
Eman M. Marei
Department of Agricultural Microbiology, Virology Laboratory, Faculty of Agriculture, Ain Shams University, P.O. Box 68, 11241 Hadayek-Shoubra, Cairo, Egypt
LiveDNA: 20.23985
Pakistan Journal of Biological Sciences
Year: 2020 | Volume: 23 | Issue: 4 | Page No.: 491-500







ABSTRACT
Background and Objective: Pseudomonas aeruginosa is a free living bacterium in widely different areas such as plants, soil, water and other moist locations. It is pathogenic to plants and humans. P. aeruginosa causes several disease symptoms to plants such as wet rot and curved leaves. The virulent bacterial viruses of P. aeruginosa were found to be of widespread occurrence in nature and isolated from widely different sources. Bacterial viruses were applied to control pathogenic bacteria in different fields and successfully. Therefore, this work aimed to study the different characteristics of P. aeruginosa lytic phage isolates. Moreover, the bio-control of P. aeruginosa by lytic phage isolates was also studied. Material and Methods: Different physical and molecular characteristics were assayed and determined of P. aeruginosa lytic bacteriophages. Also, the effect of phage isolates on P. aeruginosa as a bio-control under lab condition was studied. Results: Pseudomonas aeruginosa pathogenic bacterium was isolated from a sewage water sample. Two lytic bacteriophages specific to P. aeruginosa were isolated from same sewage water sample and designated Pa1 and Pa2. Both phage isolates (Pa1 and Pa2) found to be stable in 90°C and different pH low and high levels. The total count of P. aeruginosa decreased after 48 h in broth treated with lytic phages. RAPD-PCR amplification was indicated that the two phage isolates (Pa1 and Pa2) are belonging to two different phage types. Conclusion: The results of this study indicated that both lytic phage isolates could be used as a biological control agents against the plant pathogen P. aeuroginosa.
PDF Abstract XML References Citation
Copyright: © 2020. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Eman M. Marei, 2020. Isolation and Characterization of Pseudomonas aeruginosa and its Virulent Bacteriophages. Pakistan Journal of Biological Sciences, 23: 491-500.
DOI: 10.3923/pjbs.2020.491.500
URL: https://scialert.net/abstract/?doi=pjbs.2020.491.500
DOI: 10.3923/pjbs.2020.491.500
URL: https://scialert.net/abstract/?doi=pjbs.2020.491.500
INTRODUCTION
Pseudomonas aeruginosa is a free living bacterium in widely different areas such as plants, soil, water and other moist locations. It is pathogenic to plants and humans1,2. Moreover, it is stated that Pseudomonas aeruginosa is widely distributed in nature, it has been found in external environmental conditions surrounding dairy animals such as air, sewage, soil, grass3,4.
Pseudomonas aeruginosa causes several disease symptoms to plants such as wet rot and curved leaves. A bacterial leaf spot disease of tobacco caused by P. aeruginosa was detected in Philippines5 and China6.
Also, in California, 2 types of P. aeruginosa were found in other plants, type B-7 was found in tomato leaves and type S was found in a celery plant7.
The virulent bacterial viruses of P. aeruginosa were found to be of widespread occurrence in nature and isolated from widely different sources such as sewage water samples4,8,9 as well as soil and vegetable materials10.
Bacterial viruses were applied to control pathogenic bacteria in foodstuffs, industrial environments and successfully used as phage therapy against some animal diseases8,11-13. Moreover, in United States bacteriophages were applied to certain meat products since August, 2006 to control Listeria monocytogenes14.
Random amplified polymorphic DNA (RAPD) technique is a powerful approach and accessible genotyping method can be used to classify P. aeruginosa isolates15-17.
On the basis of the above mentioned information, the lytic phages of P. aeruginosa could be used to control the plant diseases caused by this bacterium.
Therefore, this study aimed to study the physical and molecular characteristics of lytic bacteriophages specific to P. aeruginosa. Also, study the effect of phage isolates on P. aeruginosa as a biocontrol under lab condition.
MATERIALS AND METHODS
The study was carried out at Virology Laboratory, Department of Agric. Microbiology, Faculty of Agriculture, Ain Shams University, Cairo, Egypt (from April, 2018-March, 2019).
Source of bacteria: Pseudomonas aeruginosa was isolated from a sewage water sample collected after primary treatment from Sarpium forest site -Ismailia Governorate, Cairo, Egypt.
