
Afriza Yelnetty
Department of Animal Production, Faculty of Animal Husbandry, Sam Ratulangi University, Manado, North Sulawesi, Indonesia
Purwadi
Department of Animal Food Technology, Faculty of Animal Husbandry, Brawijaya University, Malang, East Java, Indonesia
Trina Ekawati Tallei
Department of Biology, Faculty of Mathematics and Natural Sciences, Sam Ratulangi University, Manado, North Sulawesi, Indonesia
LiveDNA: 62.16548
Pakistan Journal of Biological Sciences
Year: 2020 | Volume: 23 | Issue: 7 | Page No.: 883-890







ABSTRACT
Background and Objective: Lactic Acid Bacteria (LAB) isolated from goat milk have been known to have various medicinal properties, therefore they are considered as a source of potential probiotics. This research was aimed at evaluating and identifying the LAB isolated from spontaneously fermented goat milk as potential probiotics. Materials and Methods: The fresh goat milk fermented for 4 days was serially diluted, plated on de Man Rogosa and Sharpe (MRS) agar supplemented with 1% CaCO3 as selective medium, then purified accordingly. The isolated LAB were screened for their potential to inhibit enteric pathogen bacteria using well diffusion method. Their capabilities to withstand the bile salt and acid environment were also evaluated. The production of organic acids was also assessed. The potential probiotics were identified molecularly using 16S rRNA. Results: The study confirmed that LAB isolated from spontaneous fermentation of goat milk was Lactobacillus plantarum based on 16S rRNA gene marker. This bacterium showed antimicrobial activity against indicator bacteria, ability to live after exposure in bile salt solution and resistance to low acidic environment. The organic acids produced by this bacterium were lactic, acetic, propionic and butyric acids. Conclusion: This study concluded that Lactobacillus plantarum YN.1.3 can be further investigated as potential probiotic as it showed antimicrobial activity, withstood the acidic environment and bile salt solution, as well as produced organic acids.
PDF Abstract XML References Citation
Copyright: © 2020. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Afriza Yelnetty, Purwadi and Trina Ekawati Tallei, 2020. Indigenous Lactic Acid Bacteria Isolated from Spontaneously Fermented Goat Milk as Potential Probiotics. Pakistan Journal of Biological Sciences, 23: 883-890.
DOI: 10.3923/pjbs.2020.883.890
URL: https://scialert.net/abstract/?doi=pjbs.2020.883.890
DOI: 10.3923/pjbs.2020.883.890
URL: https://scialert.net/abstract/?doi=pjbs.2020.883.890
INTRODUCTION
Lactic Acid Bacteria (LAB) probiotic are mainly used by dairy processors in their fermented products. The probiotic characteristics of LAB, beside having the beneficial effect on health, they should give added value for fermented product such as extending the shelf life of end products. These bacteria should also inhibit the growth of spoilage and pathogenic bacteria in food1-6. In general, LAB are dominantly available and the digestive tract both humans and animals7,8 and able to produce antimicrobial compounds such as acetic acid, lactic acid, propionic acid, butyric acid, benzoic acid, hydrogen peroxide and bacteriocin which could inhibit or even kill the spoilage and pathogenic bacteria6,9-11. Bacteriocins are peptide that are ribosomally synthesized and have a bactericidal or bacteriostatic effect on closely related bacteria12.
LAB such as Lactobacillus plantarum and L. fermentum have been reported to be the most commonly LAB species associated with spontaneous lactic acid fermentation13. Soomro et al.14 reported that LAB which produce antimicrobial substances have a considerable advantage in competition with other microorganisms like pathogenic and harmful bacteria. Lactobacillus plantarum was shown to have a greater level of inhibitory activity against Bacillus cereus and Staphylococcus aureus compared to L. acidophilus. The inhibitory activity of L. plantarum was caused by the organic acids they produce i.e., acetic acid, lactic acid, propionic acid, butyric acid, benzoic acid and phenyl lactic beside bacteriocin15. Because probiotic LAB have the ability to live and colonize the digestive tract even though only in small proportions16, hence they should have the ability to withstand bile salt solution and acidic environment15. The aims of this study were to evaluate and identify the potential of LAB isolates obtained from spontaneously fermented goat for their potential as probiotics.
