
Amany M. Haroon
National Institute of Oceanography and Fisheries, Cairo, Egypt
LiveDNA: 20.11234
Usama M. Tahoun
National Institute of Oceanography and Fisheries, Cairo, Egypt
Soaad A. Sabae
National Institute of Oceanography and Fisheries, Cairo, Egypt
Walaa T. Hamza
National Institute of Oceanography and Fisheries, Cairo, Egypt
Pakistan Journal of Biological Sciences
Year: 2020 | Volume: 23 | Issue: 7 | Page No.: 861-882







ABSTRACT
Background and Objective: Damietta branch is one of the two main branches of the Nile River (Egypt), that often inhabited by many aquatic organisms, which affect and reflect its water characteristics. This study examine the relation between submerged macrophytes, their epiphytic microalgae and bacterial communities as well as the variations in their distribution and species composition with respect to season and location. Materials and Methods: Macrophytes, epiphytes and water samples were collected from 5 sites distributed along Damietta branch. Macrophytes, epiphytes and bacterial indicators of pollution were identified using standard methods. Results: Three submerged macrophytes (Myriophyllum spicatum, Ceratophyllum demersum L. and Potamogeton crispus) and 191 epiphytic algal taxa dominated by 87 Bacillariophyta and 62 Chlorophyta were recorded with significance seasonal and spatial variations. Myriophyllum spicatum was the most frequent macrophyte (p = 100%) and represent about 100, 100, 97.4 and 64.9% kg DW m–2 of the total collected macrophytes biomass during autumn, winter, spring and summer respectively. The relation between some epiphytic algal species and specific macrophytes was evident and the high organic pollution tolerant algal species like, Melosira granulata, Nitzschia palea, Synedra ulna, Oscillatoria limosa, Microcystis aeruginosa were recorded. Results of bacteriological analysis revealed a significance difference in total viable bacterial counts developed on either 22 or 37°C, total coliform, fecal coliform, fecal streptococci and Escherichia coli attributed to the seasons and sites. Conclusion: The results indicated different relations between macrophytes, epiphytes and bacteria, which is a useful biological tool for characterization of water quality in Damietta branch for different purposes.
PDF Abstract XML References Citation
Copyright: © 2020. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Amany M. Haroon, Usama M. Tahoun, Soaad A. Sabae and Walaa T. Hamza, 2020. Biological Characterization of Water in Damietta Branch of the Nile River, Egypt. Pakistan Journal of Biological Sciences, 23: 861-882.
DOI: 10.3923/pjbs.2020.861.882
URL: https://scialert.net/abstract/?doi=pjbs.2020.861.882
DOI: 10.3923/pjbs.2020.861.882
URL: https://scialert.net/abstract/?doi=pjbs.2020.861.882
INTRODUCTION
Damietta branch is one of the two main branches of the Nile River, passes through 5 governorates with an average width of 200 m, length of about 242 km and depth varying1 from12-20 m. It has considered as the major source of water needed for municipal, agricultural, industrial and feeding fish farms which dispersed between El-Serw and Faraskour region2.
This water body is often inhabited by different aquatic macrophytes life forms (submerged, floating and emergent). Submerged macrophytes are considered as important component of the aquatic ecosystem as they play key ecological roles in river, primarily as a source of food, shelter and as habitat for abundant and diverse of many aquatic organisms3-7. In addition to their role in improving water quality by removing toxic compounds and inhibiting algal blooms through the reduction of nutrients, shading and allelopathy8-10.
Epiphytic microalgae are common constituents to determine the trophic status of aquatic ecosystems11,12. They also, act as a food source for higher trophic levels13,14 and play a significant role in the functioning of shallow ecosystems, contributing to material circulation, water quality and food web interactions in aquatic ecosystems15,16. Although the presence of epiphytic algae was dependent on aquatic macrophytes as a host17 and the relationship between epiphytic algae and macrophytes plays an important role in maintaining the function and stability of the shallow ecosystems18,19. The relation between host plants and attached algae in the natural environment is still incompletely understood20,21 and their community structure is recorded to be influenced by a number of biotic and abiotic factors22-24.
Bacteriological water qualities are important issues in any water resources management especially water used in drinking purposes. Currently, total coliforms, fecal coliforms and enterococcus spp. are used as microbial indicators for predicting water pollution25. Fecal coliform bacteria are a harmful microbial contaminant, may cause diseases such as gastroenteritis, hepatitis A, typhoid fever, dysentery and cholera so, the drinking water standard requires that the Fecal coliform bacteria be totally absent from potable water26. Presence of E. coli is an indicative of fecal contamination of water source27. Very little studies dealing with the distribution of macrophytes and epiphytes had been carried out on the Nile River and its branches28-31,21. There are also some studies of El-Fadaly et al.32, Sabae et al.33 and Sabae and Rabeh34 dealing with the microbial quality of the Nile River waters. Nothing was found in literature dealing the relation between submerged macrophytes, their epiphytic microalgae and bacterial indicator of pollution at Damietta branch of the Nile River.
The present study accounted briefly on the floristic status of the submerged macrophytic plant vegetation, their epiphytic algae and bacterial indicator of pollution throughout the Damietta Branch of Nile River, Egypt. Another aim was to put the light on the role of these aquatic organisms as biological indicator of water quality of this water body.
MATERIALS AND METHODS
Sampling sites: Five sampling sites distributed along the Damietta branch of the Nile River were selected (Fig. 1, Table 1).
Macrophytes and epiphytes collection and identification: The sampling program was carried out over four season period extended from autumn (2017) to summer (2018) at the previously mentioned sites (Table 1). At each site a quadrates (50×50 cm) were used and the submerged macrophytes were handily collected from each quadrate and stored in river water before taking to the laboratory. In the laboratory the plants of each quadrate were separated into different taxa, identified based on Tackholm35.
For epiphytes collection, each collected macrophytes sample was placed separately in plastic bottles filled with deionized water. The periphyton on the macrophytes sections was scrapped off many times and the plant sections were washed with deionized water several times to completely detached all the attached algae. Epiphytes were separated from the plants by vigorous shaking for 3 min, the process was repeated at least three times and then the epiphyte suspension was poured into 1 L glass cylinder. The periphyton algal suspension was passed via a 300 μm mesh to avoid contamination by small macrophyte fragments36 poured into 1 L glass cylinder and the volume of 1 L. was completed. Lugol's iodine solution was added until the colour changed to faint tea colour. Attached algal cells were allowed to settle for 5 days then completed as mentioned above. Species identification and counting were performed in an inverted light microscope (Zeiss, Axiovert 25C) at 10· eyepiece and 400· objectives. The species were identified using the methods from previous studies37-42.
| Table 1: | Sampling sites at Damietta branch |
 | |
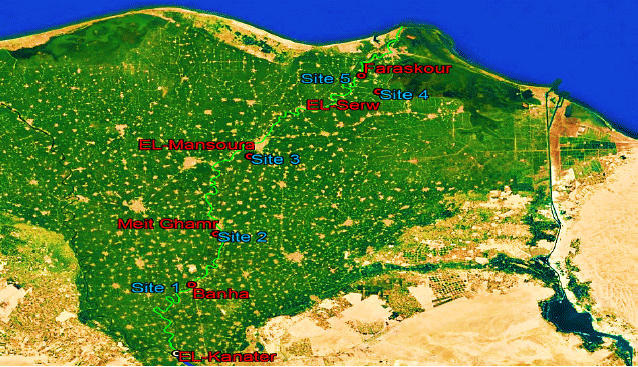 | |
| Fig. 1: | Map of Damietta branch with the selected sampling sites (Google Earth) |
Quantitative study of vegetation cover: After removing the epiphytic algae, samples of each macrophyte species were weighted for estimation of their biomass production and the results were expressed43 as kilo gram wet weight m–2 (kg wet wt m–2). This is followed by drying in shade and drying at 100°C for biomass estimation as kilo gram dry weight m–2 (kg dry wt m–2). Frequency (percent presence) of plants was calculated from the number of quadrats of occurrence for a species and the total number of quadrats sampled.