Serial dilutions of the sewage water were prepared as described by Goto and Enomoto18. Plates containing MacConkey agar medium were inoculated with the prepared dilutions (10–4 and 10–5) and incubated at 37°C for 24-48 h. A single colony was transferred onto slant surface of MacConkey agar and incubated at 37°C for 48 h. The bacterial isolate was identified in Sohag University, Egypt, using Biolog19 and was found to be Pseudomonas aeruginosa.
Detection of prophage in the isolated P. aeruginosa: Pseudomonas aeruginosa was tested for lysogeny using U.V. irradiation (240 nm) as described by Prinsloo20. To induce temperate phages, liquid culture of P. aeruginosa (24 h old) was prepared in nutrient broth medium21.
Ten milliliter of the prepared liquid culture were placed in a petri plate and exposed to UV irradiation (240 nm) at distance of 60 cm from the germicidal UV lamp. One milliliter of this culture was withdrawn every 5 up to 30 min and kept in Eppendorf tube. After overnight incubation at 37°C, bacterial cells were precipitated by centrifugation at 5000 rpm and the temperate phage was qualitatively assayed in the supernatant using the spot test22.
Isolation and enrichment of P. aeruginosa lytic phages: The liquid enrichment technique22 was carried out to isolate virulent phages specific to P. aeruginosa from the collected sewage water sample. Nutrient broth medium21 was used to grow the bacterial host (P. aeruginosa).
Detection of phages: Bacteriophages were detected using the spot test as described by Adams22.
Bacteriophages purification: The single plaque isolation (SPI) technique was used as described by Kiraly et al.23 to obtain pure single phage isolates.
High titer phage suspension: High titer suspensions of P. aeruginosa bacteriophages were prepared using the nutrient broth medium for enrichment technique as described by Sambrook et al.24. Titers of the prepared phage suspensions were estimated using the method described by Kiraly et al.23.
Physical properties of phages:
| • | Different pH level for phage infection: The optimum pH level for infection of P. aeruginosa with each of the isolated bacteriophages was estimated as described by Hammad et al.25 |
| • | Stability of phage to UV irradiation: The stability of phages to UV irradiation with different periods for infection of P. aeruginosa was estimated as described by Hammad et al.25 |
| • | Thermal inactivation point: The thermal inactivation point of each isolated bacterial virus of P. aeruginosa was estimated according to Hammad et al.25 |
| • | Electron microscopy: Size and morphology of the purified particles using Backman L 7-35 ultracentrifuge at 30.000 rpm for 1.5 h at 4°C of virulent phage isolates were estimated as described by Hayat and Miller26 and Stacey et al.27. Phosphotungstic acid (pH 6.8) was used for negative staining of the isolated phage particles. The stained Phage particles were examined using transmission electron microscopy (Joel, Model GEM 1010) in Sohag University, Sohag |
Effect of phages on P. aeruginosa as a biocontrol: Under lab condition experiment was carried out to study the effect of presence of Pa1 and Pa2 phages on the efficiency of Pseudomonas aeruginosa as a biocontrol. Twelve flasks (250 mL) were filled with 100 mL broth media for each one and sterilized. The flasks were divided into 4 groups, each group comprised 3 flasks. The groups were subjected to the following treatments:
| • | First group were inoculated with 5 mL of liquid culture (33×106 CFU mL–1) P. aeruginosa and 5 mL of Pa1 lytic phage (43×108 PFU mL–1) |
| • | Second group was inoculated with 5 mL of liquid culture (33×106 CFU mL–1) P. aeruginosa and 5 mL of Pa2 lytic phages (36×108 PFU mL–1) |
| • | Third group was inoculated with 5 mL of P. aeruginosa and 5 mL of Pa1+5 mL of Pa2 lytic phages |
| • | Fourth group was inoculation with 5 mL of liquid culture (33×106 CFU mL–1) P. aeruginosa only |
Total count of Pseudomonas aeruginosa was estimated of each group after 48 h. from inoculation with each treatment by using the dilution plate technique according to Clark et al.28.
