
Heba F. Gomaa
Department of Zoology, Faculty of Science, Ain-shams University, Cairo, Egypt
LiveDNA: 20.20322
Khaled G. Abdel-Wahhab
Department of Medical Physiology, National Research Centre, Giza, Egypt
LiveDNA: 20.34876
Mahmoud Ashry
Department of Zoology, Faculty of Science, Al-Azhar University, Assuit, Egypt
LiveDNA: 20.12591
Laila K. Hassan
Department of Dairy, National Research Centre, Cairo, Egypt
Fathia A. Mannaa
Department of Medical Physiology, National Research Centre, Giza, Egypt
Pakistan Journal of Biological Sciences
Year: 2021 | Volume: 24 | Issue: 1 | Page No.: 112-121







ABSTRACT
Background and Objective: Pyrethroidsare a group of synthetic pesticides similar to the natural pesticide pyrethrum, which is produced by chrysanthemum flowers. Bifenthrin is one of the pyrethroids that are widely used pesticide in households and to control crop vectors. The main goal of this work was to investigate the possible ameliorating effect of Costus Ethanolic Extract (CEE) against neurotoxicity induced by bifenthrin in adult-male rats. Materials and Methods: Rats were arranged randomly to 4 groups (8 rats each) as next. Group 1) control rats orally received 0.5 mL water for consecutive 30 days; group 2) healthy rats orally received CEE (200 mg kg) for consecutive thirty days; group 3) rats treated orally with 7 mg kg–1 day–1 bifenthrin for consecutive 30 days and group 4) included rats treated with bifenthrin for consecutive 30 days followed by administration with CEE another consecutive 30 days. Results: The results showed that CEE succeeded to decline the neurotoxicity-induced by bifenthrin; this was evidenced by the significant reduction in TNF-α, IL- 1β, MDA and nitric oxide levels in cortex, hippocampus and striatum concomitant with marked improvement in the values of GSH, dopamine, serotonin, AChE-ase, SOD, GPx and catalase that were diminished by bifenthrin intoxication. CEE improved also cognitive impairment and the deficits in motor coordination induced by bifenthrin. Conclusion: CEE was found successful, to a great extent, to counteract the bifenthrin-induced brain oxidative stress and neurochemical deteriorations and possesses a protective potential against brain-induced neurotoxicity. Therefore, it may be a promising supplement for the amelioration of BF-neurotoxicity.
PDF Abstract XML References Citation
How to cite this article
Heba F. Gomaa, Khaled G. Abdel-Wahhab, Mahmoud Ashry, Laila K. Hassan and Fathia A. Mannaa, 2021. The Estimation of the Anti-neurotoxic Effect of Costus Ethanolic Extract against Bifenthrin-Intoxication in Male Rats. Pakistan Journal of Biological Sciences, 24: 112-121.
DOI: 10.3923/pjbs.2021.112.121
URL: https://scialert.net/abstract/?doi=pjbs.2021.112.121
DOI: 10.3923/pjbs.2021.112.121
URL: https://scialert.net/abstract/?doi=pjbs.2021.112.121
INTRODUCTION
Bifenthrin, a non-alpha cyano-pyrethroid insecticide, that is from the most popularly used insecticides in pest control in the field of agriculture due to its properties, such as it has very low vapor pressure and good stability to hydrolysis and photolysis, in addition to that it induces irritation to eyes and skin1. Generally, neurotoxicity is identified as detrimental stress on the nervous system that resulted from a biological, chemical, or physical agent2. Many environmental pollutants, including pesticides, are described to disturb the balance of free radical’s formation and removal thus resulting in oxidative stress. Different cellular mechanisms are included in modulating oxidative stress and restoring the damaged macromolecules. The mechanism that behaved by most pesticides includes initiation of oxidative stress that damages membranes’ lipids, proteins and DNA1. The participation of free radicals in neurodegenerative diseases, like Alzheimer’s diseases and Parkinson’s disease, is well reported. Pyrethroid compounds are considered to stimulate liberation and release of Reactive Oxygen Species (ROS) leading to oxidative injuries in the living organisms as a consequence of their exposure3.
Bifenthrin is one of the most commonly used pesticides. It is a synthetic pyrethroid (type I) that is used at a large scale for agricultural applications and public health management4. Bifenthrin is a cyclopropane carboxylate ester of alcohol. It has strong insecticidal properties due to the ability to alter the functioning of insect nerves through altering the kinetics of charge-sensitive sodium channels. It was found to alter gait and other motor functions in rodents5.
Herbal medicines are being used throughout the world more than 4000 years ago for the management of many diseases as they contain beneficial chemical constituents. In essence, medical potentials of plants belong in their phytochemical constituents that perform exact pharmacological actions when they are ingested into the human body.
Extracts of costus exhibit central nervous system depressant activities and so can be used as a smooth muscle relaxant and to enhance antispasmodic activities6. The use of costus in treatment exhibits a strong inhibitory efficacy against genotoxicity alterations initiated by streptozotocin (STZ), thus the costus extracts might behave anti-genotoxic property, evidencing the protective ability of this compounds7. Saussurea costus roots performed many pharmacological effects such as anodyne, anti-bacterial, anti-spasmodic, etc. They were used internally in the relief of pain and distension of abdomen, pains of the chest, liver problems and jaundice, pain of gall bladder, constipation associated with energy stagnation and asthma8. Many plants include potent bioactive compounds that can prevent and treat most oxidative related stresses9 and have often been used in traditional medicine10. Therefore, this study was conducted to investigate the ameliorative role of Costus Ethanolic Extract (CEE) against the adverse effect of bifenthrin-induced neurotoxicity.
MATERIALS AND METHODS
Study area: This study was carried out during the period from January-May, 2019 at the Department of Medical Physiology and Animal Colony, National Research Centre, Egypt.
Plant and extraction: Costus (Saussurea costus) roots were obtained from a local supplier (Abdel-Rahman Harraz, Bab el-Khalk Zone, Cairo, Egypt); then identified and authenticated by scientific botanists and found carrying a taxonomic serial number 780691. The ethanolic extract of dry powdered roots was carried out according to the modified method of the previous menthod11 in brief, roots' powder was soaked in absolute ethanol (1:10 w/v) at room temperature for 48 h under continuous stirring. Extraction was performed in triplicate; finally, the mixtures were combined then filtered through sterile filter papers (Whatman number 42, England). The solvent was evaporated using a rotary evaporator and then the extract was stored at -20°C until further use.
