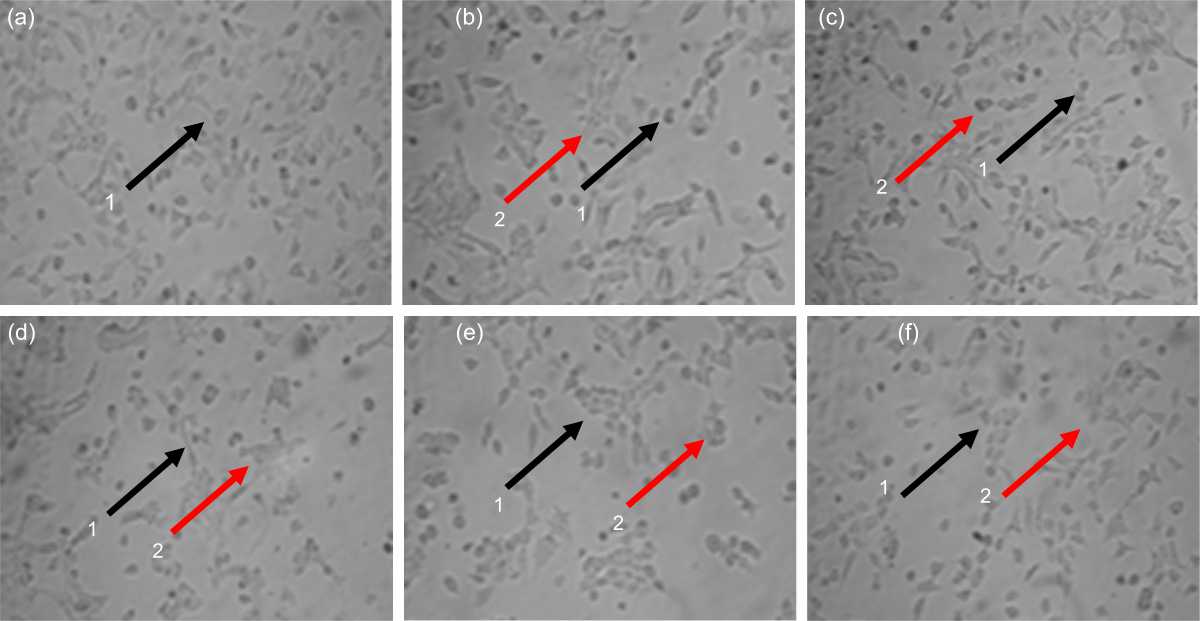 |
| Fig. 2(a-f): | 4T1 cell morphologies after and before treatment with IC50 concentration on each fraction for 24 hrs as followed, (a) Untreated cell, (b) n-Hexane, (c) Chloroform, (d) n-Butanol, (e) IFB and (f) Cisplatin 1: Viable cell, 2: Dead cell. Bar: 100 μm |
The results of the cytotoxic test fraction of another test against Cisplatin as positive control showed a significant difference (p<0.05). The potential of a compound is classified as a strong cytotoxic agent if the IC50 value <20 μg mL–1, moderate if the IC50 value <50 μg mL–1 and weak if the IC50 value >50 μg mL–1.
The selectivity index of chemo preventive agents is a test parameter to determine that only cells identified as cancer cells are inhibited growth. The higher the selectivity index value means that the test compound can inhibit cancer cells and has a low cytotoxic effect on normal cells. A small selectivity index indicates that the compound is less toxic to normal cells rather than to cancer cells. Compounds with high SI values offer the potential for safer and more effective therapy in cancer therapy. The test results showed that IFB was selective against 4T1 cells with SI values>3, while the other fractions were less selective against 4T1 cells with SI<3.
Morphology fraction-treated 4T1 cells: Morphological changes in 4T1 cells after administration of each fraction and positive control with a concentration of IC50 for 24 hrs are shown in Fig. 2. Living cells appeared as leaf-shape and had black spots, while dead cells as round and white. Cells show morphological changes that are possible because the cytoskeleton is cut and proteins that play a role in cell adhesions do not undergo polymerization so that the cell bonds are released and the lipid membrane will become round (Fig. 2a-f).
