
Albertus Fajar Irawan
Department of Research and Development, PT Sampoerna Agro Tbk, Jl. Basuki Rahmat No. 788, Palembang, 30127 South Sumatra, Indonesia
LiveDNA: 62.31180
Gregorius Baskara
Department of Research and Development, PT Sampoerna Agro Tbk, Jl. Basuki Rahmat No. 788, Palembang, 30127 South Sumatra, Indonesia
Ruli Wandri
Department of Research and Development, PT Sampoerna Agro Tbk, Jl. Basuki Rahmat No. 788, Palembang, 30127 South Sumatra, Indonesia
Dwi Asmono
Department of Research and Development, PT Sampoerna Agro Tbk, Jl. Basuki Rahmat No. 788, Palembang, 30127 South Sumatra, Indonesia
Pakistan Journal of Biological Sciences
Year: 2020 | Volume: 23 | Issue: 5 | Page No.: 667-673







ABSTRACT
Background and Objective: Phosphate-solubilising bacteria (PSB) are useful for plant growth. They inhabit different soil ecosystems such as colonizing the root environment. The aims of this study was to isolate PSB from oil palm rhizosphere and to conduct a comparative analysis of the solubility of inorganic phosphates. Materials and Methods: Rhizospheric soil samples at 0-20 cm depth collected from the distance of 2 m away from the palm were isolated and their chemical and physical properties were analyzed. Qualitative estimation of the suspected PSB was screened by inoculating and growing them at 27°C for 10 days on NBRIP agar medium with bromophenol blue. Their abilities to solubilize AlPO4, FePO4 and Ca3(PO4)2 were examined. Phosphate solubilizing activities were tested on the NBRIP growth medium by analyzing solubilisation efficiency and soluble-P content. Genomic DNA was isolated using QIAamp® genomic DNA kit. Results: A total of 15 PSB were successfully isolated from oil palm rhizosphere. During 5 days of incubation, isolate K3.1, A4 and K3.3 solubilized 53.5, 63.5 and 58.6 mg L–1 phosphate inoculated in Al3PO4, Fe3PO4 and Ca3(PO4)2, respectively. Based on the 16S rRNA gene sequence analysis, those isolates were closely related to Burkholderia arboris, Burkholderia gladioli and Burkholderia seminalis, respectively. In soil analysis, P2O5, C-organic and CEC had positive correlation with the total PSB. Conclusion: The existence of P promoting bacteria in oil palm rhizosphere may offer effective solution on biofertilizer agent for sustainable agriculture.
PDF Abstract XML References Citation
Copyright: © 2020. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Albertus Fajar Irawan, Gregorius Baskara, Ruli Wandri and Dwi Asmono, 2020. Isolation and Solubilisation of Inorganic Phosphate by Burkholderia spp. from the Rhizosphere of Oil Palm. Pakistan Journal of Biological Sciences, 23: 667-673.
DOI: 10.3923/pjbs.2020.667.673
URL: https://scialert.net/abstract/?doi=pjbs.2020.667.673
DOI: 10.3923/pjbs.2020.667.673
URL: https://scialert.net/abstract/?doi=pjbs.2020.667.673
INTRODUCTION
Phosphate (P) is the main macronutrient compound which is essential for plant growth. The maximum part of soil phosphate, approximately 95-99% is present in the form of insoluble phosphates and hence it cannot be easily utilized by the plants1. Depending on some environmental and biological factors, it can be the main growth-limiting nutrient2.
Phosphate is most readily available3,4 at pH around 6-7. The insoluble forms of P such as tricalcium phosphate (Ca3(PO4)2), aluminum phosphate (AlPO4) and iron phosphate (FePO4) may be converted to soluble P by phosphate-solubilising bacteria (PSB) inhabiting different soil ecosystems5,6,7. The role of bacteria in dissolving P is more widely used because bacteria are also able to produce other acids such as amino acids, vitamins and growth-promoting hormones such as gibberellins and IAA which are useful for plant growth5. PSB can play an important role in dissolving both of fertilizer P and bound P in the soil that is environmentally friendly and sustainable8. Moreover, according to Behera et al.9, PSB in solubilizing P derived from soil or fertilizing activities in several agricultural fields depend much on their abilities to secrete organic acids such as citric acid, formic acid, oxalic acid, lactic acid, acetic acid and acid malat.