Extract yield: The whole filtrate was poured in an empty quick fit round bottom flask with known weight (W1) and then weighed again (W2) after solvent evaporation; finally, the yield percentage was calculated from the following formula12:
 |
where, W1 is the weight of empty and dry quick fit flask in grams, W2 is the weight of the flask after rotary evaporation in grams; W3 is the weight of the used crude powdered herb in grams12.
Total phenolic content: The total phenolic content of the CEE was performed by dissolving 5 mg of the extract in a 10 mL mixture of acetone and water (6:4 v/v). Then, a sample of 0.2 mL was mixed with 1.0 mL of Folin-Ciocalteu reagent (10 fold diluted) and 0.8 mL of sodium carbonate solution (7.5%). After 30 min at room temperature, the absorbance was measured at 765 nm using Cary 100 UV-Vis spectrophotometer. Total phenolic content (as catechin equivalent) was estimated by using standard curve13.
DPPH radical scavenging activity: The capacity of antioxidants of CEE to quench DPPH radical was determined as previously14. A 0.2 mL of extract/methanol solution (200 ppm w/v) was completed to 4 mL by methanol and then 1 mL DPPH solution (6.09×10–5 mol L–1 in methanol) was added. Control or reference was prepared by adding 1 mL of DPPH solution to 4 mL methanol. After 10 min at room temperature, the absorbance of both tested mixture and control sample was measured at 516 nm. Triplicate measurements were made and the percentage of radical scavenging activity was calculated according to the equation14.
Reducing power: Reducing power of CEE was determined according to the method described by previous method15 with some modifications. From CEE extract and ascorbic acid, 0.5 mL of different concentrations (50, 100, 200, 400 and 800 μg mL–1) was mixed with 2.5 mL phosphate buffer (pH 7.4) and 2.5 mL potassium ferricyanide (0.1 M); the mixture was kept at 50°C in a water bath for 20 min; then after cooling, 2.5 mL of trichloroacetic acid (10%) was added and centrifuged at 3000 rpm for 10 min. Immediately, 2.5 mL of the upper layer of the solution was mixed with 2.5 mL distilled water and 0.5 mL of a freshly prepared ferric chloride solution (40% w/v). The absorbance of the sample and standard was measured at 700 nm. Control blank included mixture components without sample or standard.
HPLC analysis of phenolic constituents: The HPLC analysis was carried out using an Agilent 1260 series. The separation was carried out using a Kromasil C18 column (4.6×250 mm i.d., 5 μm). The mobile phase consisted of water (A) and 0.05% trifluoroacetic acid in acetonitrile (B) at a flow rate of 1 mL min–1. The mobile phase was programmed consecutively in a linear gradient as follows: 0 min (82% A); 0-5 min (80% A); 5-8 min (60% A); 8-12 min (60% A); 12-15 min (85% A) and 15-16 min (82% A). The multi-wavelength detector was monitored at 280 nm. The injection volume was 10 μL for each of the sample solutions. The column temperature was maintained at 35°C16,17.
Animals and experimental design: Total 32 adult male albino rats (150-200 g) were obtained from the animal colony, National Research Centre where the animals were maintained under temperature (25±1°C) and light (12/12 hrs light/dark cycle) controlled conditions on free access to food and water for a week before starting the experiment for acclimatization. The rats received human care in compliance with the standard institution’s criteria for the care and use of experimental animals according to the procedures approved by the Ethics Committee of the National Research Centre (FWA 00014747) that follows the recommendations of the National Institutes of Health Guide for Care and Use of Laboratory Animals.
After the animals were being acclimatized with experimental room conditions, they were divided randomly into 4 groups (8 animals each): group 1) healthy control rats orally received 0.5 mL water for consecutive 30 days, group 2) healthy rats orally received CEE (200 mg kg–1, dissolved in water) for consecutive 30 days, group 3) rats were treated orally with 7mg kg–1 day–1 of bifenthrin (purity>98%, Sigma, USA) dissolved in olive oil for consecutive 30 days16 and group 4) rats were treated with bifenthrin for 30 days followed by administration with CEE another consecutive 30 days.
At the end of the treatment period, all animals were subjected to rotarod performance and spatial memory test (Morris water maze) for behavioral studies, then fasted overnight, anesthetized with sodium pentobarbital (200 mg kg–1 diluted 1:3 with phosphate-buffered saline, IP)18 and rapidly sacrificed by decapitation. On ice-cooled glass plate, their brain areas (cortex, striatum and hippocampus) were immediately anatomized and dissected out, washed in ice-cold saline, then stored at -80°C till assessments. The specific weight of each brain area was homogenized (Ultrasonically) in Tris-HCl buffer (pH 7.4) to give 10% w/v homogenate. The homogenate was centrifuged at 5000 rpm at 4°C for 10 min and the supernatant was stored at -20°C until analysis. This supernatant was used for the determination of oxidative stress markers, cytokines and acetylcholinesterase.
Rotarod performance: The motor coordination and balance of animals were studied using a rotarod (IMCORP, Ambala, India) as previously described procedure19. A set of animals from each group was trained to stay on the rotating rod until it achieved the criterion of staying on the rod for 60 sec. Final observations were conducted by placing the rats on the rotating rod (25 rpm) with 180 sec as the cut-off time. The time of fall from the rotating rod was determined as a measure of motor coordination and scoring was carried out by a person blind to the treatment condition. Each rat was subjected to three consecutive trials after a gap of 5 min.
Spatial memory test (Morris water maze): The Morris Water Maze (MWM) test was used to assess spatial learning in rats following the procedure described20. The spatial memory of animals was manifested by the time spent in the quadrant where the platform was placed.
Determination of acetylcholinesterase activity: Acetylcholinesterase (AchE) activity was determined by the modified method21. The hydrolysis of acetylthiocholine iodide by acetylcholinesterase yields thiocholine, which reacted with 5, 5’-dithiobis-(2-nitrobenzoic acid), reducing it to thionitrobenzoic acid, whose yellow color was read at 412 nm.