Bacteriological analysis
Sampling: Twenty water samples were collected seasonally from Damietta branch at autumn 2017 to summer 2018 at the 5 selected sites. The samples were collected using sterilized glass bottles. The collected samples were brought in an iced insulated container during transport and delivered to the analytical laboratory for microbial analyses.
Methods of analysis: All bacteriological analysis of samples was carried out according to APHA44, Samples were analyzed for TVBC at 22 and 37°C by poured plate techniques. The TC, FC and FS using MPN techniques and E. coli enumerations take placed by membrane filtration technique.
Poured plates technique: Total bacterial load growing on nutrient agar medium after decimal dilution of samples, the inoculated plates were incubated44 for 24-48 h at 37°C and 48-72 h at 22°C.
Most probable number (MPN) techniques: The decimal dilution of samples were used to inoculate 5 tubes, each tube containing 5 and 10 mL of suitable medium at single strength and double strength respectively, the tube were s incubated at 37°C for 2 days. The positive presumptive tubes were used to inoculate the confirmed test as following44:
| • | Total coliform bacteria (TC): the media used at presumptive test were Lauryl tryptose broth media, tubes showed gas and acid were positive test and for confirmation test inoculate using brilliant green lactose bile broth medium. Production of gas and acid considered a positive confirmed test |
| • | Fecal coliform bacteria (FC): From positive brilliant green lactose bile broth medium tubes inoculation in the EC broth tubes were occurred, incubated at 44.5°C for 1 day, gas production indicated positive result, for confirmation the bacteria streaked on Eosin Methylene blue agar medium (EMB), incubated at 37°C for 1 day. Metallic sheen colonies considered as a positive result |
| • | Fecal streptococci bacteria (FS): The media used for presumptive test of FS detection are Azide dextrose broth media turbid tubes were positive, for confirmation, inoculum into ethyl violet azide broth medium, (EVA) were used, incubation at 37°C for 2 days and turbid tube were considered positive results |
Enumeration of Escherichia coli: Samples were filtrate using bacterial filter paper (0.45 μm pore size), the filter transferred into EMB agar plates, incubated at 44.5°C for 1 day. Then filters were examined for typical colonies (2-3 mm in diameter, smooth with entire edge and green metallic sheen). Colonies were counted. Colonies purified by streaking on nutrient agar plate and subject to microscopic examination and biochemical tests for confirmation44.
Statistical analysis: The data recorded in this study were subjected to Pearson’s correlation analysis and principal component analysis (PCA) in order to evaluate the relationships between the 3 recorded aquatic macrophytes, their epiphytic microalgae (dominant species), bacteria and different water characteristics of the study area using XL Stat (2016) program.
RESULTS AND DISCUSSION
Vegetation analysis: Damietta branch of Nile River in Egypt is obviously populated by different communities of aquatic macrophytes that spread very rapidly and fill up the whole water body in some ecological sites. Three submerged macrophytes species Myriophyllum spicatum L., Ceratophyllum demersum L. and Potamogeton crispus were recorded throughout the study period (Table 2). This is coming in agreement with the study of El-Amier et al.30, they recorded similar number of species in Damietta branch (3 species), but higher than that found at El-Rayah Al-Behery (2 species) by Haroon and Hussian31. Lower than that mentioned by Zahran28 who recorded the presence of 21 of submerged macrophyte species, dominated by 8 species in the Nile River as a whole and also lower than that detected by Hussian and Haroon21 they recorded the existence of 5 submerged macrophyte species related to 3 genera in the Nile River. This variation in species numbers compared with the previously mentioned results could be related to different abiotic and biotic factors such as sampling sites, environmental conditions, as well as the effect of human impact on water bodies.
Throughout the study period the distribution and abundance of recorded macrophytes species was being affected by both season and sampling site (Table 2). Autumn and winter showed the poorest seasons in species number, where Myriophyllum spicatum L. was the only recorded species found in all sites and represent p =100% of the total collected macrophytes sample. While spring and summer recorded the highest species number seasons (3 species). In this study Myriophyllum spicatum L. was recorded as the most abundant species during spring present in 80% of the macrophytes sampling sites. However both Potamogeton crispus and Ceratophyllum demersum L. were the least frequent species present in 20% of the macrophytes sampling sites. Like the other three seasons Myriophyllum spicatum L. was found as the most abundant species registered in all sites (p = 100%). However, Potamogeton crispus was found in 2 sites (site 1 and 3, p = 40%) and Ceratophyllum demersum L. was recorded at one site (site 4, p =20%).
Vegetation standing crop: Biomass estimation is an important tool in aquatic plant research for studies such as species distribution and abundance, succession and assessment of weed management operations45. As shown in results (Fig. 2a, b) during autumn, winter and at sites 3 and 4 M. spicatum shows it is highest standing crop value (9.49 and 23.22 kg wet wt m–2, respectively).
| Table 2: | Floristic composition of the different ecological sites in Damietta branch of the Nile River, Egypt |
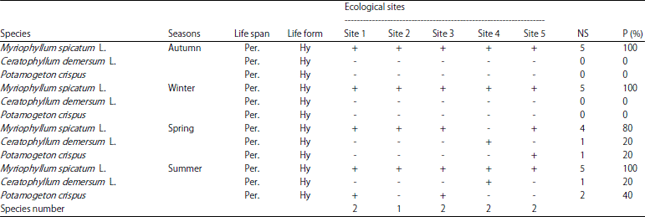 | |
NS: Number of sites in which the plants is recorded, P (%): Presence percentage, life span: Per.: Perennials, Life form: Hy: Hydrophytes, +: Presence and -: Absence | |
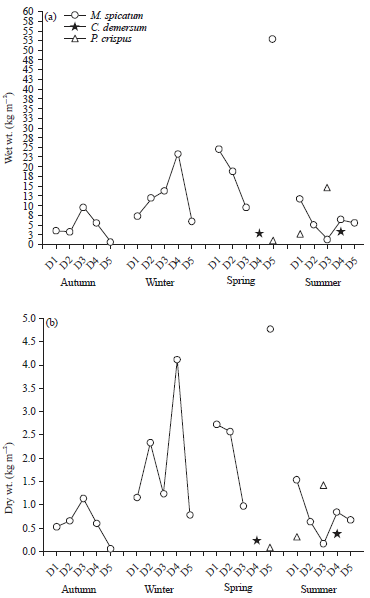 | |
| Fig. 2(a-b): | Seasonal vegetation cover percentage for the recorded submerged Macrophytes species |
However, during spring the maximum growth production value (52.78 kg wet wt. m–2) was detected for the same species from site 5. In summer the submerged macrophyte species P. crispus recorded the highest standing growth values (14.30 and kg dry wt. m–2) compared with the other 2 species. During the whole study period M. spicatum showed the highest biomass production value and constituent 100, 100, 96.89 and 59.0% wet wt of the total collected macrophytes biomass (Fig. 3).
