
Aziiz Mardanarian Rosdianto
Department of Pharmacology and Clinical Pharmacy, Faculty of Pharmacy, Padjadjaran University, Jalan Raya Bandung-Sumedang km 21, Jatinangor, 45363 West Java, Indonesia
Irma Melyani Puspitasari
Department of Pharmacology and Clinical Pharmacy, Faculty of Pharmacy, Padjadjaran University, Jalan Raya Bandung-Sumedang km 21, Jatinangor, 45363 West Java, Indonesia
Ronny Lesmana
Division of Molecular Physiology, Department of Anatomy, Physiology and Cell Biology, Faculty of Medicine, Padjadjaran University, Jalan Raya Bandung-Sumedang km 21, Jatinangor, 45363 West Java, Indonesia
Jutti Levita
Department of Pharmacology and Clinical Pharmacy, Faculty of Pharmacy, Padjadjaran University, Jalan Raya Bandung-Sumedang km 21, Jatinangor, 45363 West Java, Indonesia
LiveDNA: 62.1290
Pakistan Journal of Biological Sciences
Year: 2020 | Volume: 23 | Issue: 3 | Page No.: 264-270







ABSTRACT
Background and Objective: Flavonols in plants are catalyzed by flavonol synthase (FLS) enzyme. FLS was reported expressed in flowers and fruits, i.e., Dianthus caryophyllus L. (Caryophyllaceae), Petunia hybrida Hort. (Solanaceae), Arabidopsis thaliana L. (Brassicaceae), Citrus unshiu Marc. (Rutaceae). However, none reported about FLS in medicinal plants, particularly those which possess anti-inflammatory activity. This study was aimed to extract and identify FLS in the rhizome of Boesenbergia rotunda (Zingiberaceae) and to determine quercetin in the ethanol extract of the rhizome. Materials and Methods: The protein extraction of the rhizome was carried out by employing Laing and Christeller’s (2004) and Wang’s (2014) methods. The extracted-proteins were separated by using SDS-PAGE, followed by the measurement of FLS intensity by using Gel Analyzer. The FLS-1 of recombinant A. thaliana was employed as the standard. The determination of quercetin in the rhizome was carried out using LC-MS. Results: The FLS occurred as a thick band at 38 kDa with intensity 116-158. The LC chromatogram of the extract indicated a small peak at 7.94 min similar to that of quercetin standard. The MS spectra at 7.94 min indicated that quercetin is present in the B. rotunda rhizome (m/z = 303.0549). The concentration of quercetin in the extract is 0.022% w/v. Conclusion: The FLS, an enzyme which plays an important role in producing quercetin, was detected in B. rotunda rhizome planted in Indonesia. As a consequence, quercetin in a small amount, was also quantified in the rhizome of this plant. This report will add a scientific insight of B. rotunda for biological sciences.
PDF Abstract XML References Citation
Copyright: © 2020. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Aziiz Mardanarian Rosdianto, Irma Melyani Puspitasari, Ronny Lesmana and Jutti Levita, 2020. Determination of Quercetin and Flavonol Synthase in Boesenbergia rotunda Rhizome. Pakistan Journal of Biological Sciences, 23: 264-270.
DOI: 10.3923/pjbs.2020.264.270
URL: https://scialert.net/abstract/?doi=pjbs.2020.264.270
DOI: 10.3923/pjbs.2020.264.270
URL: https://scialert.net/abstract/?doi=pjbs.2020.264.270
INTRODUCTION
The secondary metabolites of plants, e.g., flavonoids, have been known to represent the plant’s pharmacological activity. Flavonols, a small class of flavonoids, have been widely reported to exert anti-inflammatory activity. The FLS catalyzes the conversion of dihydroflavonols to flavonols, i.e., dihydrokaempferol to kaempferol and dihydroquercetin to quercetin, to form a variety of flavonols and glycosidic derivatives1-3.
