
Hadeer Yahia Darwish
Department of Biotechnology, Faculty of Science, Taif University, Taif, Saudi Arabia
LiveDNA: 20.30744
Hala Abdelmigid
Department of Biotechnology, Faculty of Science, Taif University, Taif, Saudi Arabia
LiveDNA: 966.5094
Sarah Albogami
Department of Biotechnology, Faculty of Science, Taif University, Taif, Saudi Arabia
LiveDNA: 966.24332
Saqer Alotaibi
Department of Biotechnology, Faculty of Science, Taif University, Taif, Saudi Arabia
LiveDNA: 966.26553
Ahmed Nour El-Deen
Department of Biology, Faculty of Sciences, Taif University, Saudi Arabia
Alaa Alnefaie
Department of Biotechnology, Faculty of Science, Taif University, Taif, Saudi Arabia
Pakistan Journal of Biological Sciences
Year: 2020 | Volume: 23 | Issue: 8 | Page No.: 1025-1036







ABSTRACT
Background and Objective: Rosmarinic acid is considered as one of the most important secondary metabolites in medicinal plants especially of family Lamiaceae. Rosmarinic acid can prevent both the tumor initiation and promotion stages of carcinogenesis. The aim of current study was to evaluate the antiproliferative effects of Hyssopus officinalis and Thymus vulgaris callus crude extracts contained rosmarinic acid on breast cancer cells with correlation to phenylpropanoid biosynthetic pathway genes expression. Materials and Methods: Calli of both plants were maintained on Murashige and Skoog medium supplemented with kinetin and 2,4-D. Rosmarinic acid was determined spectrophotometrically in both seed-germinated plants (control) and callus tissues. Transcriptional profiling of rosmarinic acid pathway genes was performed with RT-PCR system. The human breast cancer cell line MCF-7 was treated with different levels of crude extracts at different time intervals in order to show their effects on the cell proliferation using a cell viability colorimetric assay (MTT). Results: The results showed a significant increase of rosmarinic acid content up to 6.5% in callus compared to control. The transcriptional profile of the selected rosmarinic acid genes in callus tissues indicated significant effects on the rosmarinic acid content in both genotypes. T. vulgaris (90 μg mL–1) and H. officinalis (150 μg mL–1) callus extracts had exhibited highest reduction in the cell MCF-7 viability after 48 h of exposure. Conclusion: It was concluded that rosmarinic acid production increased in callus tissue, showed the higher gene expression levels and remarkably inhibited growth of human breast cancer cell line.
PDF Abstract XML References Citation
Copyright: © 2020. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Hadeer Yahia Darwish, Hala Abdelmigid, Sarah Albogami, Saqer Alotaibi, Ahmed Nour El-Deen and Alaa Alnefaie, 2020. Induction of Biosynthetic Genes Related to Rosmarinic Acid in Plant Callus Culture and Antiproliferative Activity Against Breast Cancer Cell Line. Pakistan Journal of Biological Sciences, 23: 1025-1036.
DOI: 10.3923/pjbs.2020.1025.1036
URL: https://scialert.net/abstract/?doi=pjbs.2020.1025.1036
DOI: 10.3923/pjbs.2020.1025.1036
URL: https://scialert.net/abstract/?doi=pjbs.2020.1025.1036
INTRODUCTION
Medicinal plants have proficiently been utilized for thousands of years in the different fields of ethnobotany and ethnopharmacology as valuable sources for production of secondary metabolites1. Flavonoids, alkaloids, terpenoids, thionins, phenylpropanoid and polypeptides are secondary metabolites have many medical and health benefits and cell protection in humans, these natural products are usually preferred to synthetic drugs2. However, it is difficult to synthesize several natural products. So, continuous supply of large amounts of natural secondary product is important demand. Hence, modern productive and bio-technological approach under controlled culture conditions represents a valuable and desirable technique for the industrial mass creation of phytochemicals on an industrial scale from calli3.
Plant biotechnology has appropriate methods including cell culture, tissue culture, genetic engineering and molecular markers which can increase gene expression and production4-6. Among of these approaches, plant tissue culture considered as an affordable productive technique of secondary products and has been improved for optimization of culture conditions and increase the generation of natural products in in vitro1. Moreover, illustration and identification of these compounds biosynthesis and the exact genes that catalyze their synthesis are crucial to enhance the yield of these compounds7. Advance comparative analysis in sequencing and transcriptome exploration has provided valuable candidates for biosynthetic pathway analysis.