Biomass production of submerged aquatic macrophytes is regulated by climatic and environmental factors like: Temperature, solar irradiance, day length, humidity, wind and water physicochemical characteristics46-49. In the present investigation, the maximum biomass production value for M. spicatum (52.78 and 4.76 kg dry wt. m–2) was found during spring at site 5, however, C. demersum (3.35 kg wt. m–2) and P. crispus (14.30 kg wet wt. m–2) attended their highest production values during summer at sites 4 and 3, respectively (Fig. 2a). According to scientists46,48, the summer highest production values of these species may be due to the plant avoids the competition for light, nutrient and space with the aquatic plants which have their peak of growth in summer. These data were relatively lower than that reported by Hussin and Haroon21 during cold season for M. spicatum (66.00 kg wet wt. m–2) and C. demersum 18.85 kg wet wt. m–2 from Nile River Egypt, but higher than the value of 3.35 kg wet wt. m–2 that was reported for P. crispus. On the other hand, lower values were recorded by El-Sheekh et al.29 and Shaltout et al.49 for Ceratophyllum demersum (0.036-1.094 kg wet wt. m–2) and P. crispus (0.400 kg dry wt. m–2) from different sites of the Nile Delta, respectively.
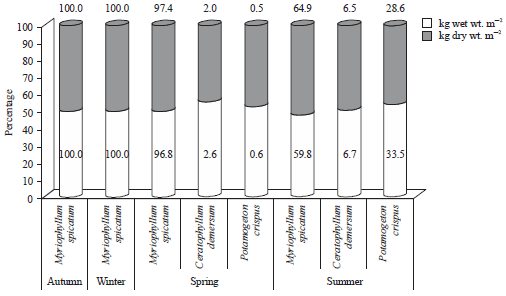 | |
| Fig. 3: | Mean percent of each macrophyte species biomass in relation to the total collected macrophytes biomass during study period |
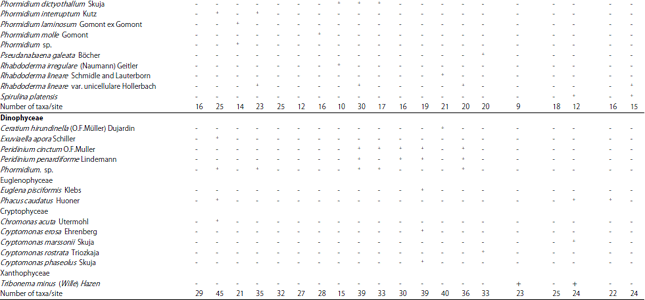
Epiphytic algae on the three aquatic plants in Damietta branch: Changes in the submerged aquatic macrophyte community can be considered as one of the most important factors that affecting changes in the epiphytic algal community structure50. During the study period a total of 191 algal taxa were identified on the three collected macrophyte species (Table 3, 4, Fig. 4). Which being higher than that recorded by Deyab et al.51 and Ahmed52 they recorded 141 epiphytic algal species on Phragmites australis and 105 attached algal species in the Nile River and irrigation canals at Sohag district. At the same time the present results were relatively lower than that detected by Hussian and Haroon21 they recorded 200 epiphytic species on submerged aquatic macrophytes of the Nile River (Egypt), in which Bacillariophyceae were detected as the dominant algal group (92.3%) on all aquatic macrophytes, with 119 taxa related to 31 genera. In this investigation Bacillariophyceae was recorded as the dominant algal group 87 algal species followed by Chlorophyta 62, Cyanophyceae 29, Dinophyceae 5, Euglenophyceae 2, Cryptophyceae 5 and Xanthophyceae 1 species. This variation may be due to the variation in nutrients concentration, released substances from macrophyte and water pollution53,54. As shown in Table 3 and 4 the percentage of epiphyte algae varied according to the host plant species, stations and seasons. Diatoms were the dominant group with the highest presence percentage during winter and summer seasons (91.94 at site 4 and 96.23% at site 5, respectively), however Chlorophyceae attended their highest presence during spring (40.16% at site 5) and Cyanophyceae increased in spring (11.74% at site 3). At the same time xanthophyceae was recorded only during summer.