Despite the agronomic benefits that may be provided by PSB, their abundance in soil is not always sufficient to compete with other microorganisms established in the rhizosphere10. In order to establish sustainable oil palm plantation especially on marginal soils and those with less dependent on inorganic fertilizers, it needs to identify and search the important soil bacteria which have high influence on plant growth, such as PSB11. The aims of this study were to isolate PSB from oil palm rhizosphere and to conduct a comparative analysis of the solubility inorganic phosphates from identified PSB.
MATERIALS AND METHODS
Time and venue: The study was performed in oil palm cultivated area belonged to PT Sampoerna Agro Tbk. located in Ogan Komering Ilir, South Sumatra Province, Indonesia (3°47’ 04.1” S to 3°47’12.8”S and 105°10’26.6”E to 105°10’ 59.0”E). It lasted from May to October, 2016. The palm was at productive stage and aged about 10 years old after planting. The soil was classified as acidic soil.
Soil sampling and bacterial isolation: Rhizospheric soils at 0-20 cm depth were collected from 3 different points at the distance of 2 m from each palm. A total of 6 palms were used in this study. Those collected soil samples were then mixed and composited into one samples with total fresh weight of 3 kg. About 1.5 kg of soil was used for the analysis of chemical (soil pH, N-total, C-organic, CEC, Cation exchange of K, Ca and Mg, P-total, available P and Exch. Al and H) and physical (composition of silt, clay and sand) properties. The rest was used for bacterial isolation.
Ten gram of fresh rhizospheric soil samples were transferred into 250 mL Erlenmeyer flasks containing sterilized 90 mL of 0.85% NaCl solution. A series of ten-fold dilutions of the suspension was made by pipetting 1 mL aliquots into sterilized 9 mL of 0.85% NaCl solution. Aliquots of 0.1 mL of the sample from each of these dilutions were spread on to a petri dish with National Botanical Research Institute Phosphorus (NBRIP) agar medium containing 10 g of glucose, 3 g of AlPO4 or 5 g of FePO4, 5 g of MgCl2.6H2O, 0.25 g of MgSO4.7H2O, 0.2 g of KCl, 0.1 g of (NH4)2SO4 and 10 g of gellan gum agar in 1 L distilled water12. The pH of the media was adjusted to 5.0 using HCl. The plates were incubated for 7 days in an incubator at 30°C. The colonies with halo zone were considered to be PSB. These bacteria were further purified by re-streaking on the fresh NBRIP agar plates at 30°C.
Qualitative and quantitative estimation of phosphate solubilisation: An efficient protocol was developed for qualitative screening PSB, based upon visual observation. Qualitative estimation of all the suspected PSB was screened by inoculating and growing them at 27°C for 10 days on NBRIP agar medium with bromophenol blue. The diameter of the halo zone was measured after 42 h, up to 10 days. The phosphate solubilisation efficiency (PSE) was identified by measuring the total halo zone of the colony and the colony diameter13. The calculation was as follow:
The quantitative bioassay was carried out using erlenmeyer flasks (250 mL) containing 100 mL NBRIP agar media inoculated using bacterial isolates with approximately 1×108-109 Colony Forming Units (CFU) per mL following Saraswati and Sumarno's method14. The bacteria were inoculated in the medium having pH 5.0, which was adjusted before autoclaving the medium. The flasks were incubated at 30°C in a shaker for 5 days at 100 rpm. The cultures were collected for centrifugation for 10 min at 5,500 rpm. The supernatant was decanted and filtered through Whatman No. 41 filter paper. The available P content in the supernatant was estimated by the phosphomolybdate blue complex colorimetric method at 660 nm wavelength15. From this method, the abilities of selected isolates to solubilize AlPO4, FePO4 and Ca3(PO4)2 were examined.