Molecular characterization:
| • | Extraction of bacteriophages DNA: Genomic DNA of each bacteriophage isolate specific to P. aeruginosa was extracted as described by Maniatis et al.29 |
| • | Determination of DNA concentration: The quality of the DNA extract was estimated by 0.8% agarose gel electrophoresis using 1X TBE (Tris-Borate-EDTA) buffer. Ethidium bromide was used to stain Gels. The genomic DNA concentration was estimated by measuring (UV-1601 UV/VIS Japan spectrophotometer) the optical density at 260 nm. The DNA quality was estimated using the 260/280 nm ratio and gel electrophoresis |
| • | Digestion of bacteriophages DNA with HindIII restriction enzyme: The reactions were assembled by mixing the following components in a sterile 0.5 mL Eppendorf vial: 1 μg bacteriophage DNA, 3 units of HindIII restriction enzyme (enzynomics), 4 μL enzyme buffer and sterile deionized water to a final volume of 20 μL |
The reaction mixtures were incubated for 2-3 h at 37°C. Five μL of bromophenol blue dye stop [450 mM tris borate pH 8.3, 50 mM EDTA, 50% (v/v) glycerol, 0.2% (w/v) bromophenol blue] were added and the reactions were heated at 60°C for 5 min to stop the reactions. The digested DNA was stored at 4°C and analyzed in 1% agarose gel using 1X TBE (Tris-Borate-EDTA) buffer for 1.5 h at 90 V as described by Peacock and Dingman30.
| • | PCR reaction and amplification condition: PCR amplification was carried out using two random 10 mer arbitrary primers, OPH-01 and OPH-02 were supplied by Bio Basic Inc, Canada. Whereas, 3 SCoT primers, SCoT-7, SCoT-8 and SCoT-9 were obtained from iNtRON Biotechnology, Inc, Korea with GC content of 60 % were also used. Sequences of the used RAPD and SCoT primers were presented in Table 1 |
The PCR amplification was conducted according to Williams et al.31 using 2X PCR Master mix solution [(i-TaqTM) iNtRON Biotechnology] in 10 μL reaction volume contained 1 μL (40 ng) genomic DNA as a template, 5 μL of 2X PCR Master mix solution [(i-TaqTM) iNtRON Biotechnology], 1 μL of primer (10 pmol μL–1) and 3 μL double distilled water. The reaction mixtures were over-laid with 20 μL of mineral oil/sample.
| Table 1: | Names and nucleotide sequence of the used primers |
 | |
The PCR amplification was carried out in a DNA Thermal Cycler (Cetus, Perkin Elmer) programmed for a first denaturation step of 5 min at 94°C followed by 40 cycles of 1 min at 94°C, 1 min annealing temperature at 30°C for RAPD primers and at 50°C for SCoT primers, 1.5 min for extension at 72°C, followed by a final 7-min elongation step at 72°C and then held at 4°C until the tubes were removed.
The amplification products were separated by a horizontal gel electrophoresis unit using 1.5 % agarose gel for RAPD and SCoT markers. Gels were then run for 1.5 h at current 90 V. Bands were documented using Benchtop UV-transilluminator and photographed using photo Doc-ItTM imaging system. The molecular size of the amplified products was determined against 1 Kb (+) DNA ladder (enzynomics). Data were created from photographs of gels and analyzed using Gel Analyzer 2010a program.
RESULTS
Isolated bacterium: The isolated bacterium formed 2-3 mm in MacConkey agar medium plates, flat and smooth, colonies with regular margin. This bacterium was identified in Sohag University, Egypt and was found to be Pseudomonas aeruginosa.
Temperate and lytic phages of P. aeruginosa: Pseudomonas aeruginosa was treated with UV irradiation (240 nm) to induce temperate phages. No temperate phages were detected.
Two lytic bacteriophages specific to P. aeruginosa were isolated from a sewage water sample collected from sewage treatment plant. The spot test was used for detection of phages in the collected sample. As shown in Fig. 1 this test indicated that phages of P. aeruginosa were found to be common in the collected sample.
Purification of phages: The single plaque isolation (SPI) technique was used to obtain pure phage isolates of P. aeruginosa. As shown in Fig. 2 the P. aeruginosa specific phages produced different plaques morphologies. The diameters of different plaques were (1 and 3 mm). The plaques were circular single and clear in appearance. These two phage isolates were designated as Pa1 and Pa2.
High titer phage suspensions: The titer of each isolate in one hundred ml was estimated to be 43×108 and 36×108 for Pa1 and Pa2, respectively.
Physical properties of phages
Different pH level for phage infection: The stability of both phage isolates was estimated with different pH levels (pH 4-12) as shown in Table 2. Both phage isolates produced lysed spots at all pH tested levels. Both phage isolates formed the widest lysed spots at pH 6.
Sensitivity to ultraviolet irradiation: As shown in Table 3, the UV irradiation at wave length of 254 nm. was able to inactivate the two isolated phages (Pa1 and Pa2) after exposure for 45 min.