The bioactive compound, rosmarinic acid (RA) is a phenol with multifunctional activity that belongs to plant phenylpropanoids, a group of phenylalanine derived active secondary products, such as lignins, flavonols, isoflavonoids, phenolic acids and anthocyanins8. They have various essential functions in plants. For instance, lignins support cell walls9, flavonoids and isoflavonoids play roles in UV filtration10,11 and symbiotic fixation of nitrogen12 and anthocyanins could act to protect plants against harmful photo-oxidative effects and UV irradiation13. Phenylpropanoids could have some positive effects on the health of human as anticancer14 and anti-inflammatory15.
Breast cancer is a multifactorial disease rising from breast tissues forming a tumor. In Saudi Arabia, it is the second most diagnosed type of cancer in women, some studies suggested that breast cancer would score even higher incidence rates in the upcoming years16-19. Current cancer treatments are based on the induction of apoptosis in cancer cells to prevent progression, although they proved to be effective but still have limitations such as drug resistance, also these treatments might as well contribute to cancer reoccurrence and metastasis20,21.
In previous reports, biotechnological and metabolic engineering approaches were applied to produce RA in plants22-24. Plants still unsatisfactory resources toward the growing demand of RA. The amino acids, L-phenylalanine and L-tyrosine have been determined as precursors of the biosynthetic pathway of RA in plants species coleus and thyme25. In phenylalanine pathway of RA biosynthesis, phenylalanine ammonia-lyase (PAL) works to catalyze the oxidative deamination of phenylalanine, to form t-cinnamic acid and cinnamic acid 4-hydroxylase (C4H) which it can lead to the para-hydroxyl group into the aromatic ring of t-cinnamic acid, where 4-hydroxycinnamic acid CoA-ligase (4CL) promotes catalysts activation of cinnamic acids with coenzyme A. Moreover, tyrosine aminotransferase (TAT) in the tyrosine-derived pathway encourage, as a catalytic agent, the transamination process of tyrosine to 4-hydroxyphenylpyruvic acid, which converted to 4-hydroxyphenyllactic acid by the activity of hydroxyphenylpyruvate (HPPR). The RA is formed finally by the catalytic effect of RA synthase (RAS) on the two synthesized precursors26-28.
Lamiaceae is one of the most extended plant families in terms of ethnomedicine. Its pharmaceutical value is based on the unstable oils level29. Numerous plants of this family have some tags e.g. they can be easily cultivated by stem cuttings, their aromatic characteristics and includes various secondary metabolites as well. This family comprises approximately 236 genera, around 7,200 species and includes rich and economic species that considerable for humans, such as mint, basil, sage, oregano and thyme. Furthermore, the genes from RA biosynthesis from many species of lamiaceae such as C. blumei25, Agastache rugosa30, Perilla frutescens 31, Melissa officinalis32 and Salvia miltiorrhiza33 were determined and identified as RA biosynthetic genes. These genes are shared by many biosynthetic secondary products beside RA, so global searches of these genes in the pathway of RA biosynthesis would be contributory to achieve quantity increment of RA in lamiaceae.
The current study was accomplished to examine the role of plant tissue culture technology on the enhancement of rosmarinic acid in two members of family Lamiaceae, as well as to determine whether the culture conditions influence the phenylpropanoid biosynthetic pathway genes expression. The two-species selected for this study are Hyssopus officinalis (Hyssop) and Thymus vulgaris (Thyme) plants. Another objective of this research was to evaluate whether thyme and hyssop plants seed embryo and callus extracts possess an antiproliferative and toxic effect against human breast cancer cells MCF-7.
MATERIALS AND METHODS
Study area: The study was carried out at Biotechnology Department, Plant Tissue Culture and Biotechnology Lab from September, 2017-April, 2019.
Plant materials and growth conditions: Seeds of H. officinalis and T. vulgaris were purchased from Amazon Company. Seeds of equal numbers and uniform size were washed thoroughly under running tap water for 15 min with few drops of mild detergent. They were immersed in sterilized solution (5.0% sodium hypochlorite plus 2 drops of Tween-20) for 15 min under laminar air flow chamber. Seeds were then rinsed with autoclaved distilled water for 3 times (5 min each) and then transferred into 70% (v/v) ethanol solution for 30 sec and rinsed with sterile distilled water 3 times (5 min each). Sterilized seeds were inoculated on the surface of sterilized watered cotton. Germinated seeds were then cultured on Murashige and Skoog34 medium without hormones and then transferred to growth chamber (16 h light/8 h dark photoperiod, at 24°C) for further growth. After 4 weeks, explants were subcultured on the selected basal solidified medium (MS medium with plant hormones 2.0 mg L–1 kinetin (KIN), 2.0 mg L–1 2,4-dichlorophenoxyacetic acid (2,4-D), 8 g L–1 agar and 3% sucrose which produced highest yield of callus. All cultures were maintained in the dark condition at 25±2°C. Data were recorded after 6 weeks on fresh and dry weight of callus. Germinated seeds grown in MS medium without hormones were used for comparison as control plants.