DNA extraction, PCR amplification and sequencing of 16S rRNA gene: Genomic DNA was isolated using QIAamp® genomic DNA kit following the manufacturer’s instructions and 16S rRNA gene was amplified using the universal primers, 27f (5’AGAGTTTGATCCTGGCTCAG 3’) and 1492r (5' TACGGCTACCTTGTTACGACTT 3'). The PCR thermal cycling conditions consisted of an initial denaturation step at 94°C for 3 min, followed by 30 cycles of denaturation (1 min at 94°C), annealing for 1 min at 57°C and extension for 2 min at 72°C, followed by a final extension at 72°C for 8 min.
PCR amplified products were separated on 1.0% agarose gels in 1×TBE buffer at 70 V cm–1 for 20 min. Partial 16S rRNA genes of selected isolates in each group were sequenced by FIRSTBASE, Singapore and compared to that of other bacteria by way BLAST (http://www.ncbi.nlm.nih.gov/BLAST/Blast.cgi). In the best isolate(s) (phosphate-solubilizing ability) and three isolates were selected to sequence and the results were compared to sequences of GenBank based on partial 16S rRNA sequences to show relationships between PSB strains and a phylogenetic tree was constructed by the maximum-likelihood method using the MEGA software ver. 6.06 based on 1,000 bootstraps.
Statistical analysis: The in vitro experiment was conducted in complete randomized design with 3 replications and data were expressed as the mean value±standard error (SE). The treatments of identified isolates and inorganic phosphate solubilisation were used. The treatment of inorganic phosphates solubilisation consisted of AlPO4, Ca3(PO4)2 and FePO4. All data were analyzed using the Predictive Analytics Software (PSAW)’s statistical software and mean differences were separated using Duncan’s multiple range test at the 5% level of probability.
RESULTS AND DISCUSSION
Soil analysis: The soil collected from this study contained majority 75.27% sand, minor proportion of silt (4.21%) and the clay was 20.53% (Table 1).
| Table 1: | Soil characteristics collected from the rhizosphere (0-20 cm layer) of oil palm |
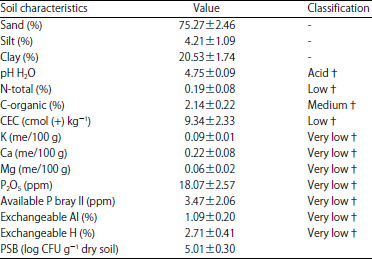 | |
| †: Classification of the ranges based on Hardjowigeno16 | |
According to Hardjowigeno16, it was classified as acid with soil pH at 4.75. It contained low N, medium C-organic, low CEC and very low exchangeable K, Ca and Mg as well as available P and total P. The exchangeable Al and H were very low.
According to Sanchez17 and Rao et al.18, the natural acidification process in acidic soils had caused Al toxicity, deficiencies in Ca, Mg and Mo and a frequently low availability of P. Moreover, they found that in acid mineral soils, K and N tend to be deficient because they are usually highly weathered or have low organic matter content. The severe limitation of inorganic phosphate in acid soils was also reported by Zheng et al.4. Most of the soil analysis results reported in this study had agreed with those findings.
Total PSB reported in the present study was 5.01 or 2.24×105 CFU g–1 of soil. It was higher than those reported by several studies in acidic soil in Kenya, Uruguay and Argentina. Ndung'u-Magiroi et al.19 reported the amount of PSB in 13 different sites of Kenya ranging from 0.38 to 9.1×105 CFU g–1 of soil. Similarly, Azziz et al.2 reported that PSB present under crop-pasture rotations in Uruguay varied from 0.65 to 62×105 CFU g–1 of soil. In contrast, Fernandez et al.20 recorded a low amount of PSB, located between 0.03 and 0.08×105 CFU g–1 of soil, in the most productive region of the Argentina.