MATERIALS AND METHODS
Sample preparation and fermentation: The study was conducted at Microbiology Laboratory Sam Ratulangi University and Gadjah Manado University, Indonesia from March-July, 2019. The freshly milked goat milk was fermented for four days in room temperature until there was a decrease in pH to 4. This fermented goat milk was used as source of LAB. One milliliter of fermented goat milk was added with 9 mL of peptone water and diluted serially up to 10–9. The isolation method was conducted following the procedure suggested by Tallei et al.17 with a slight modification. One milliliter of sample was taken from each solution and plated on de Man, Rogosa and Sharpe (MRS) agar supplemented with 1% CaCO3 and incubated at 37°C for 48 h. Each colony that grew and formed a clear zone was picked, grown on MRS agar slant and incubated at 37°C for 48 h. The growing colonies were then picked and re-planted in a Petri dish containing MRS agar and CaCO3 with streak method to obtain pure colonies. The method was repeated three times until it was assured that each of the growing colonies was originated from one cell.
Analysis of organic acids produced by lactic acid bacteria: The selected LAB were assessed for their potential to produce organic acid by fermenting them for 5 days in MRS broth. The organic acids analyzed were lactic, acetic, propionic and butyric acids following the method from Bevilacqua and Califano18. The analysis of these compounds was conducted using High Performance Liquid Chromatography (HPLC). Ten milliliters of each sample were added with 5 mL acetonitrile, homogenized then centrifuged at 6000 rpm for 5 min. The supernatant was filtered using 0.45 μm filter paper and repeated twice. The aqua bidest was used for elution with the flow rate of 1.5 mL min–1. The UV-Vis Shimadzu SPD-6AV with a wavelength of 214 nm was used as a detector. The pH of each sample was measured using pH meter (Orion model 420 A). As much as 20 μL of each sample was injected into the HPLC.
Determination of antimicrobial activity: Well diffusion method was used to identify the nature of inhibitory substances following the method of Rammelsberg and Radler19. The inhibitory effect of LAB isolates on pathogenic and spoilage bacteria were studied. Three negative pathogenic bacteria (Esherichia coli, Listeria monocytogenes and Salmonella typhimurium) and two positive pathogenic bacteria (Staphylococcus aureus and Bacillus cereus) were used as indicator microorganisms. The indicator bacteria were maintained on Trypticase Soy Agar (TSA) and moved to Nutrient Agar (NA) for evaluation. The procedure was originated from Tallei et al.17 with a modification. Briefly the LAB were grown in MRS broth for 24 h at 37°C in 1 mL Eppendorf tubes. The LAB were heat-killed for 2 h at 60°C and centrifuged at 10,000 for 10 min to prepared Cell Free Culture Supernatants (CFSs). The pH of other half of CFSs was adjusted (neutralized) to 6.0 with 6 mol L–1 NaOH to eliminate the action of lactic acid on the indicator bacteria. The heat-killed non-neutralized and neutralized CFSs were used as sources of antimicrobial substances. The indicator bacteria were poured onto NA agar media which already had 5 mm deep-wells. Approximately 100 μL of each CFSs were poured into the wells. The plates were incubated at 37°C for 48 h. The resulting zones of inhibition were measured.
Determination of bile salt tolerance: Determination of bile salt tolerance was carried out using the method as described by Vinderola and Reinheimer20 with a slight modification. The Cell (108 CFU mL–1) were grown in MRS broth for 24 h. The tolerance against bile salt was determined using MRS broth containing 0.5% bile salt (Oxoid) which was inoculated with active culture of the LABs and incubated at 37°C for 4 h. The viable LABs were enumerated after 4 h of incubation using pour plate technique on MRS agar. The probiotic Lactobacillus acidophillus was used as control.
Resistance to low pH: Determination of resistance to low pH (3.0) was carried out using the method as described by Tallei et al.17 with modification. Active cultures of LAB were inoculated in MRS broth for 24 h. The media were adjusted to 3 with 1 N HCl and the cultures were incubated at 37°C for 4 h. Viable microorganisms were enumerated using pour plate techniques on MRS Agar. The probiotic Lactobacillus acidophillus was used as control.
Identification of potential probiotics: Identification was performed to selected LAB isolates which showed the best results on antibacterial activity, tolerance to bile salt and acid resistance. Each isolate was confirmed using Gram-staining, API 50 CH stripes system (bioMe ́rieux, Lyon, France) and molecular identification using 16S rRNA gene. Carbohydrate fermentation pattern was determined by API 50 CH stripes system according to the instruction manual. The data collected using API 50 CH test kit were confirmed using probabilistic identification at API Web Database V51. The molecular identification was carried out according to the procedure provided by Fatimawali et al.21. The identification was conducted using Ez-Taxon database.