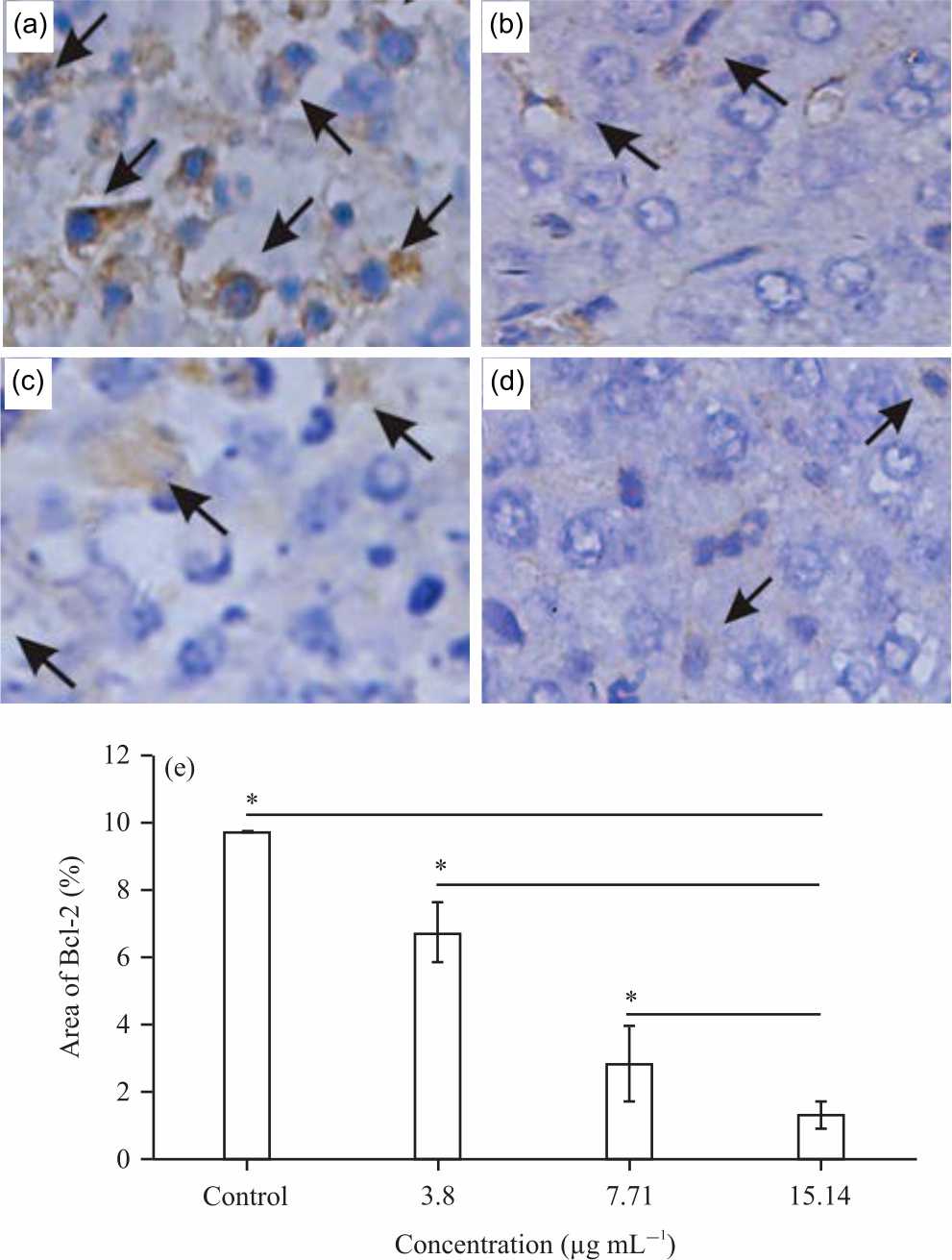 |
| Fig. 3(a-e): | Bcl-2 expression of IFB on 4T1 cells. 4T1 cells were treated with (a) Control, (b) 3.8, (c) 7.71, (d) 15.14 μg mL–1 IFB and (e) Graphical representation of Bcl-2 expression Cells were processed with immunohistochemistry to detect Bcl-2 as described in Methods. Semiquantified Bcl-2 expression with ImageJ. Bar: 150 μm. Values area means±SD, n = 5 Replicates, *p-value<0.05 then indicate statistically significant differences in comparison to control |
| Table 1: Results of in vitro cytotoxicity assay of five fractions and cisplatin against 4T1 and Vero cell growth | |||
| Samples | 4T1 | Vero | index |
| n-hexane | 748.24±23.41* | 838.13±24.70* | 1.12 |
| Chloroform | 77.98±12.38* | 223.73±18.13* | 2.86 |
| Ethyl acetate | 89.24±16.74* | 257.88±33.14* | 2.88 |
| n-butanol | 88.24±18.83* | 256.18±18.13* | 2.90 |
| Insoluble n-butanol | 15.14±2.81 | 194.41±19.18* | 12.84 |
| Cisplatin | 6.62±0.13 | 87.82±1.33 | 13.27 |
| Values area means±SD, n = 5 replicates, *p-value<0.05 then indicate statistically significant differences in comparison to cisplatin | |||
Bcl-2 expressions of IFB-treated 4T1 cells: The expression of Bcl-2 can be seen in Fig. 3. Expressions of Bcl-2 were indicated by the presence of Bcl-2 monoclonal antibodies bonds, detected in brown color in the cytoplasm and membrane of 4T1 cells (Fig. 3a-d).