The dominancy of diatoms among other attached algal classes was observed in different Damietta sites (Table 3). This is in accordance with Dere et al.55, Albay and Aykulu56 and Deyab et al.51. That dominancy may be due to their ability to resist different environmental stresses such as deficiency of light and their ability to grow on different aquatic substrate due to their possessing of silicate cell wall57,58. Chlorophyceae followed diatoms in importance and Cyanophyceae increased in spring and summer. As shown in Fig. 4 the percentage of the most common epiphytic algal species recorded was varied seasonally. In addition some algal species appeared during all seasons of study period such as Cyclotella ocellata Pant and Syndra ulna (Nitzsch) Her., while Scenedesmus quadricauda (Turpin) Brébisson, Lyngbya limnetica Lemmermann and Microcystis aeruginosa Kützing species appeared during most seasons of study period as shown in results (Table 4) which may be due to their wide range in temperature tolerance as pointed out by Hickman and Klarer59.
Organic carbon released by macrophytes is a major substrate for epiphytic bacteria60,61. While, little phosphorus is released by living aquatic plants62 and epiphytic algae obtain less than 10% of their phosphorus from the host macrophyte63. On the other hand, epiphyte algae may rapidly assimilate phosphorus released from decomposing macrophytes64 and provide food for grazers. According to the abundance and seasonal occurrence of the three hydrophytes namely Myriophyllum spicatum, Ceratophyllum demersum and Potamogeton crispus along the study area, their epiphytic microalgae were analyzed. Seasonal and spatial data about the occurrence of different epiphytic microalgae on the three macrophytes are given in Table 4.
| Table 3: | Abundance (%) of the different recorded classes of epiphytic algae attached to macrophytes in Damietta branch during study period |
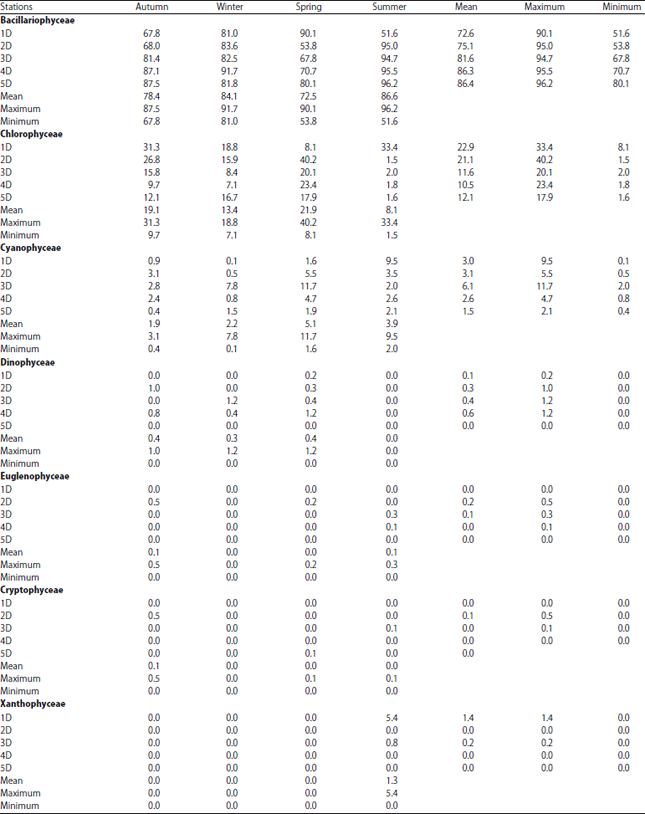 | |
| Table 4: | Seasonal and spatial occurrence of different epiphytic microalgae of the selected hydrophytes along the study area |
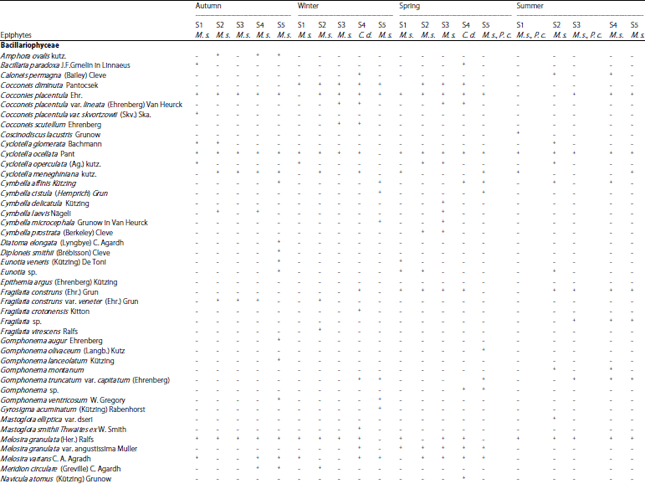  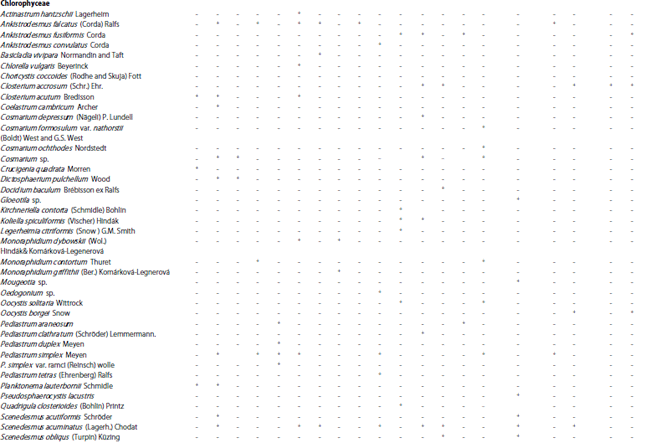 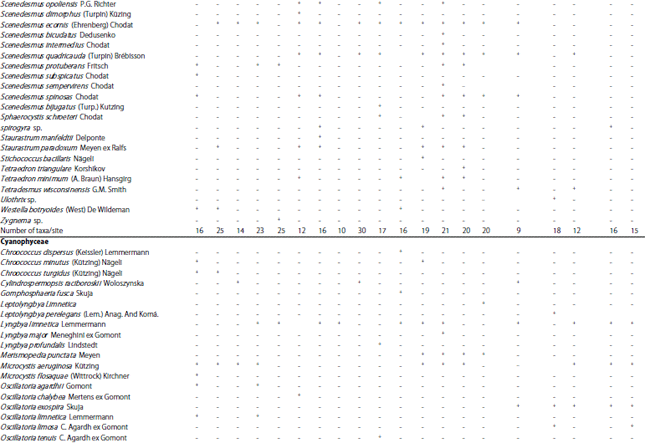  | |
M.s.: Myriophyllum spicatum, C.d.: Ceratophyllum demersum, P.c.: Potamogeton crispus, +: Presence, -: Absence, S: Site | |
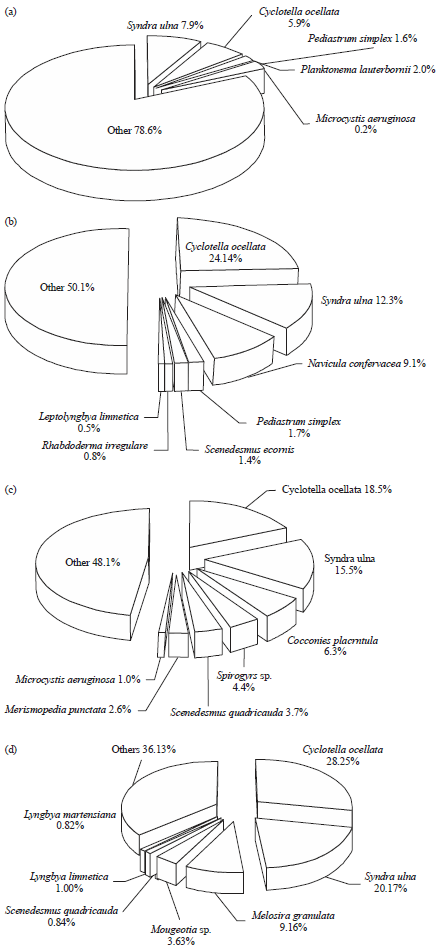 | |
| Fig. 4(a-d): | Average of the percentage of the most common epiphytic algal species recorded in Damietta branch during study period, (a) Autumn 2017, (b) Winter 2018, (c) Spring 2018 and (d) Summer 2018 |
These data indicated that epiphytes were largely dependent on season, plant species and sampling sites. During the study period there are some epiphytic algal species occurred with specific submerged macrophyte species (Table 4) like: Amphora ovalis kutz., Diatoma elongata (Lyngbye) C. Agardh., Diploneis smithii (Brébisson) Cleve), Fragilaria construns var. veneter (Ehr.) Grun, Meridion circulare (Greville) C. Agardh, Navicula punctulata W. Smith, Nitzschia sigmoidea (Nitzsch) W. Smith, Synedra actinastroides Lemmermann, Syndra acus Kutz, (Bacillariophyceae), Dictosphaerium pulchellum Wood, Monoraphidium spp., Zygnema sp., ( Chlorophyceae) Chroococcus turgidus (Kützing) Nägeli and Phormidium spp. (Cyanophyceae) were occurred only in autumn and winter, which indicated the specificity of these spp. to M. spicatum. While Cryptophyceae and Xanthophyceae appeared only in spring and summer seasons with the occurrence of C. demersum and. P. crispus. The same result was observed by Albay and Aykulu56 and Ahmed52.
Bacteriological analysis
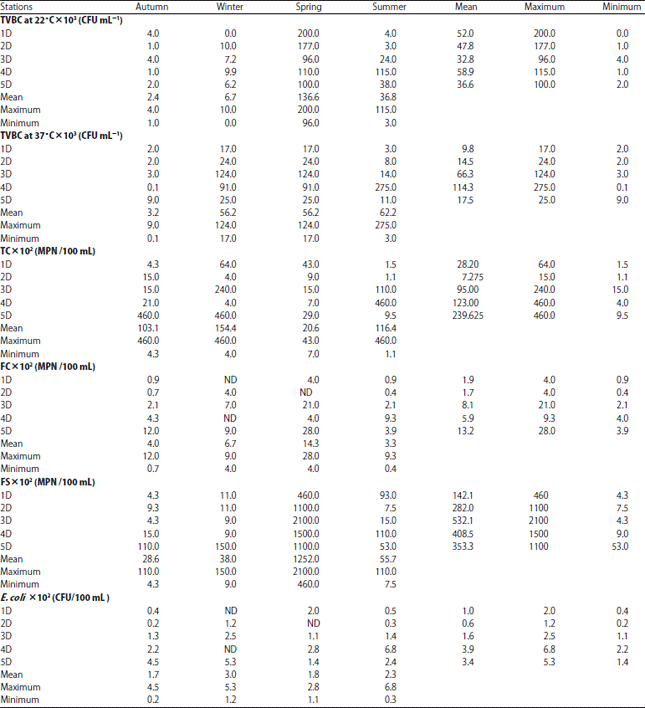
Total viable bacterial count: Bacteriological water qualities are important issues in any water resources management especially water used in drinking purposes. During the study period, the total viable bacterial counts developed on either 22 or 37°Cat Damietta branch water were varied significantly with season and sampling site (Table 5). They were particularly higher in spring and summer (up to 200×103 and 275×103 CFU mL–1, respectively) compared to the other two seasons. In addition, water samples from sites 1 and 4 were found to have the minimum numbers of TVBC developed on either 22 or 37°C (0 and 0.1× 103 CFU, respectively).