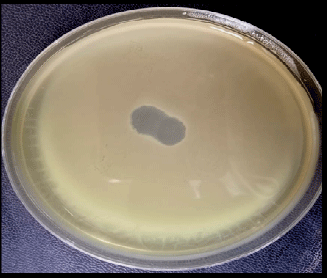 | |
| Fig. 1: | Lysis of Pseudomonas aeruginosa spotted with lytic phage lysate |
| Table 2: | Stability of P. aeruginosa bacteriophages (Pa1 and Pa2) at different pH levels |
 | |
Thermal inactivation point of the phage isolates: Data presented in Table 4 showed that both bacteriophages Pa1 and Pa2 specific to P. aeruginosa were inactivated at 90°C for 10 min.
Morphological properties of phage particles: The two phage isolates (Pa1 and Pa2) specific to P. aeruginosa were found to be of head and tail phages (Fig. 3). Both phage isolates possess long non-contractile tail. Therefore, these two phage isolates could be classified under order Caudovirales, Family Siphoviridae. As shown in Table 5 these two phage isolates were found to be different in their head diameter as well as in tail length and width.
Effect of Pa1 and Pa2 phages on P. aeruginosa as a biocontrol: Data presented in Fig. 4 indicated that, the numbers of P. aeruginosa were determined after 48 h from inoculation with each treatment. Data showed that, the total count of P. aeruginosa decreased after 48 h in broth treated with treated by bacteria+Pa1+Pa2 phages, bacteria+Pa1 phage and bacteria + Pa2 phage respectively, when compared with inoculation with Pseudomonas aeruginosa only.
Molecular characterization
Concentration and purity of genomic DNA: DNA concentration of Pa1 and Pa2 phage isolates were 983.43 and 795.19 μg mL–1, respectively. DNA purity of Pa1 and Pa2 phage isolates was verified by agarose gel electrophoresis and calculated as the ratio of A260/A280 and found to be 1.91 and 1.89 for Pa1 and Pa2, respectively.
Restriction patterns of the isolated DNA: The DNA of each phage isolate (Pa1 and Pa2) was extracted and digested with the restriction enzyme HindIII. As shown in (Fig. 5).
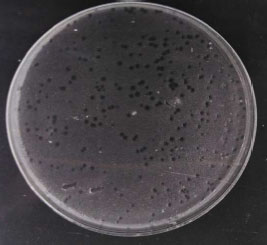 | |
| Fig. 2: | A Petri dish containing different single plaques resulting from lytic bacteriophages specific to Pseudomonas aeruginosa |
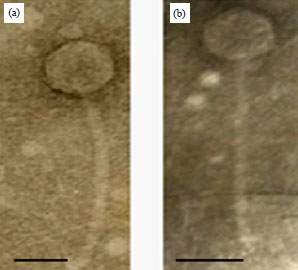 | |
| Fig. 3(a-b): | Electron micrographs of lytic phages (a) Pa1 and (b) Pa2 infected P. aeruginosa stained with phosphotungstic acid |
| Magnification bar = 50 nm | |
| Table 3: | Effect of UV irradiation (254 nm) on the 2 P. aeruginosa specific phages |
 | |
| +: Lysis, -: No lysis | |
| Table 4: | Thermal inactivation points of bacteriophages Pa1 and Pa2 specific to P. aeruginosa exposed to 40-95°C for 10 min |
 | |
| +: Lysis, -: No lysis | |
| Table 5: | Dimensions of bacteriophages Pa1 and Pa2 specific to P. aeruginosa |
 | |
| SD: Standard deviation, (the recorded values represent the average of 5 replicates) | |
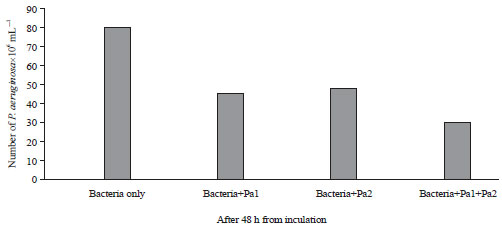 | |
| Fig. 4: | Total number of Pseudomonas aeruginosa in broth media after 48 h with different treatments |
| Bacteria only: Pseudomonas aeruginosa, Pa1: Pa1 Phage, Pa2: Pa2 Phage | |
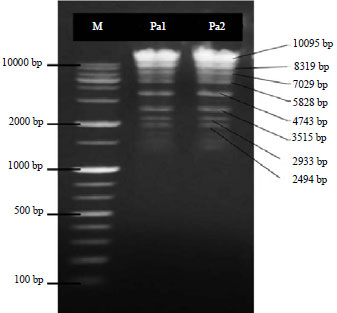 | |
| Fig. 5: | Electrophoresis of HindIII DNA restriction fragments of phage isolates Pa1 and Pa2 specific to P. aeruginosa |
| M: 1 Kb (+) DNA ladder | |
Digestion of Pa1 and Pa2 bacteriophage DNAs with HindIII produced eight fragments with the same molecular sizes ranging from 2.494-10.095 kbp.