 |
| Fig. 4(a-e): | Bax expression of IFB on 4T1 cells 4T1 cells were treated with (a) Control, (b) 3.8, (c) 7.71, (d) 15.14 μg mL–1 IFB and (e) Graphical representation of Bax expression Cells were processed with Immunohistochemistry to detect Bax as described in Methods. Semiquantified Bcl-2 expression with ImageJ. Bar: 150 μm. Values area means±SD, n = 5 Replicates, *p-value<0.05 then indicate statistically significant differences in comparison to control |
The arrows point to the positive-staining cells. The Bcl-2 expression semi quantification of control (1% ethanol), 3.8 (¼ of IC50), 7.71 (½ of IC50) and 15.14 (IC50) μg mL–1 IFB-treated 4T1 cells were 9.72, 6.67, 2.83 and 1.31% (Fig. 3e). These results showed that the treatment of 3.8, 7.71 and 15.14 μg mL–1 IFB decreased the expression of Bcl-2 until 22, 65 and 91%, respectively. The test results at each test concentration of all fractions showed a significant difference with the control group (p<0.05).
Bax expressions of IFB-treated 4T1 cells: The expression of Bax can be seen in Fig. 4. Expressions of Bax were indicated by the presence of Bax monoclonal antibodies bonds, detected in brown color in the cytoplasm and membrane of 4T1 cells (Fig. 4a-d). The arrows point to the positive-staining cells.
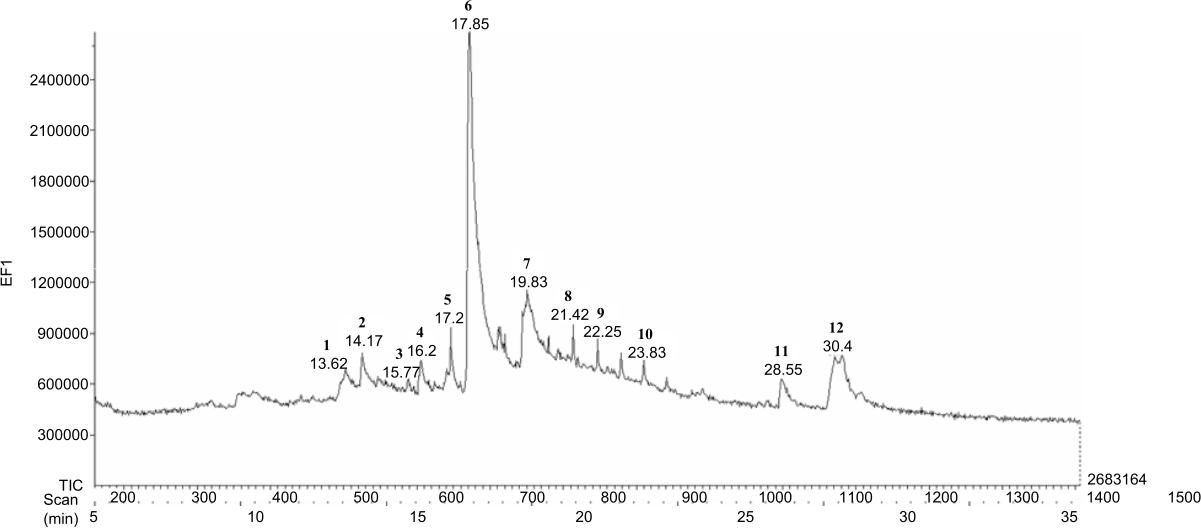 |
| Fig. 5: | GC chromatogram of IFB of P. alata bark |
| Table 2: Compounds have been identified from IFB of P. alata bark | |||||
| Retention | Abundance | ||||
| Peak | time (min) | (%) | Compound | Activity | References |
| 1 | 13.62 | 6.3 | Germacrene | Cytotoxic potential against LNCaP cancer cells | Srivilai et al.19 |
| 2 | 14.17 | 7.13 | Methyl eugenol | Cytotoxic against retinoblastoma RB355 cells | Meng et al.20 |
| 3 | 15.77 | 2.15 | Palmitic acid | Cytotoxic potential against neuroblastoma cells SH-SY5Y | Ng et al.21 |
| 4 | 16.2 | 6.79 | 1-eicosene | Cytotoxic | Yeo et al.22 |
| 5 | 17.2 | 8.2 | 1,3-dimethylbutyl acetate | Cytotoxic | Zhu et al.23 |
| 6 | 17.85 | 26.12 | Beta-caryophyllene | Cytotoxic against MCF-7, DLD-1 and L-929 human tumor | Legault and Pichette24 |
| 7 | 19.83 | 9.65 | Beta-bisabolene | Cytotoxic against HCT-116 | da Silva et al.25 |
| 8 | 21.42 | 9.33 | Hexadecanoic acid | Cytotoxic against Hep G2 | Gómez-Lechón et al.26 |
| 9 | 22.25 | 6.39 | Beta-sesquiphellandrene | Cytotoxic against KG-1 and HL-60 cells | Tyagi et al.27 |
| 10 | 23.83 | 5.79 | m-aminobenzaldehyde oxime | Cytotoxic | Canario et al.28 |
| 11 | 28.55 | 4.96 | Z-13-Octadecen-1-yl acetate | Cytotoxic | Gupta et al.29 |
| 12 | 30.4 | 6.43 | Thionodecalactone | Cytotoxic | Xu et al.30 |
The Bax expression semi quantification of control (1% ethanol), 3.8 (¼ of IC50), 7.71 (½ of IC50) and 15.14 (IC50) μg mL–1 IFB-treated 4T1 cells were 1.13, 3.17, 6.38 and 8.5% (Fig. 4e). These results showed that the treatment of 3.8, 7.71 and 15.14 μg mL–1 IFB increased the expression of Bax until 16, 45 and 65%, respectively. The test results at each test concentration of all fractions showed a significant difference with the control group (p<0.05).