RESULTS AND DISCUSSION
Isolation of lactic acid bacteria: A total of 3 pure colonies, designated as YN1.1, YN1.3 and YN1.6 were able to be isolated and purified from MRS agar containing CaCO3 based on their distinct morphological characters. The presence of clear zones (Fig. 1a) indicated that these bacteria metabolized sugar to produce acid, such as lactic acid. The clear zone was a result of reaction between lactic acid and CaCO3 in the media to produce soluble calcium lactate. Each colony was purified several times to obtain pure isolates (Fig. 1b-d).
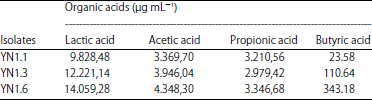
Production of organic acids: At the end of fermentation, an analysis of organic acids produced by each of LAB isolates was carried out. Organic acids are the result of LAB metabolism during fermentation. The analysis was out using HPLC included the determination of the amount of lactic, acetic, propionic and butyric acids in fermentation media. The result of the analysis (Table 1) showed that lactic acid was the most organic acid produced by all isolates compared to other organic acids. Isolate YN1.6 produced the highest amount of organic acids. Organic acids produced by probiotics act as antimicrobial substances and have a direct activity against pathogens22. The main LAB in dairy products fermentation are lactobacilli which produce lactic, acetic, formic, caproic, propionic, butyric and valeric acids23.
| Table 1: | Organic acids produced by LAB isolates |
 | |
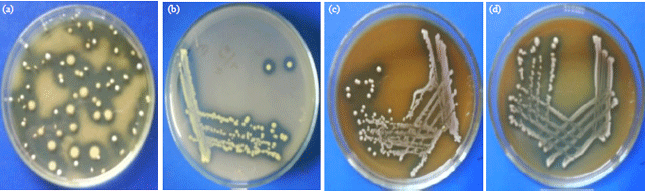 | |
| Fig. 1(a-d): | (a) LAB colonies that produced clear zones on MRS agar supplemented with 1% CaCO3, (b) Purified isolate YN1.1, (c) Purified isolate YN1.3 and (d) Purified isolate YN1.6 |
| Table 2: | Antibacterial activity produced by heat-killed non-neutralized CFSs of isolates YN1.1, YN |
 | |
*L. acidophillus as control, **ND: Not detected | |
| Table 3: | Inhibitory ability of neutralized CFSs of all isolates against spoilage and pathogenic bacteria |
 | |
| *L. acidophillus as control, **ND: Not detected | |
Antibacterial activity of heat-killed non-neutralized Cell Free Supernatants (CFSs): Antibacterial activity experiments against indicator bacteria were performed in triplicate and all plates were incubated at 37°C for 48 h. Table 2 shows the mean diameter of inhibition zones of heat-killed non-neutralized CFSs. Almost all isolates (YN1.1, YN1.3 and YN1.6) showed antibacterial activities against the indicator bacteria. However L. acidophillus (FNCC) and isolate YN1.1 did not show antibacterial activity against L. monocytogenes. According to Mirzaei et al.24, the inhibition zones were classified as follows: <11 mm (negative-), 11-16 mm (mild +), 17-22 mm (strong ++) and >23 mm (very strong +++). All isolates including control were very effective in inhibiting the growth of S. aureus. The biggest inhibition zone was shown by isolate YN1.3 by 22.70±0.50 mm (strong to very strong inhibitory activity). Mild inhibitory activity was shown by all isolates on S. typhimurium and B. cereus. All isolates showed negative inhibitory activity against E. coli and L. monocytogenes.
Ogunbanwo et al.25 noted that L. plantarum F1 inhibited Gram-positive bacteria B. acillus cereus ATCC 9634 (10 mm), S. aureus ATCC 14458 (8 mm) and Gram-negative bacterium S. tiphymurrium (6 mm), but did not inhibit L. monocytogenes CHRL 587. Further more, Liasi et al.26 recorded that L. plantarum LA 22 isolated from Budu fish product had inhibitory effect on pathogen bacteria Salmonella entertica and S. aureus (15-18 mm, respectively), E. coli (10-14 mm), but less effective on Listeria monocytogenes (6-9 mm). Tambekar et al.27 also noted that L. plantarum G.7 isolated from goat milk inhibited S. tiphymurium (25 mm) and E. coli (19 mm). Buntin et al.28 reported that Lactobacillus pentaceus had the ability to inhibit the growth of S. aureus, Salmonella sp. and E. coli.