Genotyping and polymorphism using RAPD and SCoT-PCR analysis: RAPD and SCoT-PCR analysis were used to study the genetic diversity of DNA products of the two pseudomonas aeuroginosa virulent phages (Pa1 and Pa2) under study. Four primers succeeded to generate reproducible polymorphic DNA products with different sizes with each primer and one primer succeeded only with the Pa1 virulent isolate. The total amplified products were 49 DNA fragments for the 2 virulent isolates, with an average of 9.8 bands/primer (Table 6 and Fig. 6).
Thirty seven bands were found to be unique polymorphic bands for the two virulent phage isolates with all primers (Table 6) with an average of 7.4 polymorphic bands/primer.
| Table 6: | Molecular size and polymorphism percentage using RAPD and SCoT markers |
 | |
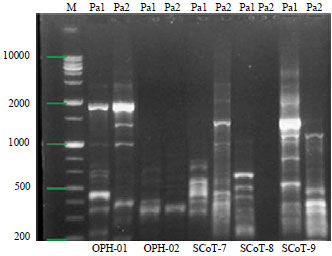 | |
| Fig. 6: | DNA fingerprints of RAPD and SCoT amplification products generated by OPH-01, OPH-02, SCoT-7, SCoT-8 and SCoT-9 primers for Pa1 and Pa2 phage isolates of P. aeuroginosa |
| M: 1 Kb (+) DNA ladder | |
The total percentage of polymorphism was 75.51%. The sizes of the amplified fragments were ranged from 170-4971 bp. with all used primers.
For OPH-01 primer, eight DNA unique bands were observed, five of them with Pa1 phage isolate and three with Pa2 isolated phage. These sizes ranged from 170-4066 bp. While, for OPH-02 primer, only Pa1 phage isolate exhibited 3 unique DNA bands. The band sizes ranged between 314-696 bp. Regarding SCoT-7 primer, the results showed that 3 unique DNA bands were recorded to Pa1 phage isolate and 6 unique bands were detected with Pa2 phage isolate. The molecular sizes of these bands were between 286-4111 bp. For SCoT-9 primer, eight DNA unique bands were appeared only with Pa2 phage isolate and only one DNA unique band was appeared only with Pa1 phage isolate. The sizes of these bands were in the range of 251-4971 bp.
On the other hand, SCoT-8 primer did not show any response with Pa2 but succeeded to generated eight unique DNA bands only with Pa1 phage isolate. The sizes of the generated bands ranged between 205-1291 bp.
DISCUSSION
In this study Pseudomonas aeruginosa was isolated from a sewage water sample after primary treatment collected from Sarpium forest site-Ismailia Governorate, Cairo, Egypt.
The isolated P. aeruginosa was tested for lysogeny using UV irradiation (240 nm). No temperate phages were detected.
Lytic bacteriophages specific to P. aeruginosa were successfully isolated from the collected sewage water sample. Two phage isolates specific to P. aeruginosa were picked and designated Pa1 and Pa2. Similarly results were obtained by Kumari et al.4, Filali et al.32, Alsaffar and Jarallah33, El Didamony et al.34 and Elmaghraby et al.35 isolated bacteriophages specific to P. aeruginosa from sewage water collected from different locations.
Both phages were found to be tolerant to alkaline and acidic reactions. The optimum pH for infection was found to be the same (pH 6) for both phage isolates (Pa1 and Pa2). The 2 lytic phages exhibited the same sensitivity to UV irradiation was found to be 40 min at wave length 245 nm. Bacteriophages Pa1 and Pa2 specific to P. aeruginosa were inactivated at 90°C for 10 min. These results are in agreement with those obtained by Elsharouny36.
The electron micrographs of the two phage isolates (Pa1 and Pa2) indicated that both phages were of head and tail type. The particle dimensions of these two phage isolates were found to be different in their head diameter as well as in tail length and width, this may indicate that these two phages are belonging to Caudovirales, Family Siphoviridae. These results are in agreement with those obtained by Garbe et al.37.