Estimation of the RA content: Rosmarinic acid was extracted from control seedlings and dried callus (200 mg) of each plant species by Soxhlet continuous extraction device after grinding, filtration and concentration at 45°C to get 5 mL of sample (crude). Weight of crude was recorded and the RA content was determined photometrically35. The extraction method adopted the use of Fe2+ ions in a complex reaction. The 40 μL of sample was mixed with 4 mL of Tris/acetate buffer (pH 6.0). The FeSO4 level was 0.5 g L–1 in the buffer. The mixture was kept at room temperature for 1 h in the darkness. The absorbance was read at 572 nm in the reaction medium. Calibration was performed with standard of rosmarinic acid.
Crude extract dilutions: The extracts of studied plants were diluted in 1 mL of DMSO (Dimethyl sulfoxide) to make a stock solution with final concentrations as follow: Hyssop seedling = 700 mg mL–1, Hyssop callus = 600 mg mL–1, Thyme seedling = 300 mg mL–1, Thyme callus = 300 mg mL–1. Then each one of the stocks was further diluted in serum free medium to get the following concentrations (30, 90, 150 and 210 μg mL–1). These dilutions were freshly prepared prior treating the MCF-7 cells.
Total RNA extraction: Total RNA was extracted from seed embryo plants and callus tissue (25-d old) using Trizol reagent (Life Technologies, Ambion) as explained in the protocol of Chomczynski36. Tissues were ground and homogenized in Trizol solution (100 mg of tissue/1 mL of Trizol), extracted and precipitated as described by the manufacturer. Precipitated RNAs were solubilized in water with 0.1% (v/v) diethylpyrocarbonate (DEPC). The extracted RNAs were run on 1% agarose gel and visualized by UV transilluminator (Biometra UV star 15).
Transcriptional profiling: Reverse transcription was performed with High Capacity Access RT-PCR System (Promega) using aliquots of total RNA following the manufacturer’s instructions. To generate c-DNA of RA biosynthesis genes, specific primers (Table 1) were supplied by Macrogen Inc. (Korea) according to Kim et al.37. Five genes of PAL, C4H, TAT, 4CL1 and 4CL2 were selected to generate the transcriptional profiling of RA biosynthesis genes.
| Table 1: | Primers of RA biosynthesis genes used in semi-quantitative RT-PCR |
 | |
Semi-quantitative RT-PCR reactions (with a negative control) were performed using PXE 0.5 thermocycler (Thermo Scientific), with the following cycling program: 45 min at 45°C, 2 min at 94°C and 40 cycles of 30 sec at 94°C, 1 min at 60°C, 2 min at 68°C and a 7 min extension at 68°C. The SqRT-PCR products were visualized by agarose gel electrophoresis (1% agarose gel). Quantifications of generated bands were performed with GelPro32 (version 4.03).
Breast cancer cell line culture: The human breast cancer cell line MCF-7 was purchased from American Types culture collection (ATCC, Manassas, VA). The cells were maintained in a Sanyo humidified incubator under 5% CO2, 37°C. The used medium was low glucose Dulbecco's Modified Eagle Medium (Gibco: 21885025) supplemented with 10% fetal bovine serum (FBS, Biochrome), 1% penicillin-streptomycin (5,000 units mL–1, 5000 μL mL–1), respectively and 1% L-glutamine 200 mM. Media was changed every 48 h and the cells were passaged/sub-cultured when reach 75-80% confluency in T-75 culture flask. Media was removed and replaced with 2.5 mL of 0.05% Trypsin EDTA (aUFC Biotech) after washing with 5 mL of 1X PBS (phosphate buffer saline, UFC Biotech). The cells were incubated for 3-5 min. Adding 5 mL of complete medium to deactivate Trypsin EDTA, a cell suspension was formed and then transferred to 15 mL conical tube, centrifuged for 5 min at 1500 rpm min–1. The cell suspension’s supernatant was aspirated and a volume of 1 mL complete medium was used to re-suspend the cells pellet with gentle rapid pipetting. Cells were counted using automated cell counter (Bio-Rad-TC20). Depending on the cell count, cells were seeded either in T-25 culture flask if the cell count was less than 106 or in T-75 if it was 106 or higher. Proper number of seeded cells in the flasks helps establishing cell-to-cell contact and proper cellular growth while maintaining enough space for cell division.