Although the capability of antimicrobial activity against indicator bacteria in this study was shown by all isolates, however the inhibition effectiveness was different from one isolate to another. This might be due to different amount of organic acids produced by these microorganisms. According to Noordiana et al.29, LAB had a bigger ability in inhibiting many microorganisms, including pathogenic bacteria and the spoilage microorganisms due to organic acid, hydrogen peroxide, diacetyl, inhibitory enzyme and bacteriocin. Ray and Sandine30 reported that the ability of lactic acid to inhibit pathogenic bacteria was largely assigned to its ability to penetrate the cytoplasmic membrane of the spoilage and pathogenic bacteria, so to reduce intracellular pH which in turn disrupt the transmembrane proton motive force. Servin31 recorded that antibacterial activities of LAB involved numerous mechanisms of action among other producing H2O2, metabolites lactic acid and antimicrobial substances, such as bacteriocin and non bacteriocin molecules. Based on Gram characteristic of indicator bacteria, all isolates in this study was much more effective against Gram-positive compare to Gram-negative bacteria. Hernandez et al.32 reported that L. plantarum TF 711 had the highest inhibitory activity on Gram-positive bacteria such as B. cereus, Clostridium sporogenes and S. aureus and also had mild inhibitory effect on Gram-negative bacteria such as Shigella sonnei and Klebsiella pneumoniae.
Antimicrobial activity of neutralized Cell Free Supernatants: Neuralized cell-free supernatant of each isolate showed a different inhibition zones compared to heat-killed non-neutralized CFSs. The control LAB Lactobacillus plantarum only inhibited S. thiphymurium and S. aureus. The inhibition activities of neutralized CFSs are presented in Table 3.
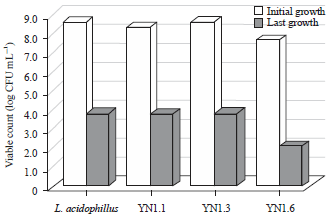 | |
| Fig. 2: | Number of viable counts of LAB after exposure for 4 h in 0.5% bile salt solution |
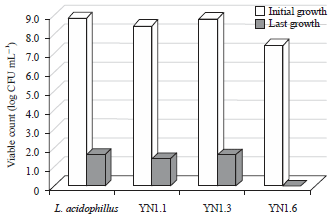 | |
| Fig. 3: | Number of viable counts of LAB after exposure for 4 h in media with pH 3 |
Inhibitory ability of each isolate was different from each other where CFS of L. acidophillus and YN1.6 had mild inhibition on S. aureus, while YN1.1 and YN1.3 had strong inhibition on S. aureus. However, all of the isolates and control bacteria showed negative inhibitory ability against all of the rest of indicator bacteria. The inhibitory activity shown by neutralized CFSs of all isolates and control bacteria indicated that the LAB not only produced organic acid, but also other antimicrobial component such as bacteriocin. Bacteriocin of LAB produced by L. plantarum is known as plantaricin. The similar result was reported by Francois et al.6 where they observed that CFS of L. plantarum inhibited several types of food spoilage bacteria and also several types of pathogenic bacteria. Hernandez et al.32 recorded that L. plantarum TF 711 had the highest inhibitory activity on Gram-positive bacteria such as B. cereus, Clostridium sporogenes and S. aureus, as well as on Gram-negative bacteria such as Shigella sonnei and K. pneumoniae. Yamato et al.33 noted that the inhibitory spectrum of bacteriocin produced by different species of Lactobacilli varies greatly and most inhibited Gram-positive bacteria. Zago et al.34 also noted that Lactobacillus isolates showed antilisterial activity and also inhibited Gram-positive bacteria better than Gram-negative bacteria.
Evaluation of bile salt resistance: One of the requirements that must be fulfilled by probiotic bacteria is the ability to withstand the bile salt juice. Resistance of the isolates in bile salt juice was based on ability of their survival by counting the number of bacteria that survived after being exposed for 4 h in 0.5% bile salt. The result displayed in Fig. 2 showed that there was a decrease in viable count of YN1.1, YN1.3 and YN1.6. This shows that isolate YN1.3 was the most resistant to bile salt amongst the LAB isolates. Control L. acidophillus FNCC also showed resistant to bile salt, because it only decreased by 4.76 (log CFU mL–1).
The reduction in total viable counts after being exposed to 0.5% bile salt solution was because of the lysis of cell. Bile acid has detergent properties so that it can damage cell walls35, resulting in cell death. The concentration of 0.5% bile salt is equivalent to the biological concentration of bile salts in duodenum36. Lactic acid bacteria from the genera Lactobacillus and Bifidobacterium have resistance mechanism to survive in bile salts by being able to efflux bile salts because of having special proteins for this purpose37. This study showed that there was a substantial decrease in total bacteria after exposure. However, half of them survived the bile salt solution. Karimpour et al.38 reported that LAB isolated from Iranian dairy product showed a greater ability to survive in bile salt and low pH. Belicova et al.39 stated that L. plantarum isolated from cheese was able to grow in 0.3% bile salt. Papadimitriou et al.40 suggested the importance of stationary-phase cells in considering probiotic bacteria because in this phase the bacteria are more resistance than log-phase cells. This is because when the bacteria are actively growing, many stress resistance mechanisms are switched on.