GC-MS analysis of IFB: The pharmacological activity of a plant will be determined by its metabolites. Therefore we analyzed the compounds contained in IFB using GC-MS to determine the metabolite profile. There are a total of 12 compounds identified in the IFB, representing about 100% of the total compound. The major components of IFB are beta-caryophyllene (26.12%), beta-bisabolene (9.65), hexadecanoic acid (9.33) and 1,3-dimethylbutyl acetate (8.20%) (Fig. 5 and Table 2). The results of the literature search show that almost all chemical compounds contained in IFB have cytotoxic activity. Likewise, we studied that there are taraxerol compounds that have antioxidant and cytotoxic activity on MCF-7 cells and T47D cell lines. As a result, the IFB on cancer and normal cells in concern with the metabolic profiles needed to be explored further.
DISCUSSION
Although P. alata has been widely known to have pharmacological activity, in anticancer tests, it has not been widely studied. This is a critical point to prove P. alata has an activity to maintain health and has anticancer activity, so our focus is to evaluate the evidence-based cytotoxic properties of metastatic breast cells, namely 4T1 cells and normal cells, namely Vero cells. The results of the study prove that cisplatin can inhibit the growth of 4T1 cells, cisplatin is reported to be able to induce Bcl-2 downregulation in T47D breast cancer cells31. It can affect DNA replication and transcription, resulting in stopping the cell cycle in the S phase and triggering apoptosis32. Downregulation of Bcl-2 (anti-apoptotic protein) will decrease cell survival and increase sensitivity to chemotherapy agents33. From the statistical test results, Cisplatin and IFB did not have a significant difference. The study indicated that IFB has almost equal power in inhibiting the growth of 4T1 cells so that IFB is used for the next test phase, especially in its role in apoptosis.
Apoptosis plays a massive role in tumor development and progression by eliminating the cancerous and preneoplastic cells form. Morphological analysis of cells demonstrating IFB induces typical apoptotic characteristics such as nuclear shrinkage and fragmentation34. As apoptosis is modulated by a group of anti-apoptotic and pro-apoptotic regulators, we examined the expression of these key regulatory proteins in apoptosis in the tumor sections. Bax promotes the release of cytochrome C into the cytosol from the mitochondria35. The antiapoptotic proteins, such as Bcl-2 preserve the integrity of the mitochondria. This blocks the release of cytochrome C that activates the effectors of apoptosis36. Since Bcl-2 functions by forming a heterodimer with its pro-apoptotic partner Bax, the Bcl-2/Bax ratio is proportional to the relative sensitivity or resistance of the cells to various apoptotic stimuli37. By comparing with vehicle control, our work indicated that IFB specifically and significantly increased the number of cells that expressed Bax and decreased the number of cells that expressed Bcl-2. These data show that increased numbers of cells underwent apoptosis after IFB treatment, suggesting that IFB inhibited breast tumor growth by induction of apoptosis. Besides, these data indicate that the molecular mechanism occurs through IFB induced mitochondria-mediated apoptosis. Apoptosis occurs by inhibiting the expression of the anti-apoptotic factor that releases cytochrome C from the mitochondria and activation of caspase-3. Besides, the results strongly supported the hypothesis of the mitochondrial apoptotic pathway involvement.