Bacterial indicators: Now-a-days, with increasing of human activities, the monitoring of fresh water pollution should be occurred for health and environmental protection. Therefore, the bacteriological characteristics for water quality of Damietta branch are studied.
Results of bacterial indicators for water samples during four seasons are illustrated in Table 5 and varied depending on the site and the season. Total coliform, Fecal coliform and Fecal streptococci at water samples of Damietta branch were in the range of (1.1×102 to 460×102), (0.4×102 to 28×102) and (4.3×102 to 2100×102) MPN/100 mL water, respectively and the numbers of E. coli were in the range of 0-6.8×102 (CFU/100 mL water). Bacterial indicators of pollution were monitored in the studied area with the highest numbers in warm seasons and the obtained results indicated of the suitability of the water for irrigation not for drinking.
Similar results were concluded for bacterial indicators of Nile River and two branches by many Authors34,65,66. Results of coliform group in the present study are not in a compliance with the Egyptian standards67 for drinking water quality where recommended that the drinking water samples should be free from bacterial indicators and pathogens (e.g., TC, FC) in 100 mL water.
High numbers of bacterial indicators at Damietta branch might be explained by direct effects of domestic and agricultural wastes discharge from different sources at the urbanized surrounding area and these results agree with Sabae and Rabeh34.
In conclusion, according to WHO68 the results of bacteriological investigation indicate that, the quality of Damietta branch water is subjected to sewage pollution and not acceptable for drinking purposes.
Relation between macrophytes, epiphytes, bacterial indicator of pollution and water characteristics: In this study the principal correspondence analysis (PCA) and simple linear correlation coefficient and at α = 0.05 show different relations between macrophytes, epiphytes, bacteria and different water parameters (Fig. 5, 6, Table 6). Relations between epiphytic microalgae and the host aquatic plant show the whole ecosystem character and the ecosystem responses to the altering environmental conditions69. There are some epiphytic algal classes were closely correlated with special macrophytes species (Fig 5a). Bacillariophyceae was associated with the three macrophytes species, however, Cryptophyceae and Euglenophyceae were significantly correlated with P. crispus (r = 0.350 and 0.557, respectively) and Dinophyceae was more associated with C. demersum.
The correlation statistical analysis in Damietta branch (Fig. 5a) show a significance positive relation between TVBC at 22°C and the submerged macrophytes species M. spicatum and C. demersum (r = 0.411 and 0.360, respectively). In addition it shows a very weak positive relation with Bacillariophyceae, Chlorophyceae, Cyanophyceae and a significance relation with the algal species Cocconies placentula Ehr (r = 0.208. 0.284, 0.231 and 0.446, respectively). TVB°C at 37°C showed a significant positive relation with M. spicatum, C. demersum, Bacillariophyceae, Cyanophyceae and Dinophyceae. Where, the total coliform bacteria attended their highest values during autumn and winter associated with the presence of M. spicatum. However, during summer the highest number was found in site 4 occupied by M. spicatum and C. demersum.
| Table 5: | Bacteriological analysis of water samples at Damietta branch |
 | |
TVBC: Total viable bacterial count, TC: Total coliform, FC: Fecal coliform, FS: Fecal streptococci | |
Both Fecal Coliform and Fecal streptococci bacteria recorded their highest numbers during spring at sites 5 and site 3 associated with the presence of M. spicatum, P. cripus and M. spicatum. However E. coli recorded their highest number at site 4 during summer associated with the presence of M. spicatum and C. demersum.
Several studies have shown reductions in the species richness of submerged and floating-leaved macrophyte communities with increasing nutrient levels70,71.
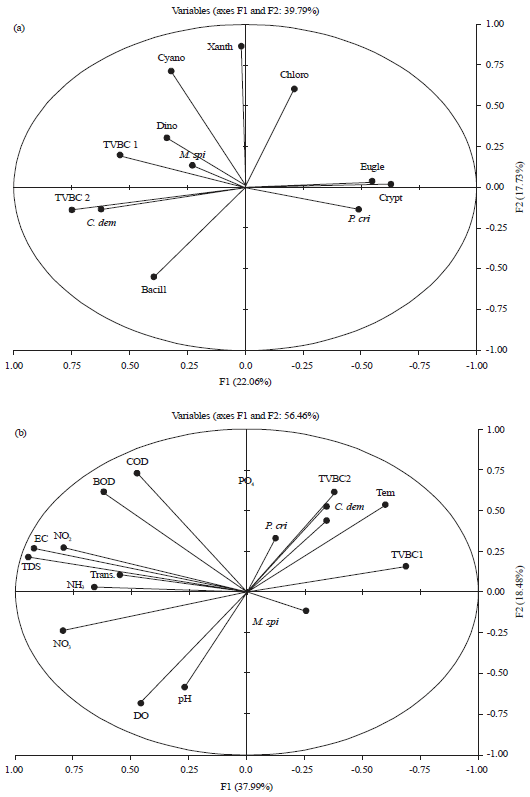 | |
| Fig. 5(a-b): | Principal component analysis (PCA) (Axis 1 and 2) performed on relation between submerged macrophytes, epiphytic algal classes, TVB°C at 22, 37°C and some water variables throughout the study period |
| M. spi: Myriophyllum spicatum, P. cri: Potamogeton crispus, C. dem: Ceratophyllum demersum, TVBC1:TVBC at 22°C, TVBC2: TVBC at 37°C, Bacill: Bacillariophyceae, Chloro: Chlorophyceae, Cyano: Cyanophyceae, Dino: Dinophyceae, Crypto: Cryptophyceae, Eugle: Euglenophyceae, Xanth: Xanthophyceae | |
The relation between macrophytes distribution and water physicochemical characteristics (Fig. 5b) shows that the existence of submerged macrophytes species M. spicatum was associated with decreasing levels of EC, TDS, DO, BOD and nitrite (r = -0.314, -0.247, -0.230, -0.229 and -0.224, respectively) without any clear relation with water temperature which explain its appearance throughout the study period. The two other macrophytes species C. demersum and P. crispus were negatively correlated with some water variables like: Trans., pH, DO and NO2, however, they were positively correlated with temperature which discussed the appearance of these 2 species during hot seasons only (spring and summer).
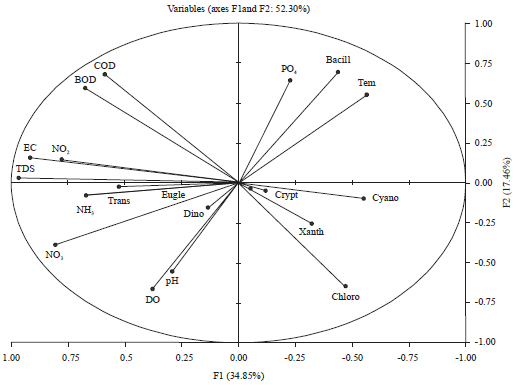 | |
| Fig. 6: | Principal component analysis (PCA) (Axis 1 and 2) performed on different algal classes with different water variables |
| Bacill: Bacillariophycae, Chloro: Chlorophyceae, Cyano: Cyanophyceae, Dino: Dinophyceae, Crypto: Cryptophyceae, Eugle: Euglenophyceae, Xanth: Xanthophyceae | |
Uedeme-Naa et al.72 mentioned the presence of positive relation between aquatic macrophytes and water temperature, as the photosynthetic activity is increased by increasing of temperature.