Evaluation of low pH resistance: The resistance of each isolate and control bacteria to withstand acidic environment was conducted by challenging the bacteria in low pH liquid media after 4 h of incubation. The result in Fig. 3 showed that there was a decrease in viable count of YN1.1, YN1.3 and L. acidophillus FNCC. The isolate YN1.6 did not survive in acidic environment since there was no viable count of bacteria after being exposed in media with pH 3.
The resistance of LAB to low pH is also an important selection criterion for probiotic. The gastric juice in the stomach destroys most ingested microorganisms.
| Table 4: | Identification of LAB species using API test CHL 50 and 16S rRNA gene |
 | |
Wang et al.41 reported that the survival rate of Lactobacillus strains after exposure to pH 2.5-4.0 was not influenced, however it dropped with the decreasing of pH. In this research, isolates YN1.1 and YN1.3 were more resistant to acidic environment than YN1.6. McDonald et al.42 noted that L. plantarum had the ability to grow at the high acid condition and this may be due to their high acid tolerance. According to Yousef and Juneja43, the resistance of LAB isolates may be because of the function of ATPase to maintain intracellular pH and protect the cells during exposure to acid by transferring protons out of the cell membrane. While Srinu et al.44 reported that L. plantarum 20 showed good survivability at low pH of 1.5-3.5.
The decrease in the number of bacteria in the acidic environment was much higher than that of in bile salt solution. Isolates YN1.1 and YN1.3 showed more tolerance to acids and bile salt compared to YN1.6. Even though some LAB such as L. plantarum have the ability to grow and colonize on intestinal tract of humans and animals45, however their survivability decreased by 53.8% in46 pH 1.5. The difference in the survivability amongst all isolates suggested that not all of the cells had the same resistance capability to strong acids. Vernazza et al.47 reported that LAB exposed to pH 2 died, but survived in pH 3 and 4. This opens up the opportunity to conduct further research to encapsulate LAB which has the potential as probiotics.
Identification of potential probiotics using API CHL 50 and 16S rRNA gene: Table 4 shows the result of identification of LAB isolates using API CHL 50 test which complemented with molecular identification using 16S rRNA gene. Fermentation profiles were interpreted by comparing all results obtained for the tested isolates with information from the computer aided database, apiweb™ identification software with database (V5.1) (https://apiweb.biomerieux.com). Based on API results, all isolates were L. plantarum with the similarity of 89, 99.9 and 97.3%, for YN1.1, YN1.3 and YN1.6, respectively. Using Ez-Taxon database to compare the obtained sequence with reference sequences, it was confirmed that isolate YN1.1 was L. senmaizukei (80.56% similarity), while isolates YN1.3 and YN1.6 were L. plantarum with 97.27 and 97.73% similarity, respectively.
Using 16S rRNA gene marker, Kim and Chun48 stated that 98.65% similarity was currently recognized as the cutoff for delineating species. While Vetrovsky and Baldrian49 suggested delineation cutoff for species was <97 and <94% for genera. This is because at 97% similarity, there will be 21.3% of OTUs contained sequences of multiple species. This current study posed the ambiguous identification results, except for YN1.3 which was confirmed as L. plantarum using both identification method. Because of the low similarity to Lactobacillus using both method, it is likely that isolate YL1.1 does not belong to Lactobacilli group. Further investigation needs to be conducted to identify the isolates YN1.1 and YN1.6.
Cho et al.50 reported that the lactobacilli associated with a fermented goat milk product from Tajikistan were more varied, among others were L. plantarum, L. pentosus, L. delbrueckii, L. helveticus and L. paracasei. Zhang et al.51 found that LAB species in traditional natural fermented goat milk included L. delbrueckii, L. acidophilus, Streptococcus thermophilus, Leuconostoc mesenteroides, Pediococcus acidilactici and Aerococcus urinaeequi. Mathara et al.45 reported that L. plantarum was the most dominant species among LAB strains isolated from traditional fermentation milk in Kenya. Apparently different location gave different results, as this might be contributed by feed given to livestock.