Cell viability assay (MTT): In order to show the crude extract effects on the cell proliferation, a cell viability colorimetric assay of 3-(4,5-Dimethylthiazol-2-yl)-2,5-Diphenyltetrazolium Bromide (MTT) (Invitrogen, USA) was selected to quantify cell number changes over the course of 48 h. The cell seeding density was (1×104 cells/well) in 96-well plate using complete medium. Then cells were incubated for 24 h. On the next day, the cell starvation phase was initiated by using serum free medium for overnight. On the third day, freshly prepared extracts dilutions in phenol red-free and serum-free medium were used to treat the cells and left for 24 and 48 h. The MTT procedure was performed according to the manufacturer. The cells were exposed to the MTT at the 4th (24 h) and 5th day (48 h) to see the effect of plant crude extracts on the cells using an ELISA reader (spectra max plus 384, California, USA) at wavelength of 540 nm and the data were analyzed on computer using (soft max pro 6.2.1, USA) software. The cell viability was determined based on the optical density (OD) detection.
Statistical analysis: Data was analyzed using GraphPad Prism 8 (GraphPad Software, La Jolla, CA, USA). One-way analysis of variance (ANOVA) with Duncan’s multiple range test was used. The p-values less than 0.05 were used to indicate significant differences. Results were expressed as mean±standard deviation (SD).
RESULTS
Growth parameters: The germinated seedlings and callus tissues of thyme (A-B) and hyssop (C-D) was shown in Fig. 1. As for data presented in Fig. 2, thyme plants gave the highest values of fresh weight in tissues of both callus and control seedlings (2.1 and 1.8 g, respectively), whereas values of dry and crude weights in both callus and control seedlings were the same (0.2 and 0.3 g, respectively). Callus fresh and dry weight and crude weight of hyssop were significantly higher than those of thyme (Fig. 2). The results obtained from the thyme callus were superior to the generated seedlings where it gave the highest crude weight value (0.1 g) in which it is important because it contains the rosmarinic acid in connection with the study.
RA content: The standard curve of RA and its content in control seedlings and initiated callus of thyme and hyssop plants is shown in Fig. 3 and 4. As for data presented in Fig. 4, callus tissues significantly exhibited higher RA content compared to control in thyme and hyssop plants. Generally, either callus or seedlings of hyssop accumulated higher levels of RA in their tissues (6.39 and 6.04 mg g–1, respectively), compared to thyme plant (5.67 and 5.32 mg g–1, respectively).
Expression profiles of genes related to RA biosynthesis: To monitor in vitro tissue culture-induced gene expression of RA biosynthesis genes, the gene expression patterns of PAL, C4H, 4CL1, 4CL2 and TAT, which were reported to be associated with RA biosynthesis, were validated by sqRT-PCR in the studied taxa of lamiaceae (H. officinalis and T. vulgaris).
 | |
| Fig. 1(a-d): | Overview of initiated callus and regenerated seedlings, (a) Callus of thyme, (b) Regenerated seedlings of thyme, (c) Callus of hyssop and (d) Regenerated seedlings of hyssop |
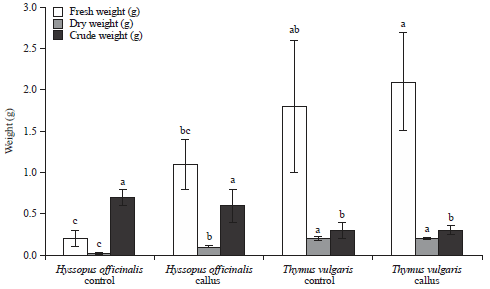 | |
| Fig. 2: | Growth parameters of in vitro generated seedlings and initiated callus of hyssop, H. officinalis and thyme, T. vulgaris plants |
Data presented as Mean±SD (n = 3), bars annotated with the same letter are not significantly (p<0.05) different according Duncan’s multiple range test | |
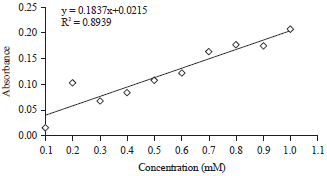 | |
| Fig. 3: | Standard curve of rosmarinic acid appears concentration relative to absorbance |
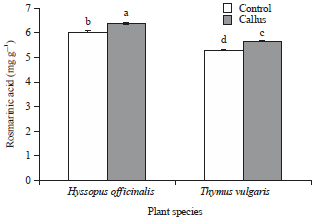 | |
| Fig. 4: | Rosmarinic acid (RA) content in control seedlings and initiated callus of H. officinalis and T. vulgaris plants |
Statistically, one-way analysis of variance (ANOVA) with Duncan’s multiple range test was performed to compare means, error bars represent the standard deviation of the mean (n = 3), letters above bars indicate significant differences between treatments at p<0.05 | |
Tubulin was used as an internal reference for standardization of band intensity transcriptional profile (Fig. 5). Analysis of candidate genes in each plant species pointed out that a single PCR amplicon without any non-specificity was produced and a defined amplification was observed in all in vitro cultured plant tissues. The presented gene expression data analysis in Fig. 5 revealed an evident differential expression pattern of the candidate genes among the 2 studied taxa in both callus and control seedlings (Fig. 5). In hyssop plants, it was noticeable that transcript levels of 4CL1 gene were the highest in both callus and control plants tissues followed by 4CL2, PAL and TAT genes for callus and TAT, PAL and 4CL2 for control, respectively. Synchronously, C4H revealed the lowest transcriptional levels in callus tissues of hyssop plants and also in control seedlings.