Biogenic amines determination: A certain weight of each brain region was homogenized in 0.1 M perchloric acid (containing 3, 4-dihydroxybenzylamine) at a final concentration of 25 ng mL–1 then centrifuged 10 min at 3600 rpm g–1. The obtained supernatant was then filtered through 0.25 mm nylon filters, Millipore, USA (specific for biogenic amines determination). Dopamine and serotonin levels were determined in the concerned brain areas using HPLC (Waters, Melford, USA) by injection of 20 μL sample supernatant into the injector port and using an electrochemical detector22. with a high-pressure isocratic pump, a sample injector valve, a C-18 reverse phase column (250×4 mm, particle size 5 μm) and an electrochemical detector (464 Pulsed electrochemical detector).
Estimation of oxidative stress markers: Superoxide dismutase (SOD), glutathione peroxidase (GPx) and catalase (CAT) activities and also the levels of reduced glutathione (GSH) and nitric oxide (NO) were determined using reagent kits obtained from Biodiagnostic Co., Giza, Egypt. Lipid peroxidation level was estimated chemically according to the method described by the previous method23. On the base of MDA reaction with thiobarbituric acid (TBA) which forms a pink complex that can be measured photometrically. In this method, 0.5 mL cortex, striatum and hippocampus homogenate supernatant were added to 4.5 mL working reagent [0.8 g TBA was dissolved in 100 mL perchloric acid 10% and mixed with 20% trichloroacetic acid in volume ratio 1-3, respectively]. In boiling and shaking water bath, the sample-reagent mixture was left for 20 min, then carried out to cool at room temperature and centrifuged for 5 min at 3000 rpm. The absorbance of the clear pink supernatant was measured photometrically at 535 nm against a reagent blank (0.5 mL distilled water +4.5 mL working reagent). The MDA level was calculated in nmol MDA g–1 tissue according to the following formula23:
MDA (nmol g–1) = A535×0.156
Cytokines determination: Using the ELISA technique (Dynatech Microplate Reader Model MR 5000), tumor necrosis alpha (TNF-α) and interleukin-1 beta (IL-1β) concentrations in brain areas were measured using rats' reagent ELISA kits purchased from Sino Gene Clon Biotech Co, Hang Zhou, China.
Statistical analysis: Comparisons between means were carried out using one-way Analysis of Variance (ANOVA) followed by post hock (Tukey) multiple comparisons test at p<0.05 according to the previous study24. This was carried out using Statistical Analysis System (SAS) program software; copyright (c) 1998 by SAS Institute Inc., Cary, NC, USA.
RESULTS
The yield, total phenolic content (TPC) and radical-scavenging activity (RSA) and reducing the power of Costus (Saussurea costus ) ethanolic extract (CEE) are shown in Fig. 1 and 2. Findings show that the yield has a moderate amount of total phenolic content that reveals high radical scavenging activity and reducing power. As shown in Fig. 3 total of 16 phenolic compounds were identified in CEE using HPLC analysis.
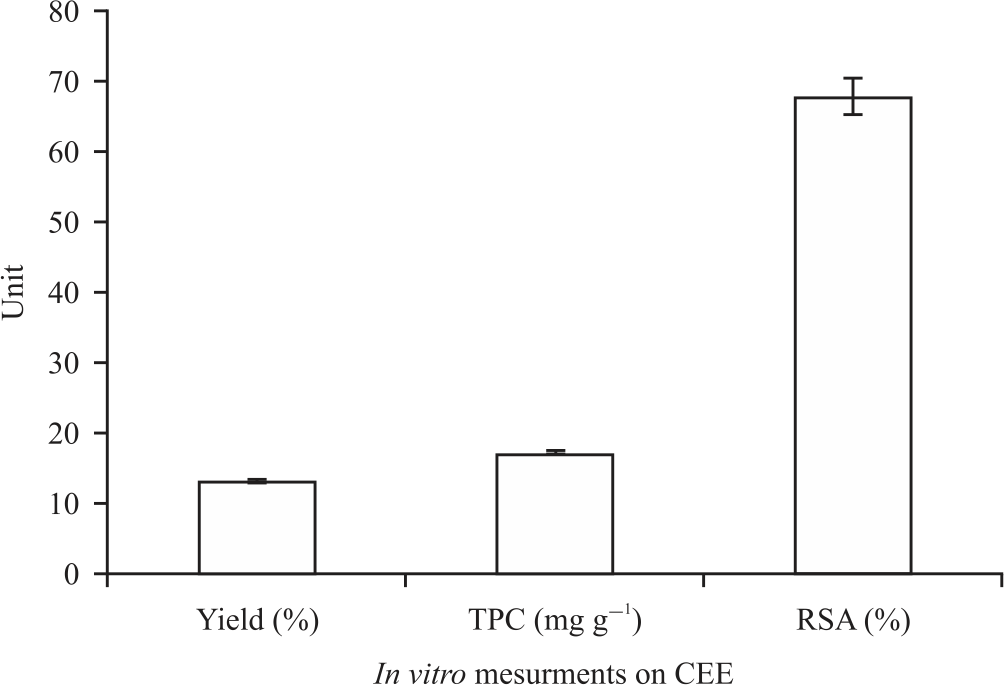 |
| Fig. 1: | Values of the yield, Total Phenolic Content (TPC) and Radical Scavenging Activity (RSA) of the ethanolic extract of dry the powdered roots of Saussurea costus; data presented as mean of triple replicates |
The compounds identified were found to include high contents of naringenin, chlorogenic acid, ferulic acid, taxifolin, gallic acid, caffeic acid (Fig. 3).
Comparing with the control group, the administration of bifenthrin (BF) resulted in a significant retardation in the rat's behavioral tests {rotarod performance (Fig. 4a) and spatial memory (Fig. 4b)}. Interestingly, the treatment of BF-intoxicated rats with CEE markedly improved the mentioned behavioral tests towards the normal values (Fig. 4a-b).
Unfortunately, the intoxication of rats with BF led to a marked drop in dopamine and serotonin levels and acetylcholinesterase (AChE-ase) activity in the three brain regions. Contrarily in a favorable manner, the administration of rats with CEE post-BF-intoxication significantly improved the two biogenic amines levels and AChE-ase activity in the concerned brain areas close to that of a normal group (Table 1).