Flavonol synthase (FLS) is an enzyme which is contained in flowers, e.g., Arabidopsis thaliana L. (Brassicaceae)1,4,5, Petunia hybrida Hort. (Solanaceae)2, Dianthus caryophyllus L. (Caryophyllaceae)6 and also in fruits, e.g., Citrus unshiu Marc. (Rutaceae)7. This particular enzyme belongs to the 2-oxoglutarate iron-dependent oxygenase family8. The oxidation reaction, which introduced the C2-C3 double bond, was considered to be specific in dihydroflavonol substrates9.
Extraction of protein from various parts of a plant is considered as a challenging task. There are many obstacles in this process, i.e., the thickness of cellulose walls in the leaves, the secondary metabolites which interfere with the protein, etc10,11. However, a few abundant proteins had successfully been identified from the leaves of Gynura procumbens by using the proteomic approach. The use of trichloroacetic acid-acetone wash before the protein extraction, could produce distinct protein bands because these solvents rinse out organic-soluble substances, thus precipitate the proteins and other insoluble substances12,13. SDS-PAGE, a polyacrylamide gel electrophoresis with a sodium dodecyl sulfate (SDS)-containing buffer, is usually employed to separate the protein. The protein is set apart by the application of an electrical field. The polar part of the buffer is SDS, a negatively-charged amphipathic detergent, while the dodecyl portion, CH3 (CH2)11, is hydrophobic. When SDS fastens a soluble protein, the hydrophilic portion of the SDS slips into the hydrophobic center of the protein and denatures the protein14,15.
In this study, SDS-PAGE was employed to extract and to separate protein from the rhizome of Boesenbergia rotunda (Zingiberaceae). The quercetin in the ethanol extract of the rhizome was determined by using the standard addition LC-MS method.
MATERIALS AND METHODS
Study area: The study was carried out at the Central Laboratory of Universitas Padjadjaran, Jl. Raya Bandung- Sumedang km 21, West Java, Indonesia, 45363 from April, 2018 to January, 2019.
Plant material and identification: The rhizome of B. rotunda was obtained from the Research Institute for Spices and Medicinal Plants (Balittro) Manoko Lembang, West Java, Indonesia (http://balittro.litbang.pertanian.go.id/?p=993& lang=en). The plant was taxonomically identified at the Laboratory of Plant Taxonomy, Department of Biology, Faculty of Mathematics and Natural Sciences, Universitas Padjadjaran, Indonesia (Letter No. 450/HB/10/2017).
Chemicals and other materials: Standard recombinant A. thaliana FLS1 (Cusabio Cat. No. # CSB-MP842601DOA), Coomassie blue (Simply Blue™ Safe Stain), PierceTM silver stain kit (Thermo Scientific Cat. No. #24612), Ladder SeeBlue™ Plus2 pre-stained standard(Invitrogen™ Cat. No. #LC5925), quercetin hydrate (Sigma-AldrichCAS No. 849061-97-8) were purchased from Quartiz Laboratories (a branch of PT Indogen Intertama (http://indogen.id/).
FLS separation and intensity measurement: The rhizome of B. rotunda (8 g) was washed with tap water and was ground in liquid nitrogen to a fine powder. About 250 mg of the powder was extracted according to Laing and Christeller16 and Wang et al.17 methods with a few modifications. The protein was separated using SDS-PAGE for 150 min 80-100 V. The protein bands were stained using Coomassie blue and silver stain compared to that of standard recombinant A. thaliana FLS1. The intensity of FLS in the rhizome was calculated using GelAnalyzer (http://www.gelanalyzer.com/).
Extraction of secondary metabolites in the rhizome: About 500 g of the rhizome was macerated in ethanol 95% for 24 h. The macerate was filtered and the residue was re-macerated in ethanol 95% for 2×24 h. The solvent was evaporated at 60-70°C 80 rpm in vacuum until a viscous extract was obtained.