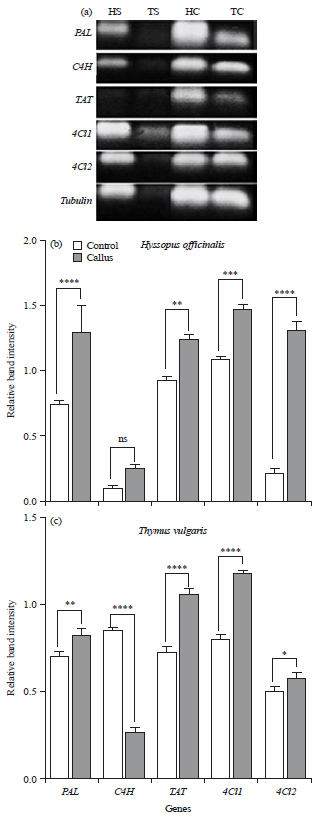 | |
| Fig. 5(a-c): | Gene expression pattern of 5 genes i.e., PAL, C4H, TAT, 4CL1 and 4CL2 in callus tissues (C) compared to control (S) for hyssop and thyme plants |
Tubulin: Housekeeping gene, H: hyssop, T: Thyme, Values are given as Mean±SD, analyzed by one-way analysis of variance (ANOVA), nsp>0.05, *p<0.05, **p<0.01, ***p<0.001, ****p<0.0001 according to Graphpad Prism 8 | |
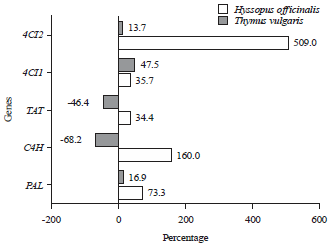 | |
| Fig. 6: | Percent of change in transcriptional levels revealed by H. officinalis and T. vulgaris plants in response to in vitro tissue cultures |
For thyme plants, the same five genes showed a variable pattern of transcriptional levels among the treatments. Even though C4H gene showed the higher percentage of transcripts in control plants tissue, it significantly declined to the less percentage in callus tissues. The PAL and 4CL2 revealed considerable transcriptional levels whether for callus tissue or control plants, whereas, the difference in transcript levels between callus tissue and control plants is less or mild. Collectively, the results of relative band intensities strengthen 4CL2 and PAL as the principal genes in RA biosynthetic pathway among seed embryo plants, where they make up to 64.5% of the total expression. 4CLs isoforms (4CL1 and 4CL2) contributed to the highest percent (39.45%) from which 4CL1 isoform contributed 72.9% of this percent. On the other hand, in callus tissue contribution of 4CLs isoforms of the total gene expression raised up to 47.5% of which 59.3% by 4CL1 and 40.7% by 4CL2. Tissue culture conditions generated a variable pattern of gene expression revealed by the number of transcripts of each gene. Both species respond to in vitro regeneration by significant elevation of PAL, 4CL1 and 4CL2 transcripts. The C4H displayed differential expression pattern, revealed by up regulation in hyssop and down regulation in thyme callus, whereas, TAT was up regulated in callus of hyssop and thyme (Fig. 5).
In hyssop callus, the ranking of transcripts elevation was as follow: 4CL2>C4H>PAL>4CL1>TAT (Fig. 6). In thyme callus, the percent of increase was higher in 4CL1 followed by PAL and 4CL2, whereas, the percent of reduction in transcript numbers of C4H were higher (-68%) relative to TAT gene (-46.4%). Comparing the sensitivity of plant species to tissue culture conditions, transcriptional profile for all genes exhibited that higher percent of change was recorded by hyssop plants (Fig. 6).