Data in Table 2 showed the effect of different treatments on oxidant-antioxidant markers in the cortex, hippocampus and striatum tissue. In comparison to the control group, BF intake markedly increased the values of cortex, hippocampus and striatum MDA and NO and significantly lowered their values of GSH, SOD, CAT and GPx. However, the administration of CEE only had no unfavorable effect on these parameters. The BF-intoxicated animals that received CEE showed significant decreases in the cortex, hippocampus and striatum MDA and NO levels cabled with significant increases in the values of GSH, SOD, CAT and GPx as compared to the BF group.
In comparison to the control group, the obtained data recorded a significant rise in levels of TNF-α and IL1β in cortex, hippocampus and striatum of the BF-intoxicated group.
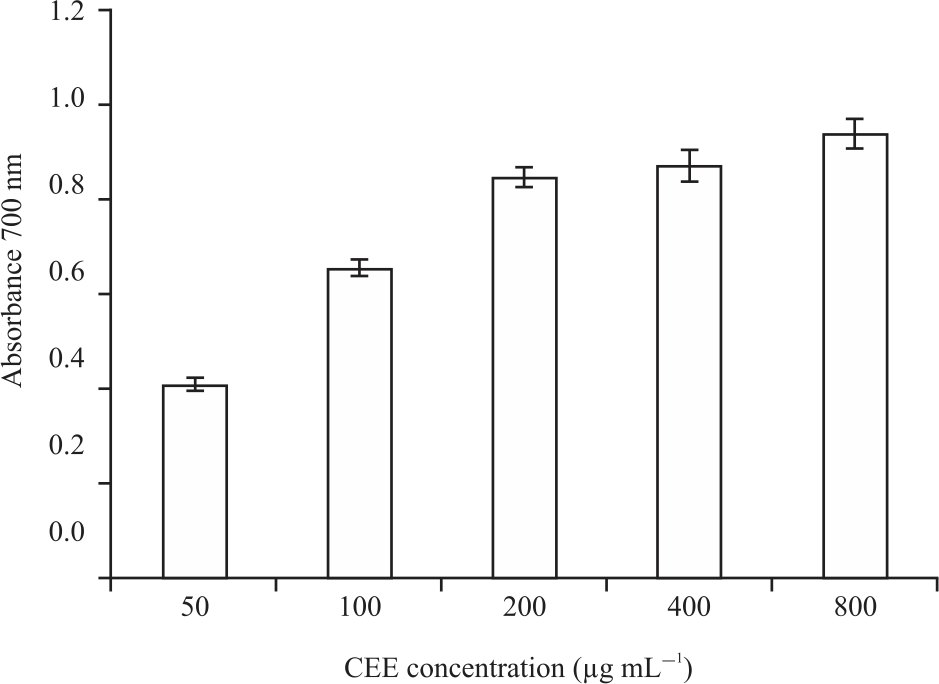 |
| Fig. 2: | Reducing power of the ethanolic extract of Saussurea costus data presented as mean of triple replicates |
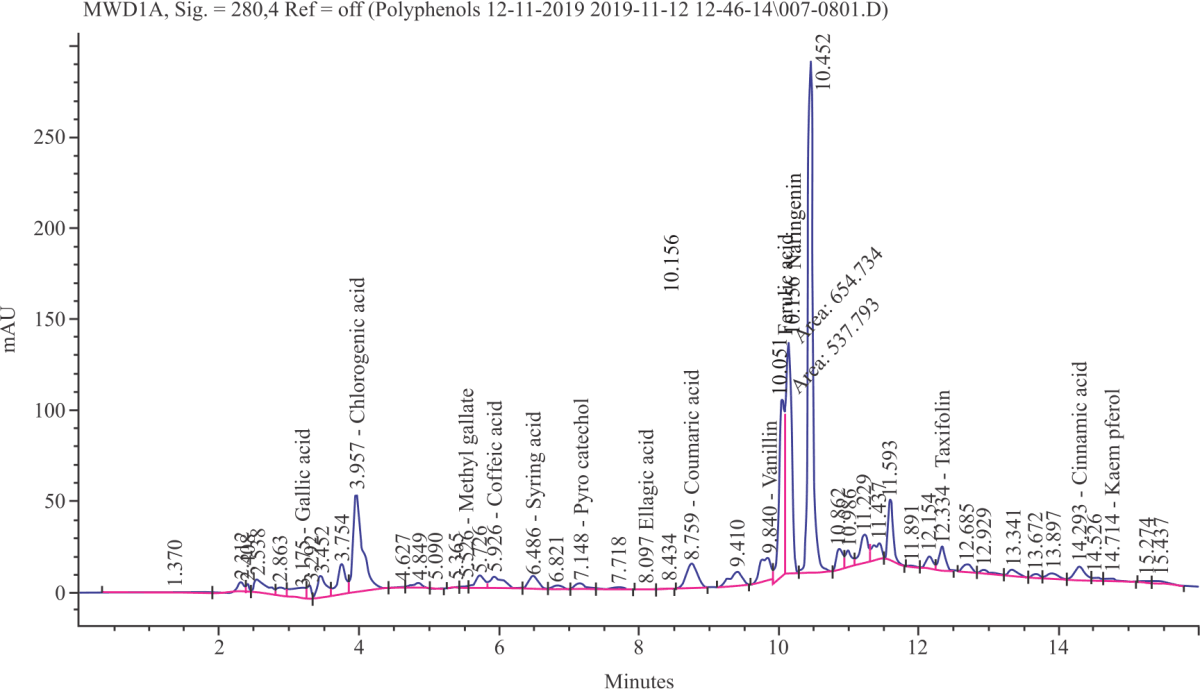 |
| Fig. 3: | HPLC imprint profile of ethanolic extract of Saussurea costus. 16 phenolic compounds were identified, other peaks not identified |
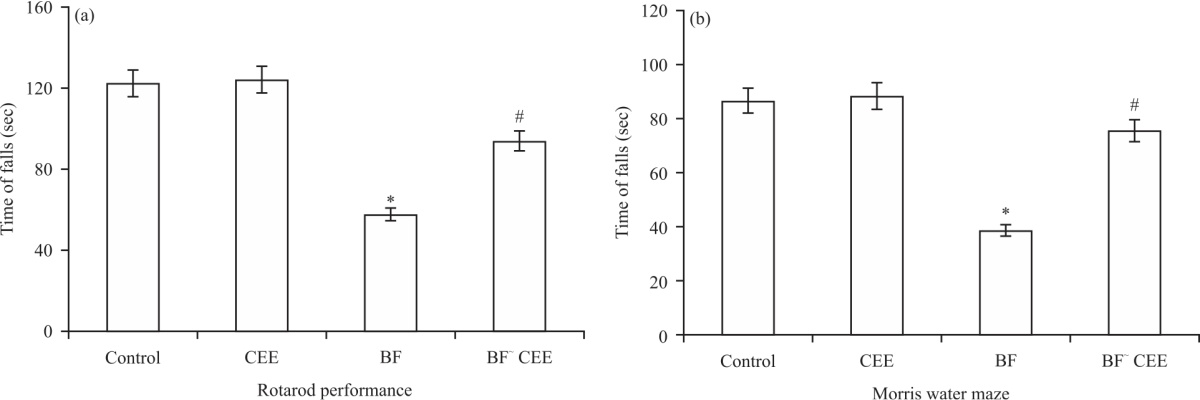 |
| Fig. 4: | Behavioral values of BF-intoxicated rats and/or CEE-treated rats |
| Table 1: Level of the biogenic amines and activity of AChE-ase in brain regions (cortex, hippocampus and striatum) of BF-intoxicated and/or CEE-treated rats | |||||||||
| Area | Parameters | Control | CEE | BF | BF then CEE | ||||
| Cortex | Dopamine (pg g–1) | 1222±74 | 1272±71 | 634±68* | 1157±57# | ||||
| Serotonin (pg g–1) | 1062±53 | 1110±55 | 684±53* | 1024±41# | |||||
| AChE-ase (μmoL min–1 g–1) | 9800±111 | 9680±121 | 7650±88* | 8912±62# | |||||
| Hippocampus | Dopamine (pg g–1) | 565±15 | 554±16 | 371±34* | 471±31# | ||||
| Serotonin (pg g–1) | 429±17 | 442±14 | 254±19* | 340±16# | |||||
| AChE-ase (μmoL min–1 g–1) | 11300±168 | 10985±122 | 8053±117* | 91802±98# | |||||
| Striatum | Dopamine (pg g–1) | 5505±99 | 5621±68 | 3522±133* | 3979±114# | ||||
| Serotonin (pg g–1) | 1098±66 | 1133±48 | 663±25* | 928±20# | |||||
| AChE-ase (μmoL min–1 g–1) | 9235±154 | 9307±111 | 8220±132* | 9370±96# | |||||
| Data are presented as mean ±SEM. Within the same row, * is significantly different from the control group, and # is significantly different from BF-group. BF: Bifenthrin; CEE: Costus ethanolic extract. | |||||||||
| Table 2: Oxidative stress status of cortex, hippocampus and striatum of control, BF-intoxicated and/or CEE- treated male albino rats | |||||||||||
| Area | Parameter | Control | CEE | BF | BF then CEE | ||||||
| Cortex | GSH (μmol g–1) | 5240±57 | 5344±61 | 4654±47* | 5057±34# | ||||||