The obtained data, it was found that the isolated virulent phages (Pa1 and Pa2) specific for Pseudomonas aeruginosa were efficiently active in reduction the number of the bacterium when compared with inoculation with Pseudomonas aeruginosa only after 48 h from inoculation. This result is in agreement with those of Lim et al.38, Hassan39 and Carstens et al.40 when evaluated the efficacy of complex phage cocktail against Pectobacterium carotovorum subsp. carotovorum as a biocontrol agent in potato plant.
To confirm if P. aeuroginosa phage isolates (Pa1 and Pa2) belong to 1 or 2 different types, two molecular genetic techniques (i.e., restriction pattern and PCR analysis) were carried out. The DNA of each phage isolate (Pa1 and Pa2) was extracted and digested with the restriction enzyme HindIII. The two phage isolates (Pa1 and Pa2) exhibited the same DNA restriction pattern. This result indicated that the two virulent phage isolates specific to P. aeuroginosa contained linear double strand (ds) DNA as a viral genome while restriction enzymes are active only on dsDNA molecules. This result is in agreement with those of Grose and Casjens41 and Comeau et al.42.
In this study, RAPD and SCoT-PCR analysis of the 2 virulent P. aeuroginosa phage isolates DNA, showed that among the 49 amplified DNA fragments, 37 bands were unique polymorphic. The sizes of amplified fragments ranged from 170-4971 bp. Generally, RAPD and SCoT-PCR results strongly confirmed that the 2 P. aeuroginosa phage isolates (Pa1 and Pa2) are belonging to two different phage types. So, it could be classified under order Caudovirales, Family Siphoviridae since, both possess long non-contractile tail. Similar results were obtained by Comeau et al.42, Gutierrez et al.43 and Winget and Wommack44, who evaluated 26 different bacteriophages infecting different bacterial strains using RAPD-PCR.
Based on the obtained results, can be recommend in the future to use the phage as a safe effective biological control agents against the plant pathogen P. aeuroginosa.
CONCLUSION
In this study, different physical, molecular characteristics and the effect of phage isolates on P. aeruginosa as a biocontrol under lab condition were applied. These two phages are belonging to Caudovirales, Family Siphoviridae. Based on the obtained results, found that both phages are highly stable. Also, both phages in this study could be used as biological control agents against the plant pathogen P. aeuroginosa.
SIGNIFICANCE STATEMENT
This study confirmed that lytic phage can be use as a effective agent against the plant pathogen P. aeuroginosa. The study contributes to using the phages as biological control under environmental condition.
ACKNOWLEDGMENT
Author express appreciation to Prof. A.M. Hammad, Department of Microbiology, Faculty of Agriculture, Minia University, Minia, Egypt, for facilities, valuable advice and comments on the manuscript. Also, I would like to thank Dr. A. A. Aboulila, Genetics Department, Faculty of Agriculture, Kafrelsheikh University, Kafr El-Sheikh, Egypt, for helping me in molecular study in this research.
REFERENCES
- Mooij, M.J., E. Drenkard, M.A. Llamas, C.M.J.E. Vandenbroucke-Grauls, P.H.M. Savelkoul, F.M. Ausubel and W. Bitter, 2007. Characterization of the integrated filamentous phage Pf5 and its involvement in small-colony formation. Microbiology, 153: 1790-1798.
CrossRefDirect Link - Gellatly, S.L. and R.E. Hancock, 2013. Pseudomonas aeruginosa: New insights into pathogenesis and host defenses. Pathog. Dis., 67: 159-173.
CrossRefDirect Link - Walker, S.J., 1988. Major spoilage micro-organisms in milk and dairy products. Int. J. Dairy Technol., 41: 91-92.
CrossRefDirect Link - Kumari, S., K. Harjai and S. Chhibber, 2009. Characterization of Pseudomonas aeruginosa PAO specific bacteriophages isolated from sewage samples. Am. J. Biomed. Sci., 1: 91-102.
CrossRefDirect Link - Shew, H.D. and G.B. Lucas, 1991. Philippine Bacterial Leaf Spot. In: Compendium of Tobacco Diseases, Shew, H.D. and G.B. Lucas (Eds.). The American Phytopathological Society, St. Paul, MN., USA., ISBN: 9780890541173, pp: 33.
Direct Link - Yu, L., X.Y. Qin, J. Du, A.Y. Wang and Y.Y. Zhao et al., 2008. Bacterial leaf spot of tobacco caused by Pseudomonas aeruginosa in China. Plant Pathol., 57: 774-774.