Phosphate solubilisation efficiency (PSE): The isolated bacterial strains were capable of solubilizing Ca3(PO4)2 in NBRIP medium and they formed large halos with varied intensity (Fig. 1) and the halo-zone increased with increase of colony diameter. Phosphate levels during 10 days of incubation showed high phosphate solubilization activity. These findings were similar as reported by Li et al.21.
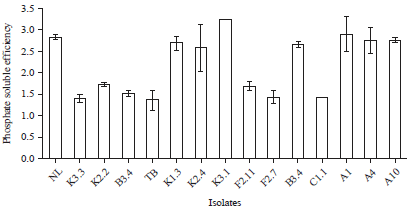 | |
| Fig. 1: | Phosphate soluble efficiency index after 10 days of incubation period |
| Table 2: | Iron, aluminum and calcium phosphates solubilisation by the selected isolates |
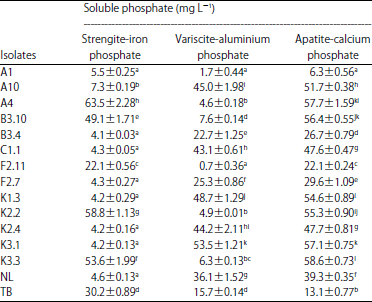 | |
Values are given as Mean±SD for triplicate samples, means followed by the same letter (s) in each column are not significantly different at p<0.05 by Duncan’s multiple ranged test | |
Among the isolates, the highest solubilisation index was found on isolate K3.1 (3.2) followed by isolate A1 (2.9). The incubation period required for exhibiting the solubilisation index for both isolates was 5 days. For isolates A4 and A10, the solubilisation index was recorded at 8 days of the incubation. Lower solubilisation index was found on K2.4 (2.6) at the same incubation period. These solubilisation activities reported in the present study were similar with those in costal soils with the PSE value13 of 2.63. In addition, Ktt.B2 isolated from tropical acidic soil solubilized phosphate22 at 3.47.
Inorganic phosphate solubilisation: Among 15 collected isolates shown in Table 2, isolates A4, K3.1 and K3.3 were the most efficient in solubilizing FePO4 at 63.5 mg L–1, AlPO4 at 53.5 mg L–1 and Ca3(PO4)2 at 58.6 mg L–1, respectively. At the most cases, all isolates were able to solubilize Ca3(PO4)2 to a greater extent than AlPO4 and FePO4. Isolate K3.1 was able to solubilize AlPO4 at 53.5 mg L–1 which was five times greater than isolate N1 from oxisol (9.32 mg L–1) as reported by Prijambada et al.23. Moreover, the solubilisation of iron phosphate by isolate A.4 was similar to isolate M150 (61.3 mg L–1) reported by Panda et al.6 in the rhizosphere of maize.. The huge variation in iron, aluminum and calcium phosphates solubilisation by the selected isolates might have indicated that the use of microbial phosphate solvent still faced several obstacles such as soil factor, since every soil type possessed different phosphate form.
Correlation between total PSB and soil physicochemical properties: The present study found that the total PSB was greatly affected by several soil physicochemical properties (Table 3). Those were with C-organic (r = 0.94), P2O5 (r = 0.85) and CEC (r = 0.84). Other physicochemical properties such as soil physics, pH, available P and macro- and semi macronutrients were not correlated. These findings was very similar to those reported by Vikram et al.24 who found strong positive correlation between organic carbon (r = 0.40, p<0.01), available N (r = 0.40, p<0.05) and PSB population. However, pH and available P showed no significant correlation. A study performed by Mahanta et al.25 reported that CEC was in strong correlation with P inflow rate and the requirement of internal P influenced PSB activity. The present study showed that available P was positively correlated with CEC (r = 0.72).