| SOD (U g–1) | 3010±62 | 3122±58 | 1825±38* | 2465±25# | |||||||
| GPx (μmoL min–1 g–1) | 17.86±0.68 | 18.34±0.77 | 13.22±0.54* | 16.54±0.57# | |||||||
| CAT (μmoL min–1 g–1) | 6200±65 | 6184±54 | 4700±57* | 5884±42# | |||||||
| NO (nmoL g–1) | 78.1±1.2 | 82.3±1.1 | 97.2±2.4* | 86.4±1.4# | |||||||
| MDA (nmoL g–1) | 485±11.3 | 479±9.8 | 653±15.3* | 532±11.2# | |||||||
| Hippocampus | GSH (μmoL g–1) | 13240±74 | 13305±66 | 10231±61* | 12174±53# | ||||||
| SOD (U g–1) | 3464±71 | 3511±45 | 2357±49* | 3229±37# | |||||||
| GPx (μmoL min–1 g–1) | 23.56±0.74 | 24.06±0.64 | 18.87±0.83* | 22.26±0.78# | |||||||
| CAT (μmoL min–1 g–1) | 6895±75 | 6912±53 | 4955±64* | 5995±61# | |||||||
| NO (nmoL g–1) | 23±3.4 | 22.2±1.8 | 33.3±4.0* | 27.9±2.5# | |||||||
| MDA (nmoL g–1) | 463±8.7 | 442±7.8 | 725±17.4* | 551±12.3# | |||||||
| Striatum | GSH (μmoL g–1) | 11420±97 | 11574±75 | 9245±66* | 10261±61# | ||||||
| SOD (U g–1) | 2959±52 | 3115±61 | 2133±38* | 3028±2# | |||||||
| GPx (μmoL min–1 g–1) | 19.85±0.94 | 20.24±0.87 | 16.91±0.72* | 21.47±0.97# | |||||||
| CAT (μmoL min–1 g–1) | 6675±71 | 6722±66 | 4895±55* | 832±64# | |||||||
| NO (nmoL g–1) | 93±3.1 | 89±2.7 | 112.4±2.4* | 96±3.9# | |||||||
| MDA (nmoL g–1) | 556±9.2 | 549±8.7 | 735±8.6* | 632±6.8# | |||||||
| Data are presented as Mean±SEM, Within the same row, *Significantly different from the control group and #Significantly different from BF-group, BF: Bifenthrin, CEE: Costus ethanolic extract | |||||||||||
| Table 3: Inflammatory cytokines (TNFα and IL-1β) concentration in brain regions (cortex, hippocampus and striatum) of BF-intoxicated and/or CEE-treated rats | |||||
| Area | Parameter | Control | CEE | BF | BF then CEE |
| Cortex | TNF-α (ng g–1) | 13.81±0.311 | 12.91±0.392 | 16.54±0.72* | 14.27±0.58# |
| IL-1β (ng g–1) | 147.5±2.13 | 141.32±3.24 | 186.54±2.14* | 163.37±3.91# | |
| Hippocampus | TNF-α (ng g–1) | 26.87±0.214 | 25.71±0.12 | 41.32±0.44* | 31.55±0.325# |
| IL-1β (ng g–1) | 167.54±4.41 | 159.87±21 | 336.66±3.77* | 211.54±3.17# | |
| Striatum | TNF-α (ng g–1) | 17.78±0.17 | 17.55±0.34 | 18.16±0.32* | 18.64±0.12# |
| IL-1β (ng g–1) | 124.54±3.78 | 118.58±2.54 | 287.65±4.66* | 168.18±71# | |
| Data are presented as Mean±SEM, Within the same row, *Significantly different from the control group and #Significantly different from BF-group, BF: Bifenthrin; CEE: Costus ethanolic extract | |||||
Favorably, CEE post-treatment of BF-intoxicated rats markedly restored the level of inflammatory cytokines closer to those of normal group; it significantly reduced TNF-α and IL1β in all the brain regions compared to the corresponding values of BF-intoxicated animals (Table 3).