Identification of flavonoids in the ethanol extract of the rhizome using UV spectrophotometry and determination of quercetin using liquid chromatography-mass spectrometry (LC-MS): The presence of quercetin in the ethanol extract of B. rotunda (EEBR) was determined according to a method proposed by Bajkacz et al.18 with a few modifications. Quercetin hydrate was used as the standard.
Standard stock solution of quercetin was prepared by dissolving the analyte in ethanol (LC grade) obtaining a concentration of 100 μg mL–1. Working solutions for calibration standard-addition samples were prepared by diluting the stock solution of quercetin with ethanol and spiked it into the extract solution. This process resulted in the quercetin-extract working solutions of 2.56, 1.28, 0.64 and 0.32μg mL–1. The 0.32μg mL–1 solution was run on a double-beam spectrophotometry to obtain the spectrum of flavonols in the EEBR.
Eventually, the 2.56, 1.28, 0.64 and 0.32μg mL–1 working solutions were filtered using a 0.2 μm millipore membrane prior to be injected into the Acquity™ Ultra Performance LC BEH shield RP18 1.7 μm, 2.1×100 mm column of Waters Acquity™ Ultra Performance LC in tandem with Waters Xevo™ Qt of MS. Mobile phase used was a mixture of phase A (H2O-formic acid 0.1%) and phase B (methanol). Flow rate was set to 0.2 mL min–1.
Phytochemical screening and identification of flavonols: Phytochemical screening was carried out as per standard method described by Tiwari et al.19 at the Central Laboratory of Universitas Padjadjaran, Indonesia.
RESULTS
FLS separation and intensity measurement: The standard recombinant A. thaliana FLS-1 indicated a thick band at 38 kDa (Fig. 1a). The rhizome of B. rotunda indicated several protein bands (Fig. 1b) and a thick band at 38 kDa. The intensity of FLS in B. rotunda rhizome is 116-158 (Fig. 2).
Identification of flavonoids in the ethanol extract of the rhizome using UV spectrophotometry: The spectrum of B. rotunda extract indicated 2 maxima at 209 and 291 nm (Fig. 3) which confirmed the presence of ring A and ring B of flavonoids.
Determination of quercetin in the ethanol extract of the rhizome using liquid chromatography-mass spectrometry (LC-MS): Liquid chromatography method of EEBR indicated the presence of quercetin (Fig. 4), which was eluted at 7.94 min, similar to quercetin standard. The MS spectrum at 7.94 min revealed the [M+H]+ peak of quercetin standard m/z = 303.0546 (Fig. 5a) and the [M+H]+ peak of quercetin in EEBR m/z = 303.0549 (Fig. 5b). The concentration of quercetin in Indonesian B. rotunda is 0.2 mg g–1 or 0.022% (linear regression equation).
 | |
| Fig. 1(a-b): | SDS-electropherograms of rhizome extract, (a) Coomassie blue and (b) Silver stain were used as staining reagents |
1: Ladder, 2: FLS-1 standard, 3: B. rotunda, red rectangle indicates the position of FLS band | |
DISCUSSIONS
The high intensity of FLS in B. rotunda is understandable, because this plant has been reported in containing flavonols and other phytochemical constituents20. The presence of FLS in B. rotunda confirms the conversion of dihydroflavonol to flavonol catalysed by this enzyme. Furthermore, plants belonged to Zingiberaceae family, have been proven in exhibiting anti-inflammatory activity by reducing prostaglandin production. This activity was predicted due to its flavonoids content21.
Many spectroscopic studies have revealed that flavons (flavonoids with a 2,3-double bond and 4-keto group) and flavonols (flavonoids with a 2,3-double bond, 3-OH and 4-keto groups) possess 2 main absorption bands: band 1 (300-385 nm) belongs to ring B, whereas band 2 (250-295 nm) for ring A. In flavanones (flavonoids with 4-keto groups) and dihydroflavonols (flavonoids with with 3-OH and 4-keto groups), band A appears as a shoulder peak at 300-330 nm and band B, in the 277-295 nm range, is the main peak 22-24.