The chemical compounds in IFB are responsible for cytotoxic activity (Table 2), lowering BCL-2 levels and increasing Bax levels. The class of compounds detected from GC-MS in IFB is a polyketide compound. Many polyketide compounds are known to have cytotoxic activity37-42. Germacrene has cytotoxic activity on LNCaP cancer cells19, besides that according to the study of Govindam et al.43 reported that germacrene isolated from the soft coral Lobophytum pauciflorum has an activity to inhibit human epidermoid carcinoma A431 cells by inhibiting the activation of NF-κB activation by IKKa/b. NF-κB has an essential role as transcription factors that regulate the expression of anti-apoptotic genes44. Beta-caryophyllene is found as a major compound (26.12%) in IFB. Major compounds have a considerable role in pharmacological activity45. Park et al.46 reported that Beta-caryophyllene isolated from Psidium guajava leaves could induce apoptosis in PC-3 and MCF-7 cells by inducing Bax. Bax induces the release of Cytochrome C from mitochondria explicitly, Cytochrome C will form a complex with Protease Activating Factor-1 (Apaf1) and Procaspase-9 apoptotic which are called apoptosome complexes47. The apoptosome will activate caspase-9 and there will be activation of caspase 6 and caspase 7 and induce apoptosis48.
Besides apoptosis, cytotoxic effects can also occur due to the induction of cell cycle49. Abdel Bar et al.50 reported that methyl eugenol has a cytotoxic effect on human cervical cancer cells (HeLa, C4-1, MEK-180, C33A) by inducing G2/M cell cycle arrest. Beta-bisabolene has also been reported to be induced G2/M cell cycle arrest and apoptosis, elevated levels of cell cycle regulatory proteins p53 and p21 and lowered levels of G2/M checkpoint proteins Cdc2 and Cyclin B1 in melanoma cells51. The cytotoxic effect can occur due to the various chemical compounds possessed by IFB; the cytotoxic effect can occur due to the synergistic effect of the compounds it has. Either through the process of apoptosis or induction of the cell cycle. However, all these things need to be proven more deeply to ensure the cytotoxic mechanism against 4T1 cells.
CONCLUSION
Pterygota alata has empirically beneficial as traditional medicine, especially on the bark. The bark of P. alata was fractionated using five types of solvents. The insoluble fraction of n-butanol (IFB) had the highest cytotoxic activity against 4T1 cells and was less toxic to normal cells. IFB induces apoptosis in 4T1 cells through the intrinsic apoptotic pathway by decreasing Bcl-2 expression and increasing Bax expression. The cytotoxic IC50 of IFB on 4T1 cells was 15.14±2.81 μg mL–1. GC-MS spectrum analysis showed that IFB had five main compounds, namely beta-caryophyllene (26.12%), beta-bisabolene (9.65), hexadecanoic acid (9.33) and 1,3-dimethylbutyl acetate (8.20%), which were proven to have cytotoxic activity.
SIGNIFICANCE STATEMENT
This research was conducted to investigate the active compounds in P. alata bark which have cytotoxic activity with apoptotic mechanisms. This study will provide new evidence and theory to researchers about the use of P. alata medicinal plants for the treatment of breast cancer.
ACKNOWLEDGMENT
This research article is part of the doctoral thesis and this research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
REFERENCES
- Soteriades, E.S., J. Kim, C.A. Christophi and S.N. Kales, 2019. Cancer incidence and mortality in firefighters: A state-of-the-art review and meta-َanalysis. Asian Pac. J. Cancer Prev., 20: 3221-3231.
CrossRefDirect Link - Hanif, N., A. Hermawan and E. Meiyanto, 2019. Caesalpinia sappan L. ethanolic extract decrease intracellular ROS level and senescence of 4T1 breast cancer cells. Indonesian J. Cancer Chemoprevention, 10: 16-23.
CrossRefDirect Link - Mattiuzzi, C. and G. Lippi, 2019. Current cancer epidemiology. J. Epidemiol. Global Health, 9: 217-222.
CrossRefDirect Link - Gao, X., X. Nan, Y. Liu, R. Liu and W. Zang et al., 2020. Comprehensive profiling of BRCA1 and BRCA2 variants in breast and ovarian cancer in Chinese patients. Human Mutat., 41: 696-708.
CrossRefDirect Link - Muflikhasari, H.A., R.I. Jenie, R.A. Susidarti and E. Meiyanto, 2019. Growth inhibitory property of pentagamavunone-0 (PGV-0) on 4T1 cells under stress condition: 2D and 3D culture model. Indonesian J. Cancer Chemoprevention, 10: 149-158.
Direct Link - Novitasari, D., S. Handayani and R.I. Jenie, 2018. Ethanolic extract of Hedyotis corymbosa L. inhibits migration and MMP-9 activity on metastatic breast cancer cells. Indonesian J. Cancer Chemoprevention, 9 : 16 -22.