Results in Fig. 2 and 5b show that, the lowest and the highest biomass production values of P. crispus were recorded in summer season at site 1 associated with the highest transparency, highest DO and low value of EC and low PO4 and at site 3 associated with relatively low transparency and high values of NO3, PO4 and lowest DO respectively. This finding was supported by Li73 he noted the ability of this species to tolerate wide ranges of water characteristics. Significance positive relation between the submerged recorded macrophytes species (M. spicatum and C. demersum) and water PO4 was observed (Fig. 5b). This agrees with the results of other investigators, like Haroon and Hussian31 they recorded a positive relation between M. spicatum and PO4 and Frankouich et al.74 they mentioned the correlation between macrophyte growth, distribution and nutrient rich environments especially nitrate and phosphate which have been noted to favour the growth of macrophytes. Moreover, the highest biomass production value of C. demersum at site 4 during summer associated with the relatively high values of TDS, NO3, NH3, PO4 and low transparency indicate the ability of this species to survive in water of high nutrients. So and as recorded by Ali et al.75 this plant could be regarded as nutrient tolerant hydrophyte.
Regarding the relation between water physicochemical characteristics and different algal classes (Fig. 6, Table 6). Bacillariophyceae and its dominant species were more affected by temperature and PO4 (r = 0.454 and r = 0.448, respectively at α = 0.05). At the same time it was negatively affected by TDS r = -0.505 and NO3 r = -0.689. Chlorophyceae shows a significance negative relation with EC, TDS, BOD, COD and NO2 (r = -0.573, -0.449, -0.640, -0.546 and -0.466, respectively). Cyanophyceae was negatively affected by water EC, TDS and NO3 (r =-0.503, -0.477 and -0.444, respectively) and Xanthophyceae was only affect by COD (r = -0.471). Different relations were observed between the dominant algal species, macrophytes and bacteria like: The significant positive relation between M. spicatum and Pediastrum simplex Meyen (0.456), Cocconies placentula Her and the submerged macrophytes species C. demersum, TVBC at 22, 37°C, Bacillariophyceae (0.628, 0.445, 0.750 and 0.693, respectively). In addition, the dominant algal species Spirogyra sp. and Microcystis aeruginosa Kützing were correlated with TVBC at 22oC and with cyanophyceae (Table 6).
| Table 6: | Simple linear correlation coefficient between the different macrophytes biomass, bacteria, different algal classes and dominant algal species |
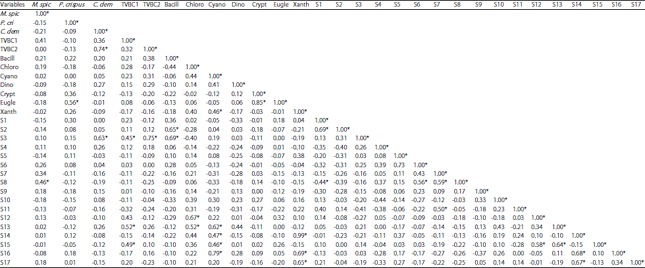 | |
*Values are different from 0 with a significance level " = 0.05, M. spi: Myriophyllum spicatum, P. cri: Potamogeton crispus , C. dem: Ceratophyllum demersum, TVBC1: TVBC at 22EC, TVBC2: TVBC2 at 37EC, Bacill: Bacillariophycae, Chloro: Chlorophyceae, Cyano: Cyanophyceae, Dino: Dinophyceae, Crypto: Cryptophycae, Eugle: Euglenophycae, Xanth: Xanthophycae, S1: Syndra ulna (Nitzsch) Ehr., S2: Cyclotella ocellata Pant, S3: Cocconies placentula Ehr., S4: Melosira varians C. A. Agradh, S5: Melosira granulata (Her.) Ralfs, S6: Navicula lanceolata Ehrenberg, S7: Planktonema lauterbornii Schmidle, S8: Pediastrum simplex Meyen, S9: Dictosphaerium pulchellum wood, S10: Scenedesmus ecornis (Ehrenberg) Chodat, S11: Scenedesmus spinosas Chodat, S12: Scenedesmus quadricauda (Turpin) Brébisson, S13: Spirogyra sp., S14: Mougeotia sp., S15: Microcystis aeruginosa Kützing, S16: Lyngbya limnetica Lemmermann, S17: Cylindrospermopsis raciboroskii Woloszynska | |
Physico-chemical analysis, macrophytes, epiphytic algal species and bacteria proved good integrated tools for reliable assessment of water quality of Damietta branch of Nile River (Fig. 5, 6, Table 6). As mentioned by Barbour et al.76 epiphytic algae can be used as a good indicators of water quality and environmental changes due to their sensitivity to external sources of pollutions. Results of the present investigation show the presence of high organic pollution tolerant algal species; Melosira granulate, Nitzschia palea, Synedra ulna Oscillatoria limosa, Microcystis aeruginosa during summer season. Similar results were recorded by Ali and El Shehawy77 they reported the presence of Microcystis aeruginosa, Cyclotella sp., Melosira granulata and Nitzschia palea along the study segment of the Nile River. In addition Scendesmus spp., which was known by its ability to grow in water polluted by heavy metals and can be used as an indicator of Eutrophication78 was also observed in various sites of the studied area characterized by it is high levels of organic matter. Moreover the diatom species Cocconeis placentula, recorded by Toporowska et al.79 as bio-indicator of slightly alkaline and meso-eutrophic water was recorded with high quantity at sites 3, 4 and 5.
The finding of this study suggested that the highest biomass production values of submerged macrophyte species M. spicatium and C. demersum during summer and spring at sites 4, 5 associated with the highest TVBC may indicate the presence of agricultural drainage sources of pollution at these areas. These findings are in accord with Ali and Soltan80 they recorded that high dry weight standing crop values of C. demersum and M. spicatum may indicate the presence of agricultural drainage sources of pollution, while high value of P. crispus may indicate the presence of industrial source of pollution.
CONCLUSION
This study indicates that the distribution and species composition of submerged aquatic macrophytes, their epiphytic algae and bacterial indicators of pollution are affected by season, environmental characteristics of the study area as well as interaction between these aquatic organisms. In addition, Damietta Branch might be subjected to different sources of biological pollution specially in summer season and at El- Serw and Faraskour, that was confirmed by prevalence of some species of either submerged macrophytes (M. spicatum, C. demersum and P. crispus) pollution tolerant algal ( M. granulate, N. palea, S. ulna, O. limosa, M. aeruginosa, Scendesmus spp. and C. placentula) and bacteria (total coliform and fecal coliform, streptococci and Escherichia coli ) which were reported previously as biological indicators of pollution.
SIGNIFICANCE STATEMENT
This study discovered that, the distribution and species composition of submerged aquatic macrophytes, their epiphytic algae and different bacterial groups are affected by different environmental variables especially water quality. That can be beneficial for using these organisms as a good biological indicators of water pollution. In addition, the allelopathic interaction between these organisms, beside the ability of submerged macrophytes to absorb nutrients in large quantities may help the researchers to determine the most suitable ways that needed for utilizing these organisms in purposes of improving water quality.
ACKNOWLEDGMENTS
The authors would like to thank Prof. Dr. Mohamed E. Goher, Head of Chemistry Lab. for providing laboratory assistance in water quality analysis. Thanks was also extend to Dr. Abd-Ellatif Mohamed Hussian for his help in statistical analysis.
REFERENCES
- Authman, M.M.N., M.A. El-Kasheif and K.A.S. Shalloof, 2009. Evaluation and management of the fisheries of Tilapia species in Damietta Branch of the River Nile, Egypt. World J. Fish Mar. Sci., 1: 167-184.
Direct Link - Barko, J.W. and W.F. James, 1998. Effects of Submerged Aquatic Macrophytes on Nutrient Dynamics, Sedimentation and Resuspension. In: The Structuring Role of Submerged Macrophytes in Lakes, Jeppesen, E., M. Søndergaard, M. Søndergaard and K. Christoffersen (Eds.)., Ecological Studies (Analysis and Synthesis), Vol. 131. Springer, New York, pp: 197-226.