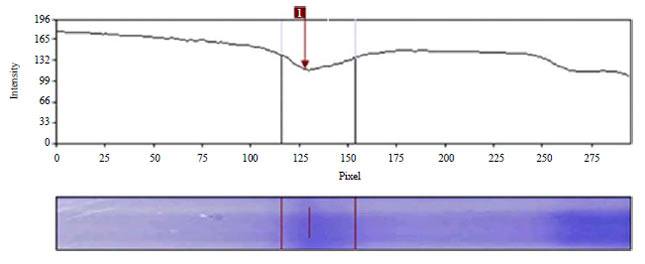 | |
| Fig. 2: | Intensity of FLS in B. rotunda rhizome |
Calculated using GelAnalyzer | |
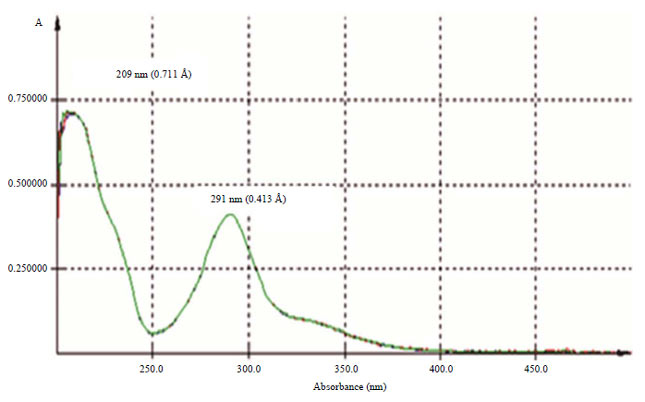 | |
| Fig. 3: | Ultraviolet absorption spectrum of B. rotunda rhizome extract in ethanol (λmax at 209 and 291 nm) |
The concentration of quercetin in Indonesian B. rotunda is 0.2 mg g–1 or 0.022% is less compared to Malaysian B. rotunda (0.58 mg g–1)20. Quercetin was also reported contained in Zingiber zerumbet (3.94 mg g–1), Z. officinale (3.66 mg g–1), Alpinia galanga (29.535 mg g–1), A. officinarum (33.66 mg g–1)19,24,25. Flavonoids, e.g., anthocyanins, flavones, isoflavones, flavonols, etc., have been extensively studied26-29. The pathway of flavonoid biosynthesis has been well understood and the k ey biosynthetic enzymes have been identified29-31.
Quercetin, although presents in a low quantity in the rhizome of B. rotunda, might contribute to this plant’s activity. Further exploration on this plant’s pharmacological activity, is still needed, to develop this plant as the future plant-based therapy.