CrossRefDirect Link - Kominos, S.D., C.E. Copeland and B. Grosiak, 1972. Mode of transmission of Pseudomonas aeruginosa in a burn unit and an intensive care unit in a general hospital. Applied Environ. Microbiol., 23: 309-312.
Direct Link - Sillankorva, S., P. Neubauer and J. Azeredo, 2008. Isolation and characterization of a T7-like lytic phage for Pseudomonas fluorescens. BMC Biotechnol., Vol. 8.
CrossRefDirect Link - Piracha, Z.Z., U. Saeed, A. Khurshid and W.N. Chaudhary, 2014. Isolation and partial characterization of virulent phage specific against Pseudomonas aeruginosa. Global J. Med. Res., 14: 1-8.
Direct Link - Green, S.K., M.N. Schroth, J.J. Cho, S.D. Kominos and V.B. Vitanza-Jack, 1974. Agricultural plants and soil as a reservoir for Pseudomonas aeruginosa. Applied Microbiol., 28: 987-991.
Direct Link - Loessner, M.J., R.B. Inman, P. Lauer and R. Calendar, 2000. Complete nucleotide sequence, molecular analysis and genome structure of bacteriophage A118 of Listeria monocytogenes: Implications for phage evolution. Mol. Microbiol., 35: 324-340.
CrossRefDirect Link - Atterbury, R.J., M.A.P. van Bergen, F. Ortiz, M.A. Lovell and J.A. Harris et al., 2007. Bacteriophage therapy to reduce Salmonella colonization of broiler chickens. Applied Environ. Microbiol., 73: 4543-4549.
CrossRefPubMedDirect Link - Wagenaar, J.A., M.A.P. van Bergen, M.A. Mueller, T.M. Wassenaar and R.M. Carlton, 2005. Phage therapy reduces Campylobacter jejuni colonization in broilers. Vet. Microbiol., 109: 275-283.
CrossRefDirect Link - Ortiz-Herrera, M., A. Geronimo-Gallegos, F. Cuevas-Schacht, L. Perez-Fernandez and R. Coria-Jimenez, 2004. [RAPD-PCR characterization of Pseudomonas aeruginosa strains obtained from cystic fibrosis patients]. Salud Publica Mex., 46: 149-157, (In Spanish).
PubMedDirect Link - Fothergill, J.L., J. White, J.E. Foweraker, M.J. Walshaw, M.J. Ledson, E. Mahenthiralingam and C. Winstanley, 2010. Impact of Pseudomonas aeruginosa genomic instability on the application of typing methods for chronic cystic fibrosis infections. J. Clin. Microbiol., 48: 2053-2059.
CrossRefDirect Link - Hafiane, A. and M. Ravaoarinoro, 2011. Characterization of Pseudomonas aeruginosa strains isolated from cystic fibrosis patients by different typing methods. Pathol. Biol., 59: e109-e114.
CrossRefDirect Link - Goto, S. and S. Enomoto, 1970. Nalidixic acid cetrimide agar. A new selective plating medium for the selective isolation of Pseudomonas aeruginosa. Jpn. J. Microbiol., 14: 65-72.
CrossRefPubMedDirect Link - Prinsloo, H.E., 1966. Bacteriocins and phages produced by Serratia marcescens. J. Gen. Microbiol., 45: 205-212.
CrossRefDirect Link - Allen, O.N., 1959. Experiments in Soil Bacteriology. 3rd Edn., Burges Publishing Co., Minneopolis, USA.
Direct Link - Adams, M.H., 1966. The Bacteriophages. Interscience Publishers Inc., New York, USA., pp: 447-461.
Direct Link - Kiraly, Z., Z. Klement, F. Solymosy and J. Voros, 1970. Methods in Plant Pathology with Special Reference to Breeding for Disease Resistance. 2nd Edn., Academia Kiado, Budapest, Hungary, pp: 183-192.
Direct Link - Sambrook, J., E.F. Fritsch and T.A. Maniatis, 1989. Molecular Cloning: A Laboratory Manual. 2nd Edn., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, USA., ISBN-13: 9780879695774, Pages: 397.
Direct Link - Hammad, A.M.M., H. Zahran, M.S. Ahmed and A. Ragab, 2016. Isolation and characterization of bacteriophages specific to root nodule bacterium Rhizobium leguminosarum bv. viciae. J. Basic Applied Sci. Res., 6: 17-25.
Direct Link - Hayat, M.A. and S.E. Miller, 1990. Negative Staining. McGraw-Hill Publishing Co., USA., ISBN-13: 9780070273221, Pages: 253.