Direct Link - Toft, J.D., C.A. Simenstad, J.R. Cordell and L.F. Grimaldo, 2003. The effects of introduced water hyacinth on habitat structure, invertebrate assemblages and fish diets. Estuaries, 26: 746-758.
CrossRefDirect Link - Haroon, A. and S.M. Daboor, 2009. The role of different macrophytes groups in water quality, sediment chemistry and microbial flora of both irrigation and drainage canals. World Applied Sci. J., 6: 1221-1230.
Direct Link - Sharshar, K.M. and A.M. Haroon, 2009. Comparative investigations on some biological and biochemical aspects in freshwater crayfish (Procambarus clarkii) fed on Eichhornia crassipes, Echinochloa stagnina L. and Polygonum tomentosum. Am.-Eurasian J. Agric. Environ. Sci., 5: 579-589.
Direct Link - Haroon, A.M., 2010. Evaluation of the nutritional status of Nymphaea lotus L. and Pistia stratiotes L. shoots in relation to their utilization as fish and animal feed. Egypt. J. Aquat. Res., 36: 587-595.
Direct Link - Haroon, A.M. and E.I. Abdel-Aal, 2016. Chemical composition and in vitro anti-algal activity of Potamogeton crispus and Myriophyllum spicatum extracts. Egypt. J. Aquat. Res., 42: 393-404.
CrossRefDirect Link - Kuiper, J.J., M.J. Verhofstad, E.L. Louwers, E.S. Bakker and R.J. Brederveld et al., 2017. Mowing submerged macrophytes in shallow lakes with alternative stable states: Battling the good guys? Environ. Manage., 59: 619-634.
CrossRefDirect Link - Casartelli, M.R. and C. Ferragut, 2018. The effects of habitat complexity on periphyton biomass accumulation and taxonomic structure during colonization. Hydrobiologia, 807: 233-246.
CrossRefDirect Link - Vis, C., C. Hudon and R. Carignan, 2006. Influence of the vertical structure of macrophyte stands on epiphyte community metabolism. Can. J. Fish. Aquat. Sci., 63: 1014-1026.
CrossRefDirect Link - Cook, F.S. and P.S. Coleman, 2007. Benthic diatoms in the salinas of the dry creek saltfields, South Australia. Hydrobiologia, 576: 61-68.
CrossRefDirect Link - Armitage, A.R., T.A. Frankovich and J.W. Fourqurean, 2006. Variable responses within epiphytic and benthic microalgal communities to nutrient enrichment. Hydrobiologia, 569: 423-435.
CrossRefDirect Link - Abe, S.I., K. Uchida, T. Nagumo and J. Tanaka, 2007. Alterations in the biomass‐specific productivity of periphyton assemblages mediated by fish grazing. Freshwater Biol., 52: 1486-1493.
CrossRefDirect Link - Drake, L.A., F.C. Dobbs and R.C. Zimmerman, 2003. Effects of epiphyte load on optical properties and photosynthetic potential of the seagrasses Thalassia testudinum Banks ex König and Zostera marina L. Limnol. Oceanogr., 48: 456-463.
CrossRefDirect Link - Song, Y.Z., J.Q. Wang and Y.X. Gao, 2017. Effects of epiphytic algae on biomass and physiology of Myriophyllum spicatum L. with the increase of nitrogen and phosphorus availability in the water body. Environ. Sci. Pollut. Res., 24: 9548-9555.
CrossRefDirect Link - Cattaneo, A., G. Galanti, S. Gentinetta and S. Romo, 1998. Epiphytic algae and macroinvertebrates on submerged and floating‐leaved macrophytes in an Italian lake. Freshwater Biol., 39: 725-740.
CrossRefDirect Link - Liboriussen, L. and E. Jeppesen, 2003. Temporal dynamics in epipelic, pelagic and epiphytic algal production in a clear and a turbid shallow lake. Freshwater Biol., 48: 418-431.
CrossRefDirect Link - Scheffer, M. and E.H. van Nes, 2007. Shallow lakes theory revisited: various alternative regimes driven by climate, nutrients, depth and lake size. Hydrobiologia, 584: 455-466.
CrossRefDirect Link - Buczkó, K., 2007. The occurrence of the epiphytic diatom Lemnicola hungarica on different European Lemnaceae species. Fottea Olomouc, 7: 77-84.
Direct Link - Hussian, A.M. and A.M. Haroon, 2019. Sensitivity of submerged aquatic macrophytes and their epiphytic microalgae to the different environmental variables in River Nile, Egypt. Int. J. Ecol. Environ. Sci., 45: 107-122.
Direct Link - Trochine, C., M.E. Guerrieri, L. Liboriussen, T.L. Lauridsen and E. Jeppesen, 2014. Effects of nutrient loading, temperature regime and grazing pressure on nutrient limitation of periphyton in experimental ponds. Freshwater Biol., 59: 905-917.
CrossRefDirect Link - Tóth, V.R. and S.C. Palmer, 2016. Acclimation of Potamogeton perfoliatus L. to periphyton accumulation-induced spectral changes in irradiance. Hydrobiologia, 766: 293-304.
CrossRefDirect Link - Hao, B., H. Wu, Y. Cao, W. Xing, E. Jeppesen and W. Li, 2017. Comparison of periphyton communities on natural and artificial macrophytes with contrasting morphological structures. Freshwater Biol., 62: 1783-1793.
CrossRefDirect Link - Rochelle-Newall, E., T.M.H. Nguyen, T.P.Q. Le, O. Sengtaheuanghoung and O. Ribolzi, 2015. A short review of fecal indicator bacteria in tropical aquatic ecosystems: Knowledge gaps and future directions. Front. Microbiol., Vol. 6.
CrossRefDirect Link - Al-Afify, A.D.G., U.M. Tahoun and M.H. Abdo, 2019. Water quality index and microbial assessment of lake Qarun, El-Batts and El-Wadi Drains, Fayoum province, Egypt. Egypt. J. Aquat. Biol. Fish., 23: 341-357.
CrossRefDirect Link - Zahran, M.A., 2009. Hydrophytes of the Nile in Egypt In: The Nile: Origin, Environments, Limnology and Human Use, Dumont, H.J. (Ed.)., Springer Science Business Media, Dordrecht, pp: 463-478.
Direct Link - El-Sheekh, M.M., A.M. Haroon and S. Sabae, 2018. Seasonal and spatial variation of aquatic macrophytes and phytoplankton community at El-Quanater El-Khayria River Nile, Egypt. Beni-Suef Univ. J. Basic Applied Sci., 7: 344-352.
CrossRefDirect Link - El-Amier, Y.A., M.A. Zahran and S.O. Al-Mamoori, 2015. Plant diversity of the Damietta branch, river Nile, Egypt: An ecological insight. Mesopotamia Environ. J., 1: 109-129.
Direct Link - Haroon, A.M and A.E.M. Hussian, 2017. Ecological assessment of the macrophytes and phytoplankton in El-Rayah Al-Behery, River Nile, Egypt Egypt. J. Aquat. Res., 43: 195-203.
CrossRefDirect Link - Sabae, S.Z. and S.A. Rabeh, 2007. Evaluation of the microbial quality of the River Nile waters at damietta branch, Egypt. Egypt. J. Aquat. Res., 33: 301-311.
Direct Link - Tackholm, V., 1974. Students Flora of Egypt. 2nd Edn., Cairo University Cooperative Printing Co., Beirut.
Direct Link - Cattaneo, A., T. Kerimian, M. Roberge and J. Marty, 1997. Periphyton distribution and abundance on substrata of different size along a gradient of stream trophy de Montréal. Hydrobiologia, 354: 101-110.