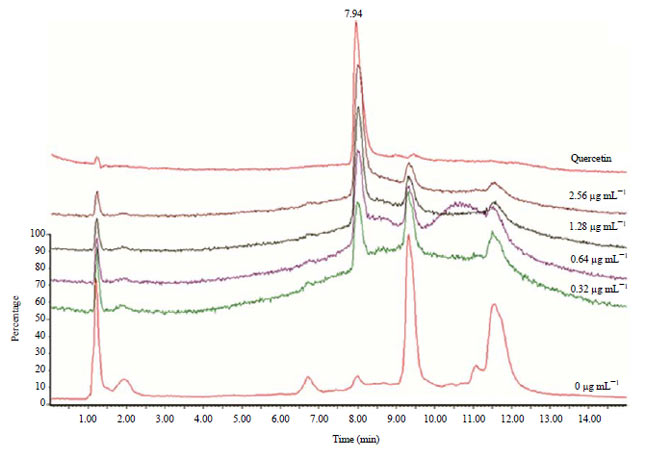 | |
| Fig. 4: | LC chromatograms of quercetin-spiked EEBR |
Quercetin is eluted at 7.94 min | |
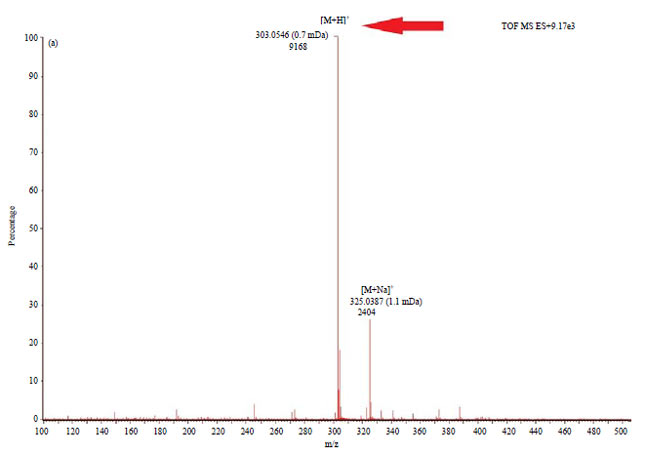 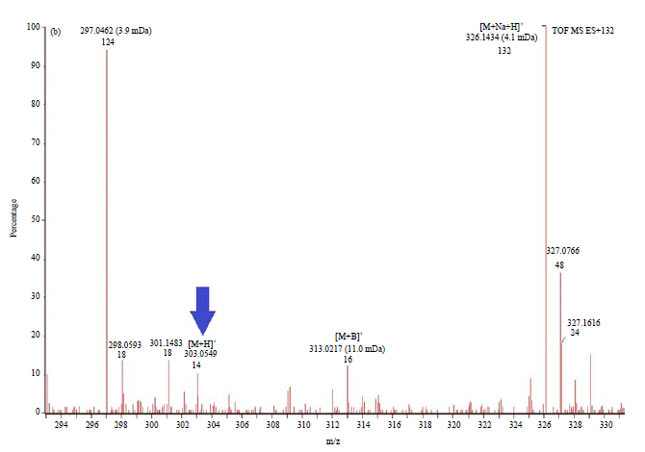 | |
| Fig. 5(a-b): | Mass spectrum of (a) Quercetin standard [M+H]+ m/z = 303.0546 (indicated by red arrow) and (b) Quercetin in EEBR [M+H]+ m/z = 303.0549 (indicated by blue arrow) |
CONCLUSION
Flavonol synthase, an enzyme which plays an important role in producing quercetin, was detected in B. rotunda rhizome planted in Indonesia. As a consequence, quercetin in a small amount, was also quantified in the rhizome of this plant. This report will add a scientific insight of B. rotunda for biological sciences.
SIGNIFICANCE STATEMENT
This study discovers the presence of flavonol synthase enzyme and quercetin in Boesenbergia rotunda rhizome planted in Indonesia, that can be beneficial for the development of traditional medicinal plants and drug discovery. This study will help the researcher to uncover the critical area of B. rotunda’s bioactivity that many researchers were not able to explore. Thus, a new theory on the plant’s pharmacological activity, may be arrived at.
ACKNOWLEDGMENT
This study, which is a part of Aziiz Mardanarian Rosdianto’s doctoral dissertation at the Doctoral Program of Pharmacy, Faculty of Pharmacy, Universitas Padjadjaran, was facilitated by Academic-Leadership Grant (ALG) (No. 872/UN6.3.1/LT/2018) of Universitas Padjadjaran. This study was supervised by Prof. Dr. Jutti Levita, Irma Melyani Puspitasari, Ph.D. and Ronny Lesmana, Ph.D.
REFERENCES
- Owens, D.K., A.B. Alerding, K.C. Crosby, A.B. Bandara, J.H. Westwood and B.S. Winkel, 2008. Functional analysis of a predicted flavonol synthase gene family in Arabidopsis. Plant Physiol., 147: 1046-1061.