Effect of crude extract on cancer cell viability: To define the overall anti-tumor effects of the crude extracts containing RA on breast cancer cell line MCF-7, an analysis of the cell viability was carried out by MTT assay. The results showed that after treatment of human breast cancer cells MCF-7 with the seed embryo and callus crude extracts, T. vulgaris callus extracts did not show any significant (p>0.05) reduction of the cell viability after 24 h of exposure to treatment compared to the untreated control (Fig. 7a), however at 48 h, a statistical significance was detected in the concentration of (90 μg mL–1) with p<0.01 and a less significance was seen with the concentrations of (30, 150 and 210 μg mL–1) with p<0.05 (Fig. 7b). Thymus vulgaris seed embryo extract had no statistical significance (p>0.05) at 24 h (Fig. 7c). Nevertheless, after 48 h of exposure, a decrease in the cell viability of (90 μg mL–1) concentration with p<0.001 and the (30, 150 μg mL–1) concentrations showed less significance with p<0.01 as exhibited in (Fig. 7d). The H. officinalis callus has no significant effects after 24 h of exposition p>0.05 (Fig. 7e), however, after 48 h of treatment, the concentration (150 μg mL–1) showed a significant (p<0.01) reduction in the cell viability and the concentrations of (210, 90 μg mL–1) also had less significant effect p<0.05 (Fig. 7f). On the other hand, hyssop seed embryo extracts also showed no significant effect after 24 h of treating the MCF-7 cells p>0.05 (Fig. 7g). Evidently, a statistical significance was spotted after 48 h with p<0.001 in the concentrations of (210, 150, 90 and 30 μg mL–1) (Fig. 7h).
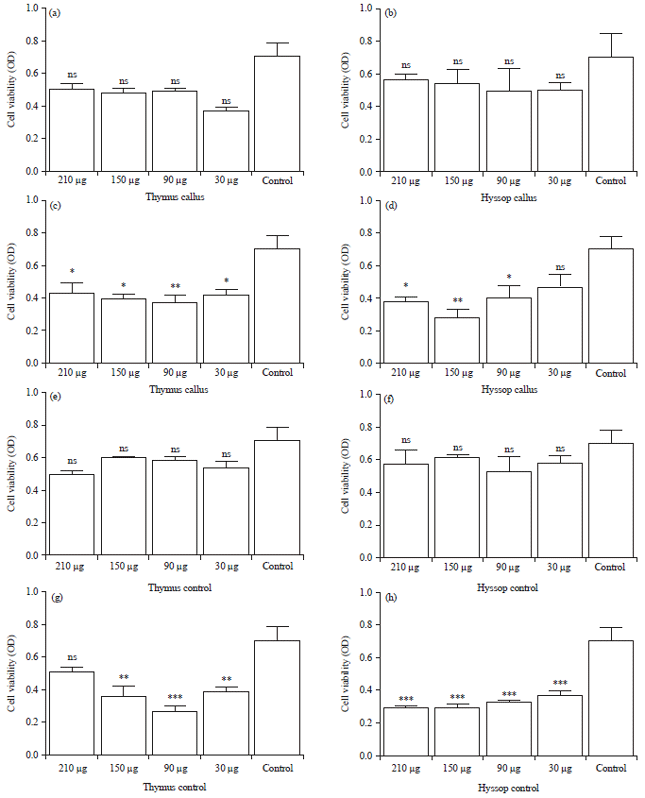 | |
| Fig. 7(a-h): | Concentration-dependent antiproliferative effects of crude extract from thyme and hyssop plants against MCF-7 cells: The effects of different extracts derived from each of the plants that are rich in rosmarinic acid as the extracts were obtained from both culture technology (callus) and seed germinated seedling (control). All the extracts were used to treat cells at different concentrations (30, 90, 150 and 210 μg mL–1), (a, c, e, g) Effects of the T. vulgaris extracts on MCF-7 at 24 and 48 h and (b, d, f, h) Effects of H. officinalis extracts on MCF-7 at 24 and 48 h |
All the results were compared to a negative control (untreated cells), results are expressed as Mean±SD values, analyzed by one-way analysis of variance (ANOVA) by (GraphPad Prism, Version 8.0.1, USA), (nsp>0.05, *p<0.05, ** p<0.01, ***p<0.001 vs. Control), OD: Optical density | |
DISCUSSION
In vitro plant cultures have been widely used, particularly for the production of high-value specialized metabolites38-41. Rosmarinic acid is considered the most important one of those metabolites that has shown many remarkable pharmacological activities42. This promising compound can be produced to around 20% of the dry biomass by using plant cell factories43. Meanwhile, RA is widespread in the plant kingdom and has been found not only in higher plants but also in lower plants. However, there is an inconsistency since not all members in each level of plant taxa contain RA24. Biotechnological approach was then taken in this work to reveal a small part of such phenomena by using 2 genera of lamiaceae. The presence of general phenylpropanoid pathway genes as well as more specialized RA biosynthesis genes has been described in this family3.