Direct Link - Stacey, G., L.A. Pocratsky and V. Puvanesarajah, 1984. Bacteriophage that can distinguish between wild-type Rhizobium japonicum and a non-nodulating mutant. Applied Environ. Microbiol., 48: 68-72.
Direct Link - Clark, H.E., E.F. Geldrich, P.W. Kabler and C.B. Huff, 1958. Applied Microbiology. 1st Edn., International Book Company, New York, pp: 27-53.
Direct Link - Maniatis, T., E.F. Fritsch and J. Sambrock, 1982. Molecular Cloning: A Laboratory Manual. 3rd Edn., Cold Spring Harbor Laboratory, Cold Spring Harbor, New York, USA.
Direct Link - Peacock, A.C. and C.W. Dingman, 1968. Molecular weight estimation and separation of ribonucleic acid by electrophoresis in agarose-acrylamide composite gels. Biochemistry, 7: 668-674.
CrossRefPubMedDirect Link - Williams, J.G.K., A.R. Kubelik, K.J. Livak, J.A. Rafalski and S.V. Tingey, 1990. DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res., 18: 6531-6535.
CrossRefPubMedDirect Link - Filali, B.K., J. Taoufik, Y. Zeroual, F.Z. Dzairi, M. Talbi and M. Blaghen, 2000. Waste water bacterial isolates resistant to heavy metals and antibiotics. Curr. Microbiol., 41: 151-156.
CrossRefDirect Link - Alsaffar, M. and E.M. Jarallah, 2016. Isolation and characterization of lytic bacteriophages infecting Pseudomonas aeruginosa from sewage water. Int. J. PharmTech Res., 9: 220-230.
Direct Link - El Didamony, G., A. Askora and A.A. Shehata, 2015. Isolation and characterization of T7-like lytic bacteriophages infecting multidrug resistant Pseudomonas aeruginosa isolated from Egypt. Curr. Microbiol., 70: 786-791.
CrossRefDirect Link - Elmaghraby, I., F. Carimi, A. Sharaf, E.M. Marei and A.M.M. Hammad, 2015. Isolation and identification of Bacillus megaterium bacteriophages via AFLP technique. Curr. Res. Bacteriol., 8: 77-89.
CrossRefDirect Link - Garbe, J., A. Wesche, B. Bunk, M. Kazmierczak and K. Selezska et al., 2010. Characterization of JG024, a Pseudomonas aeruginosa PB1-like broad host range phage under simulated infection conditions. BMC Microbiol., Vol. 10.
CrossRefDirect Link - Lim, J.A., S. Jee, D.H. Lee, E. Roh, K. Jung, C. Oh and S. Heu, 2013. Biocontrol of Pectobacterium carotovorum subsp. carotovorum using bacteriophage PP1. J. Microbiol. Biotechnol., 23: 1147-1153.
CrossRefPubMedDirect Link - Hassan, E.O., 2017. Effect of different bacteriophage isolates on managing potato soft rot caused by Pectobacterium carotovorum subsp. carotovorum. Int. J. Scient. Eng. Res., 8: 719-730.
Direct Link - Carstens, A., A. Djurhuus, W. Kot, D. Jacobs-Sera, G. Hatfull and L. Hansen, 2018. Unlocking the potential of 46 new bacteriophages for biocontrol of Dickeya solani. Viruses, Vol. 10, No. 11.
CrossRefDirect Link - Grose, J.H. and S.R. Casjens, 2014. Understanding the enormous diversity of bacteriophages: The tailed phages that infect the bacterial family Enterobacteriaceae. Virology, 468-470: 421-443.
CrossRefDirect Link - Comeau, A.M., S. Short and C.A. Suttle, 2004. The use of Degenerate-Primed Random Amplification Of Polymorphic DNA (DP-RAPD) for strain-typing and inferring the genetic similarity among closely related viruses. J. Virol. Methods, 118: 95-100.
CrossRefDirect Link - Gutierrez, D., A.M. Martin-Platero, A. Rodriguez, M. Martinez-Bueno, P. Garcia and B. Martinez, 2011. Typing of bacteriophages by Randomly Amplified Polymorphic DNA (RAPD)-PCR to assess genetic diversity. FEMS Microbiol. Lett., 322: 90-97.
CrossRefDirect Link - Winget, D.M. and K.E. Wommack, 2008. Randomly amplified polymorphic DNA PCR as a tool for assessment of marine viral richness. Applied Environ. Microbiol., 74: 2612-2618.
CrossRefDirect Link