PubMed - Kuhn, B.M., M. Geisler, L. Bigler and C. Ringli, 2011. Flavonols accumulate asymmetrically and affect auxin transport in Arabidopsis. Plant Physiol., 156: 585-595.
CrossRefDirect Link - Li, C.L., Y.C. Bai, S.J. Li, H. Chen and X.Y. Han et al., 2012. Cloning, characterization and activity analysis of a flavonol synthase gene FtFLS1 and its association with flavonoid content in tartary buckwheat. J. Agric. Food Chem., 60: 5161-5168.
CrossRefDirect Link - Preuß, A., R. Stracke, B. Weisshaar, A. Hillebrecht, U. Matern and S. Martens, 2009. Arabidopsis thaliana expresses a second functional flavonol synthase. FEBS Lett., 583: 1981-1986.
CrossRefDirect Link - Nguyen, N.H., J.H. Kim, J. Kwon, C.Y. Jeong and W. Lee et al., 2016. Characterization of Arabidopsis thaliana FLAVONOL SYNTHASE 1 (FLS1)-overexpression plants in response to abiotic stress. Plant Physiol. Biochem., 103: 133-142.
CrossRefDirect Link - Stich, K., T. Eidenberger, F. Wurst and G. Forkmann, 1992. Flavonol synthase activity and the regulation of flavonol and anthocyanin biosynthesis during flower development in Dianthus caryophyllus L. (Carnation). Zeitschrift für Naturforschung C, 47: 553-560.
CrossRefDirect Link - Wellman, F., R. Lukacin, T. Moriguchi, L. Britsch, E. Schiltz and U. Matern, 2002. Functional expression and mutational analysis of flavonol synthase from Citrus unshiu. Eur. J. Biochem., 269: 4134-4142.
CrossRefDirect Link - Xu, F., L. Li, W. Zhang, H. Cheng, N. Sun, S. Cheng and Y. Wang, 2012. Isolation, characterization and function analysis of a flavonol synthase gene from Ginkgo biloba. Mol. Biol. Rep., 39: 2285-2296.
CrossRefDirect Link - Holton, T.A., F. Brugliera and Y. Tanaka, 1993. Cloning and expression of flavonol synthase from Petunia hybrid. Plant J., 4: 1003-1010.
CrossRefDirect Link - Wang, W., R. Vignani, M. Scali and M. Cresti, 2006. A universal and rapid protocol for protein extraction from recalcitrant plant tissues for proteomic analysis. Electrophoresis, 27: 2782-2786.
CrossRefDirect Link - Pavoković, D., B. Križnik and M. Krsnik-Rasol, 2012. Evaluation of protein extraction methods for proteomic analysis of non-model recalcitrant plant tissues. Croatica Chemica Acta, 85: 177-183.
CrossRefDirect Link - Hew, C.S. and L.H. Gam, 2010. The identification of high abundant proteins in the leaves of Gynura procumbens. Biotechnol. Biotechnol. Equip., 24: 2132-2136.
CrossRefDirect Link - Niu, L., H. Zhang, Z. Wu, Y. Wang, H. Liu, X. Wu and W. Wang, 2018. Modified TCA/acetone precipitation of plant proteins for proteomic analysis. PloS One, Vol. 13, No. 12.
CrossRefDirect Link - Levita, J., D. Rahayu, N.M. Saptarini, A. Nawawi, A. Mutalib and S. Ibrahim, 2011. The inhibition of andrographolide (Andrographis paniculata) on the expression of carbonic anhydrase in LPS-induced human leucocyte cells. Asian J. Plant Sci., 10: 162-165.
CrossRefDirect Link - Roy, S. and V. Kumar, 2014. A practical approach on SDS PAGE for separation of protein. Int. J. Sci. Res., 3: 955-960.