In this study, tissue cultures were chosen since the undifferentiated cells from various producing plants have been suggested as a suitable way for RA production44. In a previous investigation, higher amounts of RA have been reported in undifferentiated tissue-cultured cells of Coleus blumei than in the wild plants45. In this study, an increase of RA content up to 6.5% in callus tissue occurred compared to seed plants. In the same context, a similar pattern was revealed by gene expression analysis as transcriptional profile of the selected RA genes in callus tissues indicated significant effects on the RA content in both genotypes. The PAL and 4CLs activities increased dramatically in hyssop and thyme plants, whereas TAT and C4H activity exhibited inconsistent response to in vitro regeneration depending on the plant species. The expression levels of PAL, C4H and 4CL have been verified to encompass in the secondary metabolites synthesis46,45. Thus, in vitro regeneration process might be able to promote the RA biosynthetic pathway, thereby increase in rosmarinic acid production in callus tissue47. Concurrently, the expression levels of PAL, 4CLs, TAT and C4H and the amount of RA accumulated in the in vitro regenerated tissues of each genotype were almost parallel. This is evident from the simultaneous elevation of transcript levels of all genes and RA content in in vitro regenerated hyssop plant compared to thyme. In callus tissues, consistency of RA gene expression with its content was slightly different from the intact plant tissues. RA content increased as 4CL1-2 and PAL expression levels increased in thyme and hyssop, whereas lower C4H and TAT expression levels were associated with higher RA content in thyme callus. These findings also supported the correlation of PAL, TAT, C4H and 4CLs activities to RA synthesis and accumulation in tissue cultures of the studied taxa48. The detected inconsistencies between gene expression and RA production could be ascribed to tissue-specific regulatory programs for individual genes49. Since RA biosynthetic genes are generally shared by biosynthesis of many different secondary metabolites, thus, specific tissue regulation may shift the reaction to the production of other phenolic compounds or lignin as opposed to RA production50,51. Overall, results exhibited variation of the gene transcript levels, contradiction and genotype-specificity. A comparative analysis between the intact plant and callus tissues revealed no exact pattern for the expression of PAL, 4CLs, TAT and C4H. They are found to be inconsistently down-and/or upregulated in each genotype in response to culture conditions. The PAL and 4CLs showed consistent upregulation in both studied species, whereas, C4H and TAT are the only genes in the RA pathway whose expression is downregulated in thyme. To monitor the response to callus conditions, the mean values of transcriptional change percent overall RA genes in each plant, could be used to compare the sensitivity of the tested plants. It might be concluded that the most responsive genus is Hyssopus as it displayed the highest percent of increase in all genes transcripts, whereas Thymus could be suggested as more tolerant to culture conditions based on its lowest percent of change transcripts. For RA genes, present findings support 4CL2 and PAL as the principal genes in RA biosynthetic pathway among seed embryo plants. Moreover, 4CL1 isoform role was verified by its higher percent of contribution among 4Cls. This is in concurrent with the expression of 4CLs genes in lignifying tissues, where 4CL1 alone has been reported to contribute to 90% of the total activity52. Meanwhile, 4CLs1-2 transcript levels in callus tissues of both taxa were up-regulated up to 6-fold, indicating the promotive effect of tissue culture conditions in enhancing the 4CLs1-2 gene expression53. Expression of PAL gene was also elevated up to 1.7-fold compared to seed embryo plants. This result is in consistent with previous reports which found that the expression of PAL, reached to higher levels in undifferentiated cells54,55. On contrary, C4H and TAT were found to be inconsistently down-and/or up regulated in each genotype in response to culture conditions. This support that PAL-derived pathway is more important than TAT-derived pathway in RA biosynthesis in callus tissues of the studied Lamiaceae species.