The unclear correlation of other chemical properties with the total PSB might have been explained that the abundance of PSB in the soil depended on plant species, soil microbial composition and soil condition26. Nahas27 reported that diversity and density of the population of PSB and their bio-activity varied from soil to another according to their nutritional status [C, N and P] and their own effectiveness in phosphate solubilization. Next to N and K, phosphorus was an essential element of energy metabolism of all life forms.
| Table 3: | Pearson correlation matrix among P solubilizing bacteria (PSB), soil physics and soil chemistry in the acidic soil (0-20cm layer) of oil palm rhizosphere |
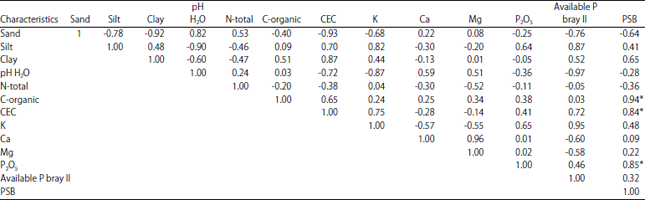 | |
| *Correlation is significant at the 0.05 probability level (2-tailed) | |
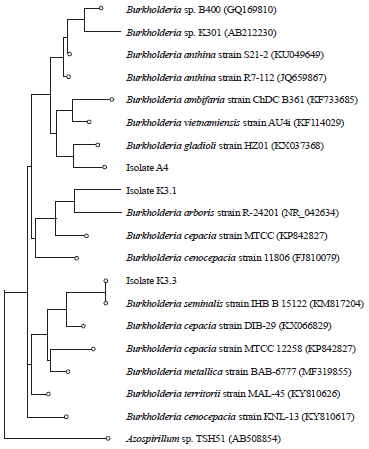 | |
| Fig. 2: | Phylogenetic tree showing the relationships between the PSB from oil palm rhizosphere in this study and their closest phylogenetic relatives based on 16S rRNA gene sequencing |
Molecular identification: Based on the 16S rRNA sequence analysis, all the strains were identified in the genus of Burkholderia spp. (Fig. 2). Isolates A.4, K3.1 and K3.3 were identified as Burkholderia gladioli, Burkholderia arboris and Burkholderia seminalis, respectively. Previous reports also described that some Burkholderia spp. were efficient as phosphate solubilizers by producing organic acids28,29. Burkholderia was known as N fixation bacteria and has the antifungal ability30,31. Moreover, it was efficient bacteria used as a biological fertilizer on agricultural land32,33.
CONCLUSION
In the present study, rhizosphere soil samples from oil palm were screened for the isolation of PSB. Based on the sequencing results, isolates K3.1, A4 and K3.3 were closely related to Burkholderia arboris, Burkholderia gladioli and Burkholderia seminalis, respectively.
SIGNIFICANCE STATEMENT
These results discover the existence of the important PSB living in the rhizosphere of oil palm growing in acid soil. It may help the researchers do the possible studies on utilization of these potential microbes in order to improve soil fertility, particularly for the phosphorus content in acid soil. Thus, a new theory on the existence of PSB in relation to soil-phosphorous source and possibly future application as soil-applied biofertilizer, may be arrived at.
ACKNOWLEDGMENT
This work was fully supported by the Department of Research and Development, PT. Sampoerna Agro Tbk.
REFERENCES
- Sundareshwar, P.V., J.T. Morris, E.K. Koepfler and B. Fornwalt, 2003. Phosphorus limitation of coastal ecosystem processes. Science, 299: 563-565.
CrossRefDirect Link - Azziz, G., N. Bajsa, T. Haghjou, C. Taule, A. Valverde, J.M. Igual and A. Arias, 2012. Abundance, diversity and prospecting of culturable phosphate solubilizing bacteria on soils under crop-pasture rotations in a no-tillage regime in Uruguay. Applied Soil Ecol., 61: 320-326.
CrossRefDirect Link - Curtin, D. and J.K. Syers, 2001. Lime-induced changes in indices of soil phosphate availability. Soil Sci. Soc. Am. J., 65: 147-152.