CrossRefDirect Link - Rollando, R., M. Engracia, E. Monica and S. Siswadi., 2020. Immunomodulatory activity test of syrup dosage form of combination Phyllantus niruri Linn. and Sterculia quadrifida R.Br. extract. Int. J. Res. Pharm. Sci., 11: 191-199.
CrossRefDirect Link - El-Sherei, M.M., A.Y. Ragheb, S.A. Mosharrafa, M.M. Marzouk, M.E.S. Kassem and N.A.M. Saleh, 2018. Pterygota alata (Roxb.) R. Br.: Chemical constituents, anti-hyperglycemic effect and anti-oxidative stress in alloxan-induced diabetic rats. J. Mater. Environ. Sci., 9 : 245-255.
CrossRefDirect Link - Lin, L., Z. Song and H. Xu, 2010. A new phenylpropanoid galactoside and other constituents from Pterygota alata (Roxb.) R. Brown. Biochem. Syst. Ecol., 38: 1238-1241.
CrossRefDirect Link - de Souza, S.A., T.M.G. da Silva, E.M.S. da Silva, C.A. Camara and T.M.S. Silva, 2018. Characterisation of phenolic compounds by UPLC‐QTOF‐MS/MS of geopropolis from the stingless bee Melipona subnitida (jandaíra). Phytochem. Anal., 29: 549-558.
CrossRefDirect Link - Coates, J.M., J.M. Galante and R.J. Bold, 2010. Cancer therapy beyond apoptosis: Autophagy and anoikis as mechanisms of cell death. J. Surg. Res., 164: 301-308.
CrossRefDirect Link - Mahib, M.R., S. Hosojima, H. Kushiyama, T. Kinoshita, T. Shiroishi, T. Suda and K. Tsuchiya, 2020. Caspase‐7 mediates caspase‐1‐induced apoptosis independently of bid. Microbiol. Immunol., 64: 143-152.
CrossRefDirect Link - Walkley, C.R. and B.T. Kile, 2019. Cell death following the loss of ADAR1 mediated A-to-I RNA editing is not effected by the intrinsic apoptosis pathway. Cell Death Dis., Vol. 10.
CrossRefDirect Link - Lu, Z., H. Zhou, S. Zhang, W. Dai and Y. Zhang et al., 2020. Activation of reactive oxygen species‐mediated mitogen‐activated protein kinases pathway regulates both extrinsic and intrinsic apoptosis induced by arctigenin in Hep G2. J. Pharm. Pharmacol., 72: 29-43.
CrossRefDirect Link - Bumbat, M., M. Wang, W. Liang, P. Ye, W. Sun and B. Liu, 2020. Effects of Me2SO and trehalose on the cell viability, proliferation, and Bcl-2 family gene (BCL-2, BAX, and BAD) expression in cryopreserved human breast cancer cells. Biopreservation Biobanking, 18: 33-40.
CrossRefDirect Link - Steenbrugge, J., N.V. Elst, K. Demeyere, O.D. Wever and N.N. Sanders et al., 2019. Comparative profiling of metastatic 4T1- vs. non-metastatic Py230-based mammary tumors in an intraductal model for triple-negative breast cancer. Front. Immunol., Vol. 10.
CrossRefDirect Link - Wu, H., L. Chen, F. Zhu, X. Han, L. Sun and K. Chen, 2019. The cytotoxicity effect of resveratrol: Cell cycle arrest and induced apoptosis of breast cancer 4T1 cells. Toxins, Vol. 11.
CrossRefDirect Link - Shen, C.F., C. Guilbault, X. Li, S.M. Elahi, S. Ansorge, A. Kamen and R. Gilbert, 2019. Development of suspension adapted vero cell culture process technology for production of viral vaccines. Vaccine, 37: 6996-7002.
CrossRefDirect Link - Srivilai, J., N. Khorana, N. Waranuch, W. Wisuitiprot, N. Suphrom, A. Suksamrarn and K. Ingkaninan, 2016. Germacrene analogs are anti-androgenic on androgen-dependent cells. Nat. Prod. Commun., Vol. 11.
CrossRefDirect Link - Meng, B., H. Ii, W. Qu and H. Yuan, 2018. Anticancer effects of gingerol in retinoblastoma cancer cells (RB355 cell line) are mediated via apoptosis induction, cell cycle arrest and upregulation of PI3K/Akt signaling pathway. Med. Sci. Monit., 24: 1980-1987.
CrossRefDirect Link - Ng, Y.W. and Y.H. Say, 2018. Palmitic acid induces neurotoxicity and gliatoxicity in SH-SY5Y human neuroblastoma and T98G human glioblastoma cells. PeerJ, Vol. 6.