DISCUSSION
Our results showed that bifenthrin caused significant decreases in the concentration of dopamine and serotonin in the frontal cortex, hippocampus and corpus striatum of rats. Bifenthrin is rapidly metabolized in the liver by carboxylesterasesresulting in the generation of Reactive Oxygen Species (ROS)25. These findings are in agreement with that previously established by previous studies16,26.
The present data showed also depletion in rotarod performance and spatial memory in bifenthrin intoxicated rats. Besides, previous study27 stated that the decrease in dopamine levels in the hippocampus, frontal, lateral and medial temporal cortex is accompanied by a reduction in cognitive and motor performance. The alterations in memory functionsare also associated with a decline in hippocampal neurogenesis28,29. As LTP and dendritic spines are important in learning and memory, their loss is accompanied by impaired brain plasticity which affects cognitive functions30. On the other hand, the reduction in serotonin levels is associated with various physiological operations such as circadian rhythm, sleeping, eating and cognition31.
In the current study, the administration of bifenthrin to rats inhibited AChE activity in the hippocampus, cortex and striatum. AChE change Ach into choline plus acetate in the synaptic cleft32,33. Bifenthrin was documented to has an anticholinesterase effect16 which occurs by reduction of AChE activity at synapses in the brain leading to the accumulation of acetylcholine and more activation of its receptor at the neuromuscular junction and in both autonomic and central nervous system. Moreover, researches in the past have declared that cognitive and emotional defects are managed by AChE inhibition34,35.
The increased MDA and Nitric Oxide (NO) levels in the hippocampus, cortex and striatum of bifenthrin-treated rats, recorded in the present study, agree with previously documented results, demonstrating that bifenthrin causes its neuronal injury via oxidative stress16. At the same time, the antioxidant defense system was negatively affected by bifenthrin intoxication, where the activity of SOD, GPx and CAT and the GSH content in the three brain areas, were found to be significantly decreased by bifenthrin intoxication.
Our results parallel with previous study16 who reported that the drop in the capacity of these enzymes strengths the radicals-production of both superoxide and hydrogen peroxide in the brain; this in its turn results in peroxidation of the membrane lipids leading to progressive peroxidation and NO production; the later comes from the activated polymorphic nuclear leukocytes in consequence to inflammation36. The liberation of NO by the inducible nitric oxide synthase (iNOS) represents the fundamental spark in the cytokine-mediated cell destruction37.
In respect to this study, an elevation in the concentration of the proinflammatory cytokines (TNFα and IL-1β) was noticed in the hippocampus, cortex and striatum of the bifenthrin-treated rats; this unfavorable rise could be attributed to the stimulation of inflammatory responses of the acute phase, directing it to release that cytokine, those counted in the production of ROS38,39. This ROS production accelerates the development of neurodegenerative diseases40. Our results are similar to those observed in other previous studies41,42 who demonstrated significantly increased NF-kB and pro-inflammatory cytokines.
Co-administration of bifenthrin-treated rats with Costus (Saussurea costus) ethanolic extract CEE was greatly effective in minimizing lipid peroxidation and improving the antioxidant status in the frontal cortex, hippocampus and corpus striatum; this effect evidenced by decreasing the elevated levels of MDA and NO towards the corresponding values of normal rats and significant improvement of the antioxidant defense system. Additionally, CEE played neuro-protective potential that could be achieved from the sharp increase in dopamine, serotonin and AChE-ase values that decreased by bifenthrin intake.
The mechanism by which CEE offered its protective effect against bifenthrin neurotoxicity, based on its antioxidant ability that proved from the data of DPPH radical scavenging activity and reducing power ability and further confirmed by the increased values of GSH, SOD, CAT and GPx in the brain areas. This, possibly by the ability of CEE to prevent GSH exhaustion and increase the endogenous defensive capacity of the brain to combat the free radicals and inflammation induced by bifenthrin; this leads to improvement of neuron integrity and prevents its destruction and consequently enhances brain function and hence an improvement of behavioral activity.
The antioxidant activity of CEE may reside mainly in its phenolic contents. These compounds possibly prevent free radicals from reaching and attacking the large molecules like DNA, lipids and proteins evidencing predictive participation of an extract as a chain blocking antioxidant opposite to lipid peroxidation; consequently, restores the membrane permeability and repair cellular integrity of the brain tissues. As shown in our results, 16 phenolic compounds were identified in the CEE using HPLC analysis. The compounds identified were found to include high contents of naringenin, chlorogenic acid, ferulic acid, taxifolin, gallic acid, caffeic acid. Previous reports have indicated that CEE contains other antioxidant and bioactive phytochemicals as described by previous stdueis43,44.
From the results of the present study, we suggest to minimize the use of this pesticide because of its neurotoxic effect and instead, recommend the use of the ethanolic extract of the Saussurea costus due to its radical scavenging activity and high phenolic content revealed from the HPLC analysis, more studies are needed to determine the most effective constituent responsible for its anti-neurobehavioral effect. CEE is also recommended for use by workers using this pesticide.
CONCLUSION
The present study concluded that Costus (Saussurea costus) ethanolic extract attenuates the brain dysfunction induced by bifenthrin as indicated by improving behavioral performance through normalize lipid peroxidation mechanism and its related biochemical changes. Costus (Saussurea costus) ethanolic extract may be promising as a neurobehavioral protective agent against pesticide neurotoxicity through its antioxidant and radical scavenging activities.