CrossRefDirect Link - Zheng, B.X., D.P. Zhang, Y. Wang, X.L. Hao and M.A. Wadaan et al., 2019. Responses to soil pH gradients of inorganic phosphate solubilizing bacteria community. Scient. Rep., Vol. 9.
CrossRefDirect Link - Khan, A.A., G. Jilani, M.S. Akhtar, S.M.S. Naqvi and M. Rasheed, 2009. Phosphorus solubilizing bacteria: Occurrence, mechanisms and their role in crop production. J. Agric. Biol. Sci., 1: 48-58.
Direct Link - Panda, B., H. Rahman and J. Panda, 2016. Phosphate solubilizing bacteria from the acidic soils of Eastern Himalayan region and their antagonistic effect on fungal pathogens. Rhizosphere, 2: 62-71.
CrossRefDirect Link - Song, O.R., S.J. Lee, Y.S. Lee, S.C. Lee, K.K. Kim and Y.L. Choi, 2008. Solubilization of insoluble inorganic phosphate by Burkholderia cepacia DA23 isolated from cultivated soil. Braz. J. Microbiol., 39: 151-156.
CrossRefDirect Link - Khan, M.S., A. Zaidi and P.A. Wani, 2007. Role of phosphate-solubilizing microorganisms in sustainable agriculture-A review. Agron. Sustain. Dev., 27: 29-43.
CrossRefDirect Link - Behera, B.C., H. Yadav, S.K. Singh, R.R. Mishra, B.K. Sethi, S.K. Dutta and H.N. Thatoi, 2017. Phosphate solubilization and acid phosphatase activity of Serratia sp. isolated from mangrove soil of Mahanadi river delta, Odisha, India. J. Genet. Eng. Biotechnol., 15: 169-178.
CrossRefDirect Link - Igual, J.M., A. Valverde, E. Cervantes and E. Velazquez, 2001. Phosphate-solubilizing bacteria as inoculants for agriculture: Use of updated molecular techniques in their study. Agronomie, 21: 561-568.
CrossRefDirect Link - Sharma, S.B., R.Z. Sayyed, M.H. Trivedi and T.A. Gobi, 2013. Phosphate solubilizing microbes: Sustainable approach for managing phosphorus deficiency in agricultural soils. SpringerPlus, Vol. 2.
CrossRefDirect Link - Nautiyal, C.S., 1999. An efficient microbiological growth medium for screening phosphate solubilizing microorganisms. FEMS Microbiol. Lett., 170: 265-270.
CrossRefPubMedDirect Link - Sanjotha, G. and S. Manawadi, 2016. Isolation, screening and characterization of phosphate solubilizing bacteria from Karwar costal region. Int. J. Res. Stud. Microbiol. Biotechnol., 2: 1-6.
CrossRefDirect Link - Saraswati, R. and Sumarno, 2008. [Utilization of soil decomposer as an agent of agriculture technology]. Iptek Tanaman Pangan, 3: 41-58, (In Indonesian).
Direct Link - Adnan, M., Z. Shah, S. Fahad, M. Arif and M. Alam et al., 2017. Phosphate-solubilizing bacteria nullify the antagonistic effect of soil calcification on bioavailability of phosphorus in alkaline soils. Scient. Rep., Vol. 7.
CrossRefDirect Link - Sanchez, P.A., 1976. Properties and Management of Soils in the Tropics. John Wiley and Sons, New York, ISBN: 9780471752004, 618.
Direct Link - Rao, I.M., R.S. Zeigler, R. Vera and S. Sarkarung, 1993. Selection and breeding for acid-soil tolerance in crops: Upland rice and tropical forages as case studies. BioScience, 43: 454-465.
CrossRefDirect Link - Ndung'u-Magiroi, K.W., L. Herrmann, J.R. Okalebo, C.O. Othieno, P. Pypers and D. Lesueur, 2012. Occurrence and genetic diversity of phosphate-solubilizing bacteria in soils of differing chemical characteristics in Kenya. Ann. Microbiol., 62: 897-904.