CrossRefDirect Link - Yeo, S.K., A.Y. Ali, O.A. Hayward, D. Turnham, T. Jackson, I.D. Bowen and R. Clarkson, 2016. β-bisabolene, a sesquiterpene from the essential oil extract of opoponax (Commiphora guidottii), exhibits cytotoxicity in breast cancer cell lines. Phytother. Res., 30: 418-425.
CrossRefDirect Link - Zhu, N.L., C. Tang, C. Xu, C.Q. Ke and G. Lin et al., 2019. Cytotoxic germacrane-type sesquiterpene lactones from the whole plant of Carpesium lipskyi. J. Nat. Prod., 82: 919-927.
CrossRefDirect Link - Legault, J. and A. Pichette, 2007. Potentiating effect of β-caryophyllene on anticancer activity of α-humulene, isocaryophyllene and paclitaxel. J. Pharm. Pharmacol., 59: 1643-1647.
CrossRefPubMedDirect Link - da Silva, J., E. Andrade, L. Barreto, N. da Silva, A. Ribeiro, R. Montenegro and J. Maia, 2017. Chemical composition of four essential oils of eugenia from the brazilian amazon and their cytotoxic and antioxidant activity. Medicines, Vol. 4.
CrossRefDirect Link - Gómez-Lechón, M.J., M.T. Donato, A. Martínez-Romero, N. Jiménez, J.V. Castell and J.E. O’Connor, 2007. A human hepatocellular in vitro model to investigate steatosis. Chem. Biol. Interact., 165: 106-116.
CrossRefDirect Link - Tyagi, A.K., S. Prasad, W. Yuan, S. Li and B.B. Aggarwal, 2015. Identification of a novel compound (β-sesquiphellandrene) from turmeric (Curcuma longa) with anticancer potential: Comparison with curcumin. Invest. New Drugs, 33: 1175-1186.
CrossRefDirect Link - Canário, C., S. Silvestre, A. Falcão and G. Alves, 2018. Steroidal oximes: Useful compounds with antitumor activities. Curr. Med. Chem., 25: 660-686.
CrossRefDirect Link - Gupta, A., B.S. Kumar and A.S. Negi, 2013. Current status on development of steroids as anticancer agents. The J. Steroid Biochem. Mol. Biol., 137: 242-270.
CrossRefDirect Link - Xu, X.Y., P. Sun, D.A. Guo, X. Liu, J.H. Liu and L.H. Hu, 2015. Cytotoxic sesquiterpene lactone dimers isolated from Inula japonica. Fitoterapia, 101: 218-223.
CrossRefDirect Link - Oladimeji, A.O., O. Babatunde, R.T. Musa, F.A. M'civer, A.T. Lawal and I.A. Ogunwande, 2016. GC-MS analysis and cytotoxic activity of essential oils from the leaves of Abrus precatorius L. gaertn. Asian Pac. J. Trop. Dis., 6: 372-375.
CrossRefDirect Link - Taher, M., W.M.N.H.W. Salleh, S.I. Alkhamaiseh, F. Ahmad, M.F. Rezali, D. Susanti and C.M. Hasan, 2020. A new xanthone dimer and cytotoxicity from the stem bark of Calophyllum canum. Zeitschrift für Naturforschung C,.
CrossRefDirect Link - Hariono, M., R. Rollando, J. Karamoy, P. Hariyono and M. Atmono et al., 2020. Bioguided fractionation of local plants against matrix metalloproteinase9 and its cytotoxicity against breast cancer cell models: In silico and in vitro study. Molecules, Vol. 25.
CrossRefDirect Link - Roihatul Mutiah, Muhammad Ragib Mustofa, Yen Yen Ari Indrawijaya, Abdul Hakim, Rahmi Annisa, Nurlaili Susanti, Ach Nashichuddin, Muhammad Zainuddin 2019. Exposure of Murattal Al-Quran Audio Enhances Cisplatin Activity on Growth Inhibition and Cell Cycle Modulation on Hela Cells Indonesian J. Cancer Chemoprevention, 10: 71-79.
CrossRefDirect Link - Kønig, S.M., V. Rissler, T. Terkelsen, M. Lambrughi and E. Papaleo, 2019. Alterations of the interactome of Bcl-2 proteins in breast cancer at the transcriptional, mutational and structural level. PLoS Comput Biol., Vol. 15.
CrossRefDirect Link - Husnaa, U., N.P.L. Laksmiani, R.A. Susidarti and E. Meiyanto, 2017. Ethyl acetate fraction of Caesalpinia sappan L. enhances cisplatin’s cytotoxicity on hela cells via G1 and S arrest through p53 expression. Indonesian J. Cancer Chemoprevention, 8: 51-60.
CrossRefDirect Link - Fitriasih, F., S.M. Komariyah, F. Sandra, N. Pratiwi and D.N. Hidayati, 2019. Mangifera indica L. leaves extract induced intrinsic apoptotic pathway in MCF-7 cells by decreasing Bcl-2 expression and inducing bax expression. Indonesian J. Cancer Chemoprevention, 10: 1-6.