CONCLUSION
The results concluded that culture of isolates YN1.1, YN1.3 and YN1.6 demonstrated potential antimicrobial activity against several spoilage and pathogenic bacteria due to the production of organic acids. The isolate YN1.3 and YN1.1 also showed resistance to acidic environment and bile salt solution. The isolate YN1.3 was confirmed as L. plantarum. This study recommends that isolate YN1.1 and YN1.3 have the potential to be developed as probiotic candidates.
SIGNIFICANCE STATEMENT
This study discovered that Lactobacillus plantarum YN.1.3 isolated from spontaneous goat milk fermentation showed antimicrobial activities, withstood the acidic environment and bile salt solution, as well as produced organic acids. This bacterium can be further investigated as potential probiotic.
REFERENCES
- Holzapfel, W.H., P. Haberer, R. Geisen, J. Bjorkroth and U. Schillinger, 2001. Taxonomy and important features of probiotic microorganisms in food and nutrition. Am. J. Clin. Nutr., 73: 365S-373S.
PubMedDirect Link - Boonmee, M., N. Leksawasdi, W. Bridge and P.L. Rogers, 2003. Batch and continuous culture of Lactococcus lactis NZ133: Experimental data and model development. Biochem. Eng. J., 14: 127-135.
CrossRefDirect Link - Rastall, R.A., G.R. Gibson, H.S. Gill, F. Guarner and T.R. Klaenhammer et al., 2005. Modulation of the microbial ecology of the human colon by probiotics, prebiotics and synbiotics to enhance human health: An overview of enabling science and potential applications. FEMS Microbiol. Ecol., 52: 145-152.
CrossRefDirect Link - Ahmed, A.I., A.A. Mohammed, B. Faye, L. Blanchard and S.A. Bakheit, 2010. Assessment of quality of camel milk and gariss, North Kordofan State, Sudan. Res. J. Anim. Vet. Sci., 5: 18-22.
Direct Link - Francois, Z.N., K.P. Marie, T.A.H. Noelle and G.W.R. Emeric, 2013. Antimicrobial activity of a Bacteriocin produced by Lactobacillus plantarum 29V and strain's viability in palm kernel oil. Int. J. Nutr. Food Sci., 2: 102-108.
CrossRefDirect Link - Olanrewaju, O., 2007. Antagonistic effect of Lactobacillus isolates from kunnu and cowmilk on selected pathogenic microorganisms. Internet J. Food Saf., 9: 63-66.
Direct Link - Rojo-Bezares, B., Y. Saenz, P. Poeta, M. Zarazaga, F. Ruiz-larrea and C. Torres, 2006. Assessment of antibiotic susceptibility within lactic acid bacteria strains isolated from wine. Int. J. Food Microbiol., 111: 234-240.
CrossRefDirect Link - Prachyakij, P., J. Schnurer, W. Charernjiratrakul and K. Kantachote, 2007. Selection and identification of lactic acid bacteria that inhibit yeast contaminants isolated from fermented plant beverages. Songklanakarin J. Sci. Technol., 29: 211-218.
Direct Link - Rouse, S., D. Harnett, A. Vaughan and D. van Sinderen, 2008. Lactic acid bacteria with potential to eliminate fungal spoilage in foods. J. Applied Microbiol., 104: 915-923.
CrossRefPubMedDirect Link - Sim, K.Y., F.Y. Chye and A. Anton, 2012. Probiotic potential and antimicrobial activities of microorganisms isolated from an indigenous fish sauce. Borneo Sci., 31: 49-55.
Direct Link - Nes, I.F., D.B. Diep, L.S. Havarstein, M.B. Brurberg, V. Eijsink and H. Holo, 1996. Biosynthesis of bacteriocins in lactic acid bacteria. Antonie Leeuwenhoek, 70: 113-128.
CrossRefDirect Link - Lei, V. and M. Jakobsen, 2004. Microbiological characterization and probiotic potential of koko and koko sour water, African spontaneously fermented millet porridge and drink. J. Applied Microbiol., 96: 384-397.
CrossRefDirect Link - Soomro, A.H., T. Masud and K. Anwaar, 2002. Role of Lactic Acid Bacteria (LAB) in food preservation and human health-A review. Pak. J. Nutr., 1: 20-24.
CrossRefDirect Link - Tharmaraj, N. and N.P. Shah, 2009. Antimicrobial effects of probiotics against selected pathogenic and spoilage bacteria in cheese-based dips. Int. Food Res. J., 16: 261-276.
Direct Link - Walter, J., 2008. Ecological role of lactobacilli in the gastrointestinal tract: Implications for fundamental and biomedical research. Applied Environ. Microbiol., 74: 4985-4996.