SIGNIFICANCE STATEMENT
This study discovered the effect of ethanolic extract of Saussurea costus on neurobehavioral toxicity can be beneficial for protection against bifenthrin neurotoxicity in male rats. This study will help the researchers to uncover the critical areas of using an extract of Saussurea costus that many researchers were not able to explore.
ACKNOWLEDGMENT
I would like to thanks the colleagues at Medical Physiology Laboratory and workers at an animal house at the National Research Centre, Dokki, Cairo, Egypt.
REFERENCES
- Rehman, H., M. Ali, F. Atif, M. Kaur, K. Bhatia and S. Raisuddin, 2006. The modulatory effect of deltamethrin on antioxidants in mice. Clin. Chim. Acta, 369: 61-65.
CrossRef - Costa L.G., G. Giordano, M.Guizzetti and A. Vitalone, 2005. Neurotoxicity of pesticides: A brief review. Front. Biosci., 13: 1240-1249.
CrossRefPubMedDirect Link - Syed, F., K.K. Awasthi, L.P. Chandravanshi, R. Verma and N.K. Rajawat et al 2017. Bifenthrin-induced neurotoxicity in rats: involvement of oxidative stress. Toxicol. Res., 7: 48-58.
CrossRefDirect Link - Domagalski, J.L., D.P. Weston, M. Zhang and M. Hladik, 2010. Pyrethroid insecticide concentrations and toxicity in streambed sediments and loads in surface waters of the San Joaquin Valley, California, USA. Environ. Toxicol. Chem., 29: 813-823.
CrossRefDirect Link - Soderlund, D.M., J.M. Clark, L.P. Sheets, L.S. Mullin and V.J. Piccirillo et al., 2002. Mechanisms of pyrethroid neurotoxicity: Implications for cumulative risk assessment. Toxicology, 171: 3-59.
CrossRef - Prabhu, M. Natesh, Pai, G. Preethi, S.V. Kumar and Gaurav, 2014. Diuretic activity of aqueous ethanolic extract of leaves of Costus speciosus in normal wistar albino rats. Res. J. Pharm. Biol. Chem. Sci., 5: 127-132.
Direct Link - Girgis S.M., T.M.T. Shoman, S.M. Kassem, A.E. El-Din and K.B. Abdel-Aziz, 2015. Potential protective effect of costus speciosus or its nanoparticles on streptozotocin-induced genotoxicity and histopathological alterations in rats. J. Nutr. Food Sci., 3: 2-7.
CrossRefDirect Link - Kuniyal, C.P., Y.S. Rawat, S.S. Oinam, J.C. Kuniyal and S.C.R. Vishvakarma, 2005. Kuth (Saussurea lappa) cultivation in the cold desert environment of the Lahaul Valley, Northwestern Himalaya, India: Arising threats and need to revive socio-economic values. Biodivers. Conserv., 14: 1035-1045.
CrossRefDirect Link - Farombi, E.O. and A. Fakoya, 2005. Free radical scavenging and antigenotoxic activities of natural phenolic compounds in dried flowers of Hibiscus sabdariffa. Mol. Nutr. Food Res., 49: 1120-1128.
CrossRefDirect Link - Okeke, I.N., A. Lamikanra and R. Edelman, 1999. Socioeconomic and behavioral factors leading to acquired bacterial resistance to antibiotics in developing countries. Emerg. Infect. Dis., 5: 18-27.
CrossRefPubMedDirect Link - Filipiak-Szok, A., M. Kurzawa, E. Szłyk, M. Twarużek, A. Błajet-Kosicka and J. Grajewski, 2017. Determination of mycotoxins, alkaloids, phytochemicals, antioxidants and cytotoxicity in Asiatic ginseng (Ashwagandha, Dong quai, Panax ginseng). Chem. Pap., 71: 1073-1082.
CrossRefDirect Link - Jayaprakasha, G.K., T. Selvi and K.K. Sakariah, 2003. Antibacterial and antioxidant activities of grape (Vitis vinifera) seed extracts. Food Res. Int., 36: 117-122.
CrossRefDirect Link - Nogala-Kalucka, K., J. Korczak, M. Dratwia, E. Lampart-Szczapa, A. Siger and M. Buchowski, 2005. Changes in antioxidant activity and free radical scavenging potential of rosemary extract and tocopherols in isolated rapeseed oil triacylglycerols during accelerated tests. Food Chem., 93: 227-235.
CrossRefDirect Link - Sethiya, N.K., A. Trivedi and S. Mishra, 2014. The total antioxidant content and radical scavenging investigation on 17 phytochemical from dietary plant sources used globally as functional food. Biomed. Prev. Nutr., 4: 439-444.
CrossRefDirect Link - Syed, F., K.K. Awasthi, L.P. Chandravanshi, R. Verma and N.K. Rajawat et al 2017. Bifenthrin-induced neurotoxicity in rats: involvement of oxidative stress. Toxicol. Res., 7: 48-58.
CrossRefDirect Link - Xu, C., B. Yang, W. Zhu, X. Li, J. Tian and L. Zhang, 2015. Characterisation of polyphenol constituents of Linderae aggregate leaves using HPLC fingerprint analysis and their antioxidant activities. Food Chem., 186: 83-89.
CrossRefDirect Link - Zatroch, K.K., C.G. Knight, J.N. Reimer and D.S.J. Pang, 2017. Refinement of intraperitoneal injection of sodium pentobarbital for euthanasia in laboratory rats (Rattus norvegicus). BMC Vet. Res., 13: 60-67.
CrossRefDirect Link - Syed, F., L.P. Chandravanshi, V.K. Khanna and I. Soni, 2016. Beta-cyfluthrin induced neurobehavioral impairments in adult rats. Chem. Biol. Interact., 243: 19-28.
CrossRefDirect Link - Chen, Q., Y. Niu, R. Zhang, H. Guo, Y. Gao, Y. Li and R. Liu, 2010. The toxic influence of paraquat on hippocampus of mice: Involvement of oxidative stress. NeuroToxicology, 31: 310-316.
CrossRefDirect Link - Chen, Q., Y. Niu, R. Zhang, H. Guo, Y. Gao, Y. Li and R. Liu, 2010. The toxic influence of paraquat on hippocampus of mice: Involvement of oxidative stress. NeuroToxicology, 31: 310-316.