CrossRefDirect Link - Liang, X., C. Xu, X. Cao and W. Wang, 2019. Isovitexin suppresses cancer stemness property and induces apoptosis of osteosarcoma cells by disruption of the DNMT1/miR-34a/Bcl-2 axis. Cancer Manage. Res., 11: 8923-8936.
CrossRefDirect Link - Qin, N., S. Lu, N. Chen, C. Chen and Q. Xie et al., 2019. Yulangsan polysaccharide inhibits 4T1 breast cancer cell proliferation and induces apoptosis in vitro and in vivo. Int. J. Biol. Macromol., 121: 971-980.
CrossRefDirect Link - Feng, Y., Q. Guo and B. Shao, 2019. Cytotoxic comparison of macrolide antibiotics and their chlorinated disinfection byproduct mixtures. Ecotoxicol. Environ. Saf., Vol. 182.
CrossRefDirect Link - Niu, S., M. Xia, M. Chen, X. Liu and Z. Li et al., 2019. Cytotoxic polyketides isolated from the deep-sea-derived fungus Penicillium chrysogenum MCCC 3A00292. Marine Drugs, Vol. 17.
CrossRefDirect Link - Narmani, A., R.B. Teponno, S.E. Helaly, M. Arzanlou and M. Stadler, 2019. Cytotoxic, anti-biofilm and antimicrobial polyketides from the plant associated fungus Chaetosphaeronema achilleae. Fitoterapia, Vol. 139.
CrossRefDirect Link - He, Y., R. Wang, B. Huang, Q. Dai and J. Lin, 2020. Pholiotone A, a new polyketide derivative from Pholiota sp. Nat. Prod. Res., 34: 1957-1961.
CrossRefDirect Link - Govindam, S.V.S., Y. Yoshioka, A. Kanamoto, T. Fujiwara, T. Okamoto and M. Ojika, 2012. Cyclolobatriene, a novel prenylated germacrene diterpene, from the soft coral Lobophytum pauciflorum. Bioorg. Med. Chem., 20: 687-692.
CrossRefDirect Link - Zheng, H., X. Wang, Y. Zhang, L. Chen, L. Hua and W. Xu, 2019. Pien-Tze-Huang ameliorates hepatic fibrosis via suppressing NF-κB pathway and promoting HSC apoptosis. J. Ethnopharmacol., Vol. 244.
CrossRefDirect Link - Arul, S., H. Rajagopalan, J. Ravi and H. Dayalan, 2020. Beta-caryophyllene suppresses ovarian cancer proliferation by inducing cell cycle arrest and apoptosis. Anticancer Agents Med. Chem., 20: 1530-1537.
CrossRefDirect Link - Park, K.R., D. Nam, H.M. Yun, S.G. Lee and H.J. Jang et al., 2011. β-Caryophyllene oxide inhibits growth and induces apoptosis through the suppression of PI3K/AKT/mTOR/S6K1 pathways and ROS-mediated MAPKs activation. Cancer Lett., 312: 178-188.
CrossRefDirect Link - Ryu, N.H., K.R. Park, S.M. Kim, H.M. Yun and D. Nam et al., 2012. A hexane fraction of guava leaves (Psidium guajava L.) induces anticancer activity by suppressing AKT/mammalian target of rapamycin/ribosomal p70 S6 kinase in human prostate cancer cells. J. Med. Food, 15: 231-241.
CrossRefDirect Link - Kim, S.W., S.M. Kim, H. Bae, D. Nam and J.H. Lee et al., 2013. Embelin inhibits growth and induces apoptosis through the suppression of Akt/mTOR/S6K1 signaling cascades. Prostate, 73: 296-305.
CrossRefDirect Link - Lee, J.H. and J.M. Berger, 2019. Cell cycle-dependent control and roles of DNA topoisomerase II. Genes, Vol. 10.
CrossRefDirect Link - Bar, F.M.A., M.A. Khanfar, A.Y. Elnagar, F.A. Badria and A.M. Zaghloul et al., 2010. Design and pharmacophore modeling of biaryl methyl eugenol analogs as breast cancer invasion inhibitors. Bioorg. Med. Chem., 18: 496-507.
CrossRefDirect Link - Abdjul, D.B., S.I. Kanno, H. Yamazaki, K. Ukai and M. Namikoshi, 2016. A dimeric urea of the bisabolene sesquiterpene from the Okinawan marine sponge Axinyssa sp. inhibits protein tyrosine phosphatase 1B activity in Huh-7 human hepatoma cells. Bioorg. Med. Chem. Lett., 26: 315-317.
CrossRefDirect Link