CrossRefDirect Link - Tallei, T.E., Fatimawali and A.M. Sumual, 2019. Potential next-generation probiotics isolated from Romaine lettuce (Lactuca sativa var. longifolia Lam.) fermented brine. Proceedings of the 10th International Seminar of Indonesian Society for Microbiology & 12th Congress of Indonesian Society for Microbiology, August 29-30, 2019, Surakarta, Indonesia.
Direct Link - Bevilacqua, A.E. and A.N. Califano, 1989. Determination of organic acids in dairy products by high performance liquid chromatography. J. Food Sci., 54: 1076-1076.
CrossRefDirect Link - Rammelsberg, M. and F. Radler, 1990. Antibacterial polypeptides of Lactobacillus species. J. Applied Microbiol., 69: 177-184.
CrossRefDirect Link - Vinderola, C.G. and J.A. Reinheimer, 2003. Lactic acid starter and probiotic bacteria: A comparative in vitro study of probiotic characteristics and biological barrier resistance. Food Res. Int., 36: 895-904.
CrossRefDirect Link - Fatimawali, B. Kepel and T.E. Tallei, 2019. Potential of organic mercury-resistant bacteria isolated from mercury contaminated sites for organic mercury remediation. Pak. J. Biol. Sci., 22: 45-50.
CrossRefDirect Link - Fernandez, M., J.A. Hudson, R. Korpela and C.G. de los Reyes-Gavilan, 2015. Impact on human health of microorganisms present in fermented dairy products: An overview. BioMed Res. Int., Vol. 2015.
CrossRefDirect Link - Zalan, Z., J. Hudacek, J. Stetina, J. Chumchalova and A. Halasz, 2010. Production of organic acids by Lactobacillus strains in three different media. Eur. Food Res. Technol., 230: 395-404.
CrossRefDirect Link - Mirzaei, E.Z., E. Lashani and A. Davoodabadi, 2018. Antimicrobial properties of lactic acid bacteria isolated from traditional yogurt and milk against Shigella strains. GMS Hyg. Infect. Control, Vol. 13.
CrossRefDirect Link - Ogunbanwo, S.T., A.I. Sanni and A.A. Onilude, 2003. Characterization of bacteriocin produced by Lactobacillus plantarum F1 and Lactobacillus brevis OG1. Afr. J. Biotechnol., 2: 219-227.
Direct Link - Liasi, S.A., T.I. Azmi, M.D. Hassan, M. Shuhaimi, M. Rosfarizan and A.B. Ariff, 2009. Antimicrobial activity and antibiotic sensitivity of three isolates of lactic acid bacteria from fermented fish product, Budu. Malaysian J. Microbiol., 5: 33-37.
Direct Link - Tambekar, D.H. and S.A. Bhutada, 2010. An evaluation of probiotic potential of Lactobacillus sp. from milk of domestic animals and commercial available probiotic preparations in prevention of enteric bacterial infections. Recent Res. Sci. Technol., 2: 82-88.
Direct Link - Buntin, N., S. Chanthachum and T. Hongpattarakere, 2008. Screening of lactic acid bacteria from gastrointestinal tracts of marine fish for their potential use as probiotics. Songklanakarin J. Sci. Technol., 30: 141-148.
Direct Link - Noordiana, N., A.B. Fatimah and A.S. Mun, 2013. Antibacterial agents produced by lactic acid bacteria isolated from threadfin salmon and grass shrimp. Int. Food Res. J., 20: 117-124.
Direct Link - Ray, B. and W.E. Sandine, 1992. Acetic, Propionic and Lactic Acids of Starter Culture Bacteria as Biopreservative. In: Food Biopreservatives of Microbial Origin, Ray, B. and M. Daeschel (Eds.). CRC Press, Boca Raton, FL., USA., ISBN-13: 9780849349430, pp: 103-136.
Direct Link - Servin, A.L., 2004. Antagonistic activities of lactobacilli and bifidobacteria against microbial pathogens. FEMS Microbiol. Rev., 28: 405-440.
CrossRefPubMedDirect Link - Hernández, D., E. Cardell and V. Zarate, 2005. Antimicrobial activity of lactic acid bacteria isolated from Tenerife cheese: Initial characterization of plantaricin TF711, a bacteriocin-like substance produced by Lactobacillus plantarum TF711. J. Appl. Microbiol., 99: 77-84.
CrossRefPubMedDirect Link - Yamato, M., K. Ozaki and F. Ota, 2003. Partial purification and characterization of the bacteriocin produced by Lactobacillus acidophilus YIT 0154. Microbiol. Res., 158: 169-172.