CrossRefDirect Link - Kim, C., M.B. Speisky and S.N. Kharouba, 1987. Rapid and sensitive method for measuring norepinephrine, dopamine, 5-hydroxytryptamine and their major metabolites in rat brain by high-performance liquid chromatography. J. Chromatogr. A, 386: 25-35.
CrossRefDirect Link - Ruiz-Larrea, M.B., A.M. Leal, M. Liza, M. Lacort and H. de Groot, 1994. Antioxidant effects of estradiol and 2-hydroxyestradiol on iron-induced lipid peroxidation of rat liver microsomes. Steriods, 59: 383-388.
CrossRefDirect Link - Steel, R.G.D. and J.H. Torrie, 1980. Principles and Procedures of Statistics: A Biometrical Approach. 2nd Edn., McGraw Hill Book Co., New York, USA., ISBN-13: 9780070609266, Pages: 633.
Direct Link - Ross, M.K., A. Borazjani, C.C. Edwards and P.M. Potter, 2006. Hydrolytic metabolism of pyrethroids by human and other mammalian carboxylesterases. Biochem. Pharmacol., 71: 657-669.
CrossRefDirect Link - El-Fattah B.M. El-Beltagy, A., K.A.M. Elbakry, M.M.A. Elghazaly, L.S. Ali and N.H.M.E.L. Daqaqq, 2019. Adverse effects of deltamethrin on the cerebellum of mothers rats and their offspring and the possible ameliorative role of melatonin. Int. J. Pure Appl. Zool., 7: 61-80.
CrossRefDirect Link - Nikhra, V., 2017. The aging brain: recent research and concepts. Gerontol. Geriatrics Stud.,.
CrossRefDirect Link - Burke, S.N. and C.A. Barnes, 2006. Neural plasticity in the ageing brain. Nat. Rev. Neurosci., 7: 30-40.
CrossRefDirect Link - Koleske, A.J., 2013. Molecular mechanisms of dendrite stability. Nat. Rev. Neurosci., 14: 536-550.
CrossRefDirect Link - Dickstein, D.L., C.M. Weaver, J.I. Luebke and P.R. Hof, 2013. Dendritic spine changes associated with normal aging. Neuroscience, 251: 21-32.
CrossRefDirect Link - Mattson, M.P., 2015. Lifelong brain health is a lifelong challenge: From evolutionary principles to empirical evidence. Ageing Res. Rev., 20: 37-45.
CrossRefDirect Link - Deiana, S., B. Platt and G. Riedel, 2011. The cholinergic system and spatial learning. Behavioural Brain Res., 221: 389-411.
CrossRefDirect Link - Gonçalves, J.F., F.T. Nicoloso, P. Da Costa, J.G. Farias and F.B. Carvalho et al., 2012. Behavior and brain enzymatic changes after long-term intoxication with cadmium salt or contaminated potatoes. Food Chem. Toxicol., 50: 3709-3718.
CrossRefDirect Link - Cardona, D., G. López-Crespo, M.C. Sánchez-Amate, P. Flores and F. Sánchez-Santed, 2011. Impulsivity as long-term sequelae after chlorpyrifos intoxication: time course and individual differences. Neurotox. Res., 19: 128-137.
CrossRefDirect Link - Cardona, D., C. López-Granero, F. Cañadas, J. Llorens, P. Flores, F. Pancetti and F. Sánchez-Santed, 2013. Dose-dependent regional brain acetylcholinesterase and acylpeptide hydrolase inhibition without cell death after chlorpyrifos administration. J. Toxicol. Sci., 38: 193-203.
CrossRefDirect Link - Zheng, S., M. Ma, Z. Li, Y. Hao, H. Li, P. Fu and S. Jin, 1992. Posttreatment of Maresin1 Inhibits NLRP3 inflammasome activation via promotion of NLRP3 ubiquitination. FASEB J., 6: 3051-3064.
CrossRefDirect Link - Azevedo-Martins, A.K., S. Lortz, S. Lenzen, R. Curi, D.L. Eizirik and M. Tiedge, 2003. Improvement of the mitochondrial antioxidant defense status prevents cytokine-induced nuclear factor- B activation in insulin-producing cells. Diabetes, 52: 93-101.
CrossRefDirect Link - Heydarpour, P., R. Rahimian, G. Fakhfouri, S. Khoshkish and N. Fakhraei et al., 2016. Behavioral despair associated with a mouse model of Crohn's disease: Role of nitric oxide pathway. Progr. Neuro-Psychopharmacol. Biol. Psychiatry, 64: 131-141.
CrossRefDirect Link - Duramad, P., I.B. Tager and N.T. Holland, 2007. Cytokines and other immunological biomarkers in childrens environmental health studies. Toxicol. Lett., 172: 48-59.
CrossRefDirect Link - Fiebich, B.L., S. Akter and R.S. Akundi, 2014. The two-hit hypothesis for neuroinflammation: role of exogenous ATP in modulating inflammation in the brain. Front. Cell. Neurosci., Vol. 8.
CrossRefDirect Link - Zhao, M., F.S. L. Wang, X. Hu, F. Chen and H.M. Chan, 2017. Acrylamide-induced neurotoxicity in primary astrocytes and microglia: Roles of the Nrf2-ARE and NF-κB pathways. Food Chem. Toxicol., 106: 25-35.
CrossRefDirect Link - Gargouri, B., N.M. Yousif, M. Bouchard, H. Fetoui and B.L. Fiebich, 2018. Inflammatory and cytotoxic effects of bifenthrin in primary microglia and organotypic hippocampal slice cultures. J. Neuroinflammation.
CrossRefDirect Link - Kotebagilu, N.P., V.R. Palvai and A. Urooj, 2014. Protective effect of selected medicinal plants against hydrogen peroxide induced oxidative damage on biological substrates. Int. J. Med. Chem., Vol. 2014.
CrossRefDirect Link - Singh, A.K., P.N. Saxena and H.N. Sharma, 2009. Stress induced by beta-cyfluthrin, a type-2 pyrethroid on brain biochemistry of Albino rat (Rattus norvegicus). Biol. Med., 1: 74-86.
CrossRefDirect Link