CrossRefDirect Link - Cleve-Euler, A., 1952. Die Diatomeen von Schweden und Finland. Band 1, 2, 3, 4 and 5. Almqvist and Wiksells Boktryckeri AB, Stockholm.
Direct Link - Patrick, R., B. Crum and J. Coles, 1969. Temperature and manganese as determining factors in the presence of diatom or blue-green algal Floras in streams. Proc. Natl. Acad. Sci. USA., 64: 472-478.
CrossRefPubMedDirect Link - Mizuno, T., 1990. Illustration of the Freshwater Plankton of Japan. 9th Edn., Hoikush Publishing Co., LOT, Japan, Pages: 353.
Direct Link - Misra, R., 1968. Ecology Work Book. Oxford and IBH Publishing Co., Calcutta, India, Pages: 244.
Direct Link - Pine, R.T., L.W.J. Anderson and S.S.O. Hung, 1989. Notes on non-destructive estimation of aquatic macrophytes biomass. J. Aquat. Plant Manage., 27: 47-49.
Direct Link - Eid, E.M., K.H. Shaltout, Y.M. Al-Sodany and K. Jensen, 2010. Effects of abiotic conditions on Phragmites australis along geographic gradients in Lake Burullus, Egypt. Aquat. Bot., 92: 86-92.
CrossRefDirect Link - Eid, E.M., K.H. Shaltout, Y.M. Al-Sodany, K. Soetaert and K. Jensen, 2010. Modeling growth, carbon allocation and nutrient budgets of Phragmites australis in Lake Burullus, Egypt. Wetlands, 30: 240-251.
CrossRefDirect Link - Eid, E.M., K.H. Shaltout and T. Asaeda, 2012. Modeling growth dynamics of Typha domingensis (Pers.) Poir. ex Steud. in Lake Burullus, Egypt. Ecol. Modell., 243: 63-72.
CrossRefDirect Link - Shaltout, K.H., E.M. Eid and T.M. El-Komi, 2016. Phytomass and nutrient value of Potamogeton crispus L. in the water courses of Nile Delta, Egypt. Rendiconti Lincei, 27: 251-259.
CrossRefDirect Link - De Souza, M.L., B.G. Pellegrini and C. Ferragut, 2015. Periphytic algal community structure in relation to seasonal variation and macrophyte richness in a shallow tropical reservoir. Hydrobiologia, 755: 183-196.
CrossRefDirect Link - Wetzel, R.G., 2001. Limnology, Lake and River Ecology. 3th Edn., Academic Press, New York, Pages: 1006.
Direct Link - Dere, Ş., D. Karacaoğlu and N. Dalkiran, 2002. A study on the epiphytic algae of the Nilüfer Stream (Bursa). Turk. J. Bot., 26: 219-234.
Direct Link - Albay, M. and G. Aykulu, 2002. Invertebrate grazer-epiphytic algae interactions on submerged macrophytes in a mesotrophic Turkish lake. Su Ürünleri Dergisi, 19: 247-258.
Direct Link - Leelahakriengkrai, P. and Y. Peerapornpisal, 2010. Diversity of Benthic diatoms and water quality of the Ping river, Northern Thailand. Environ. Asia, 3: 82-94.
Direct Link - Hickman, M. and D.M. Klarer, 1974. The growth of some Epiphytic algae in a lake receiving thermal effluent. Arch. Hydrobiol., 3: 403-426.
Direct Link - Allen, H.L., 1971. Primary productivity, chemo‐organotrophy and nutritional interactions of epiphytic algae and bacteria on macrophytes in the littoral of a lake. Ecol. Monogr., 41: 97-127.
CrossRefDirect Link - Søndergaard, M., 1983. Heterotrophic utilization and decomposition of extracellular carbon released by the aquatic angiosperm Littorella uniflora (L.) Aschers. Aquat. Bot., 16: 59-73.
CrossRefDirect Link - Barko, J.W. and R.M. Smart, 1980. Mobilization of sediment phosphorus by submersed freshwater macrophytes. Freshwater Biol., 10: 229-238.
CrossRefDirect Link - Carignan, R. and J. Kalff, 1982. Phosphorus release by submerged macrophytes: Significance to epiphyton and phytoplankton1,1. Limnol. Oceanogr., 27: 419-427.
CrossRefDirect Link - Howard-Williams, C. and B.R. Allanson, 1981. Phosphorus cycling in a dense Potamogeton pectinatus L. Bed. Oecologia, 49: 56-66.
CrossRefDirect Link - Ezzat, S.M., H.M., Mahdy, M.A. Abo-State, E.H. Abd El-Shakour and M.A. El-Bahnasawy, 2012. Water quality assessment of river Nile at Rosetta branch: Impact of drains discharge. Middle-East J. Scient. Res., 12: 413-423.
Direct Link - WHO., 2006. Guidelines for Drinking-Water Quality. 3rd Edn., World Health Organization, Geneva.
Direct Link - Liboriussen, L. and E. Jeppesen, 2003. Temporal dynamics in epipelic, pelagic and epiphytic algal production in a clear and a turbid shallow lake. Freshwater Biol., 48: 418-431.
CrossRefDirect Link - Jeppesen, E., J.P. Jensen, M. Søndergaard, T. Lauridsen and R. Landkildehus, 2000. Trophic structure, species richness and biodiversity in Danish lakes: Changes along a phosphorus gradient. Freshwater Ecol., 4: 201-218.
CrossRefDirect Link - Sand‐Jensen, K., T. Riis, O. Vestergaard and S.E. Larsen, 2000. Macrophyte decline in Danish lakes and streams over the past 100 years. J. Ecol., 88: 1030-1040.
CrossRefDirect Link - Uedeme-Naa, B., U.U. Gabriel and O.A. Akinrotimi, 2011. The relationship between aquatic macrophytes and water quality in Nta-Wogba Stream, Port Harcourt, Nigeria. Continental J. Fish. Aquat. Sci., 5: 6-16.
Direct Link - Li, Q., 2012. Influence of silts on the growth, reproduction and chlorophyll fluorescence of Potamogeton crispus in turbid water. Russ. J. Ecol., 43: 122-130.
CrossRefDirect Link - Frankovich, T.A., E.E. Gaiser, J.C. Zieman and A.H. Wachnicka, 2006. Spatial and temporal distributions of epiphytic diatoms growing on Thalassia testudinum Banks ex König: Relationships to water quality. Hydrobiologia, 569: 259-271.
CrossRefDirect Link - Barbour, M.T., J. Gerritsen, B.D. Snyder and J.B. Stribling, 1999. Rapid Bioassessment Protocols for Use in Streams and Wadeable Rivers: Periphyton, Benthic Macroinvertebrates and Fish. 2nd Edn., U.S. Environmental Protection Agency, Office of Water, Washington, D.C.
Direct Link - Ali, E.M. and A. El Shehawy, 2017. Environmental indices and phytoplankton community structure as biological indicators for water quality of the River Nile, Egypt. Egypt. J. Aquat. Biol. Fish., 21: 87-104.
CrossRefDirect Link - Toporowska, M., B. Pawlik-Skowrońska and A. Wojtal, 2008. Epiphytic algae on Stratiotes aloides L., Potamogeton lucens L., Ceratophyllum demersum L. and Chara spp. in a macrophyte-dominated lake. Oceanol. Hydrobiol. Stud., 37: 51-63.
CrossRefDirect Link - Ali, M.M. and M.E. Soltan, 1996. The impact of three industrial effluents on submerged aquatic plants in the River Nile, Egypt. Hydrobiologia, 340: 77-83.
CrossRefDirect Link