CrossRefDirect Link - Zago, M., M.E. Fornasari, D. Carminati, P. Bums and V. Suarez et al., 2011. Characterization and probiotic potential of Lactobacillus plantarum strains isolated from cheeses. Food Microbiol., 28: 1033-1040.
CrossRefDirect Link - Ridlon, J.M., S.C. Harris, S. Bhowmik, D.J. Kang and P.B. Hylemon, 2016. Consequences of bile salt biotransformations by intestinal bacteria. Gut Microbes, 7: 22-39.
CrossRefDirect Link - Moser, S.A. and D.C. Savage, 2001. Bile salt hydrolase activity and resistance to toxicity of conjugated bile salts are unrelated properties in lactobacilli. Applied Environ. Microbiol., 67: 3476-3480.
CrossRefDirect Link - Ruiz, L., A. Margolles and B. Sanchez, 2013. Bile resistance mechanisms in Lactobacillus and Bifidobacterium. Front. Microbiol., Vol. 4.
CrossRefDirect Link - Karimpour, F.M., F.N. Tkhruni, K.J. Karapetyan and S.H. Razavi, 2013. Certain probiotic properties of lactic acid bacteria from the Iranian dairy product “Richal”. Life Sci. J., 10: 508-512.
Direct Link - Belicova, A., M. Mikulasova and R. Dusinsky, 2013. Probiotic potential and safety properties of Lactobacillus plantarum from slovak bryndza cheese. BioMed Res. Int., Vol. 2013.
CrossRefDirect Link - Papadimitriou, K., A. Alegria, P.A. Bron, M. de Angelis and M. Gobbetti et al., 2016. Stress physiology of lactic acid bacteria. Microbiol. Mol. Biol. Rev., 80: 837-890.
CrossRefDirect Link - Wang, C.Y., P.R. Lin, C.C. Ng and Y.T. Shyu, 2010. Probiotic properties of Lactobacillus strains isolated from the feces of breast-fed infants and Taiwanese pickled cabbage. Anaerobe, 16: 578-585.
CrossRefDirect Link - McDonald, L.C., H.P. Fleming and H.M. Hassan, 1990. Acid tolerance of Leuconostoc mesenteroides and Lactobacillus plantarum. Appleid Environ. Microbiol., 56: 2120-2124.
Direct Link - Yousef, A.E. and V.K. Juneja, 2002. Microbial Stress Adaptation and Food Safety. 1st Edn., CRC Press, Boca Raton, FL., USA., ISBN-13: 9781566769129, Pages: 384.
Direct Link - Srinu, B., T.M. Rao, P.V.M. Reddy and K.K. Reddy, 2013. Evaluation of different lactic acid bacterial strains for probiotic characteristics. Vet. World, 6: 785-788.
CrossRefDirect Link - Mathara, J.M., U. Schillinger, P.M. Kutima, S.K. Mbugua, C. Guigas, C. Franz and W.H. Holzapfel, 2008. Functional properties of Lactobacillus plantarum strains isolated from Maasai traditional fermented milk products in Kenya. Curr. Microbiol., 56: 315-321.
CrossRefDirect Link - Nanasombat, S., S. Phunpruch and T. Jaichalad, 2012. Screening and identification of lactic acid bacteria from raw seafoods and Thai fermented seafood products for their potential use as starter cultures. Songklanakarin J. Sci. Technol., 34: 255-262.
Direct Link - Vernazza, C.L., G.R. Gibson and R.A. Rastall, 2006. Carbohydrate preference, acid tolerance and bile tolerance in five strains of Bifidobacterium. J. Applied Microbiol., 100: 846-853.
CrossRefDirect Link - Kim, M. and J. Chun, 2014. 16S rRNA gene-based identification of Bacteria and Archaea using the EzTaxon server. Methods Microbiol., 41: 61-74.
CrossRefDirect Link - Vetrovsky, T. and P. Baldrian, 2013. The variability of the 16S rRNA gene in bacterial genomes and its consequences for bacterial community analyses. PLoS One, Vol. 8, No. 2.
CrossRefDirect Link - Cho, G.S., C. Cappello, K. Schrader, O. Fagbemigun and F.A. Oguntoyinbo et al., 2018. Isolation and characterization of lactic acid bacteria from fermented goat milk in Tajikistan. J. Microbiol. Biotechnol., 28: 1834-1845.
CrossRefDirect Link - Zhang, W.Y., Y.Y. Yun, T.S. Sun, B. Menghe and H.P. Zhang, 2008. Isolation and identification of dominant microorganisms involved in naturally fermented goat milk in Haixi region of Qinghai, China. Ann. Microbiol., 58: 213-217.
CrossRefDirect Link