Direct Link - Laing. W. and J. Christeller, 2005. Extraction of proteins from plant tissues. Curr. Protocols Protein Sci.
CrossRefDirect Link - Wang, W., F. Tai and S. Chen, 2008. Optimizing protein extraction from plant tissues for enhanced proteomics analysis. J. Sep. Sci., 31: 2032-2039.
CrossRefPubMedDirect Link - Bajkacz, S., I. Baranowska, B. Buszewski, B. Kowalski and M. Ligor, 2018. Determination of flavonoids and phenolic acids in plant materials using SLE-SPE-UHPLC-MS/MS method. Food Anal. Methods, 11: 3563-3575.
CrossRefDirect Link - Tiwari, P., B. Kumar, M. Kaur, G. Kaur and H. Kaur, 2011. Phytochemical screening and extraction: A review. Int. Pharm. Sci., 1: 98-106.
Direct Link - Ling, J.J., M. Mohamed, A. Rahmat and M.F. Abu Bakar, 2010. Phytochemicals, antioxidant properties and anticancer investigations of the different parts of several gingers species (Boesenbergia rotunda, Boesenbergia pulchella var. attenuata and Boesenbergia armeniaca). J. Med. Plants Res., 4: 27-32.
Direct Link - Fikri, F., N.M. Saptarini, J. Levita, A. Nawawi, A. Mutalib and S. Ibrahim, 2016. Zingiber officinale var. Rubrum reduces the rate of prostaglandin production. Pharmacol. Clin. Pharm. Res., 1: 33-39.
Direct Link - Tsimogiannis, D.I. and V. Oreopoulou, 2006. The contribution of flavonoid C-ring on the DPPH free radical scavenging efficiency. A kinetic approach for the 3′,4′-hydroxy substituted members. Innovative Food Sci. Emerg. Technol., 7: 140-146.
CrossRefDirect Link - Tsimogiannis, D., M. Samiotaki, G. Panayotou and V. Oreopoulou, 2007. Characterization of flavonoid subgroups and hydroxy substitution by HPLC-MS/MS. Molecules, 12: 593-606.
CrossRefPubMedDirect Link - Miean, K.H. and S. Mohamed, 2001. Flavonoid (myricetin, quercetin, kaempferol, luteolin and apigenin) content of edible tropical plants. J. Agric. Food Chem., 49: 3106-3112.
CrossRefPubMedDirect Link - Devi, K.U., T.I. Devi and S.J. Devi, 2018. Essential oil content and antioxidant activity of methanolic rhizome extract of two zingiberace plants in the hill of Manipur. Int. J. Pharm. Sci. Invention, 7: 23-27.
Direct Link - Taylor, L.P. and E. Grotewold, 2005. Flavonoids as developmental regulators. Curr. Opin. Plant Biol., 8: 317-323.
CrossRefPubMedDirect Link - Grotewold, E., 2006. The genetics and biochemistry of floral pigments. Annu. Rev. Plant Biol., 57: 761-780.
CrossRefDirect Link - Subramanian, S., G. Stacey and O. Yu, 2007. Distinct, crucial roles of flavonoids during legume nodulation. Trends Plant Sci., 12: 282-285.
CrossRefDirect Link - Buer, C.S., N. Imin and M.A. Djordjevic, 2010. Flavonoids: New roles for old molecules. J. Integr. Plant Biol., 52: 98-111.
CrossRefDirect Link - Nielsen, K., S.C. Deroles, K.R. Markham, M.J. Bradley, E. Podivinsky and D. Manson, 2002. Antisense flavonol synthase alters copigmentation and flower color in lisianthus. Mol. Breed., 9: 217-229.
CrossRefDirect Link - Tanaka, Y., N. Sasaki and A. Ohmiya, 2008. Biosynthesis of plant pigments: Anthocyanins, betalains and carotenoids. Plant J., 54: 733-749.
CrossRefDirect Link