CrossRefDirect Link - Fernandez, L.A., B. Agaras, L.G. Wall and C. Valverde, 2015. Abundance and ribotypes of phosphate-solubilizing bacteria in Argentinean agricultural soils under no-till management. Ann. Microbiol., 65: 1667-1678.
CrossRefDirect Link - Li, Y., X. Liu, T. Hao and S. Chen, 2017. Colonization and maize growth promotion induced by phosphate solubilizing bacterial isolates. Int. J. Mol. Sci., Vol. 18, No. 7.
CrossRefDirect Link - Tamad, B. Radjagukguk, E. Hanudin and J. Widada, 2011. [Selection of phosphate-solublizing bacteria isolates for developing effective innoculum]. Biosfer, 29: 93-101, (In Indonesian).
Direct Link - Prijambada, I.D., J. Widada, S. Kabirun and D. Widianto, 2009. Secretion of organic acids by phosphate solubilizing bacteria isolated from oxisols. J. Tanah Trop., 14: 245-251.
Direct Link - Vikram, A., A.R. Alagawadi, H. Hamzehzarghani and P.U. Krishnaraj, 2007. Factors related to the occurrence of phosphate solubilizing bacteria and their isolation in vertisols. Int. J. Agric. Res., 2: 571-580.
CrossRefDirect Link - Mahanta, D., R.K. Rai, S.D. Mishra, A. Raja, T.J. Purakayastha and E. Varghese, 2014. Influence of phosphorus and biofertilizers on soybean and wheat root growth and properties. Field Crops Res., 166: 1-9.
CrossRefDirect Link - Katiyar, V. and R. Goel, 2003. Solubilization of inorganic phosphate and plant growth promotion by cold tolerant mutants of Pseudomonas fluorescens. Microbiol. Res., 158: 163-168.
CrossRefDirect Link - Nahas, E., 2007. Phosphate Solubilizing Microorganisms: Effect of Carbon, Nitrogen and Phosphorus Sources. In: First International Meeting on Microbial Phosphate Solubilization (Developments in Plant and Soil Sciences Series, Volume 102), Velazquez, E. and C. Rodriguez-Barrueco (Eds.). Springer, Dordrecht, The Netherlands, ISBN-13: 9781402057656, pp: 111-115.
Direct Link - Caballero-Mellado, J., J. Onofre-Lemus, P. Estrada-de los Santos and L. Martinez-Aguilar, 2007. The tomato rhizosphere, an environment rich in nitrogen-fixing Burkholderia species with capabilities of interest for agriculture and bioremediation. Applied Environ. Microbiol., 73: 5308-5319.
CrossRefPubMedDirect Link - Istina, I.N., H. Widiastuti, B. Joy and M. Antralina, 2015. Phosphate-solubilizing microbe from saprists peat soil and their potency to enhance oil palm growth and P uptake. Procedia Food Sci., 3: 426-435.
CrossRefDirect Link - Stephen, J. and M.S. Jisha, 2011. Gluconic acid production as the principal mechanism of mineral phosphate solubilization by Burkholderia sp. (MTCC 8369). J. Trop. Agric., 49: 99-103.
Direct Link - Wahdi, E., N.R. Mubarik and R. Widyastuti, 2016. Characterization of phosphate solubilising bacteria from limestone quarry in Cirebon Indonesia. J. Int. Environ. Applic. Sci., 11: 312-317.
Direct Link - Lavania, M. and C.S. Nautiyal, 2013. Solubilization of tricalcium phosphate by temperature and salt tolerant Serratia marcescens NBRI1213 isolated from alkaline soils. Afr. J. Microbiol. Res., 7: 4403-4413.
Direct Link - Rodriguez, H. and R. Fraga, 1999. Phosphate solubilizing bacteria and their role in plant growth promotion. Biotechnol. Adv., 17: 319-339.
CrossRefDirect Link