To our knowledge this is the first study that investigates and compares antiproliferative activity of the RA produced in crude extracted from callus and seed embryo of different plants from Lamiaceae family. The viability decrease was influenced by the amount of concentration and time of exposure. The obtained results showed that T. vulgaris and H. officinalis callus extracts indeed had exhibited reduction in the cell viability with the used concentrations, where after 48 h of treatment, H. officinalis callus extract was more effective at the concentration of 150 μg mL–1, while in T. vulgaris callus, most effective concentration was 90 μg mL–1. The control extracts of H. officinalis and T. vulgaris did not show effects after 24 h, however, after 48 h of exposition the H. officinalis control extracts showed a very high antiproliferative activity in all the used concentrations. These findings are in line with previous studies on T. vulgaris and H. officinalis extracts effects. A recent study by Nile et al.56 on the cytotoxic effects of 50 μL of H. officinalis aerial extracts on human breast cancer cells MCF-7, their results indicated a high antitumor of the extracts against the cells. As for the antitumor and cytotoxic effects of T. vulgaris on cell lines, a study by Abaza et al.57 indicated that after 6-24 h of exposure to this extract, a cytotoxic effects were observed in a dose-dependent manners. In a study by Oliviero et al.58 on the cytotoxic effects of T. vulgaris extracts on human lung cancer cell line H460, their results showed that thyme extracts were able to reduce the cell viability after 72 h of treatment. Results of a recent study indicated that using 10 μL mL–1 of T. vulgaris leaves ethanolic extracts had a significant cell cycle arrest effects leading to apoptosis59.
CONCLUSION
Conclusively, results demonstrated that in vitro culture technique could be useful to stimulate the accumulation of RA in lamiaceae, as it enhances the expression of most of the candidate genes involved in the phenylpropanoid pathway. Overall the present study suggested that both hyssop and thyme crude extracts containing RA as a critical component in the callus or seed-embryo plantlets hold a great antiproliferative activity against human breast cancer cell line MCF-7.
SIGNIFICANCE STATEMENT
This study confirmed that in vitro grown callus tissue is better in its content of Rosmarinic acid. The gene expression of some of the genes in this study reflects this result. The crude methanolic extract of callus tissues has also been shown to be highly effective on the cancer cells compared to the normal plants.
ACKNOWLEDGMENTS
The authors are appreciated to leaders of Taif University in particular, the Deanship of Scientific Research and Head of Biotechnology Department for their support and help to get the study out satisfactorily. This study is the outcome of a research project sponsored by Taif University (Project No. 1-438-5816).
REFERENCES
- Darvishi, E., D. Kahrizi, S. Bahraminejad and M. Mansouri, 2016. In vitro induction of α-pinene, pulegone, menthol, menthone and limonene in cell suspension culture of pennyroyal (Mentha pulegium). Cell. Mol. Biol., 62: 7-9.
CrossRefPubMedDirect Link - Daniell, H., S.J. Streatfield and K. Wycoff, 2001. Medical molecular farming: Production of antibodies, biopharmaceuticals and edible vaccines in plants. Trends Plant Sci., 6: 219-226.
CrossRefPubMedDirect Link - Efferth, T., 2019. Biotechnology applications of plant callus cultures. Engineering, 5: 50-59.
CrossRefDirect Link - Kumar, J. and P.K. Gupta, 2008. Molecular approaches for improvement of medicinal and aromatic plants. Plant Biotechnol. Rep., 2: 93-112.
CrossRefDirect Link - Mulabagal, V. and H.S. Tsay, 2004. Plant cell cultures-an alternative and efficient source for the production of biologically important secondary metabolites. Int. J. Applied Sci. Eng., 2: 29-48.
Direct Link - Taravat, E., A. Zebarjadi, D. Kahrizi and K. Yari, 2015. Isolation, cloning and characterization of a partial novel aro A gene in common reed (Phragmites australis). Pharmaceut. Biol., 53: 637-641.
CrossRefDirect Link - Ma, X.H., Y. Ma, J.F. Tang, Y.L. He and Y.C. Liu et al., 2015. The biosynthetic pathways of tanshinones and phenolic acids in Salvia miltiorrhiza. Molecules, 20: 16235-16254.
CrossRefDirect Link - Zhao, S., P.A. Tuan, X. Li, Y.B. Kim and H.R. Kim et al., 2013. Identification of phenylpropanoid biosynthetic genes and phenylpropanoid accumulation by transcriptome analysis of Lycium chinense. BMC Genomics, Vol. 14.
CrossRefDirect Link - Chabannes, M., K. Ruel, A. Yoshinaga, B. Chabbert, A. Jauneau, J.P. Joseleau and A.M. Boudet, 2001. In situ analysis of lignins in transgenic tobacco reveals a differential impact of individual transformations on the spatial patterns of lignin deposition at the cellular and subcellular levels. Plant J., 28: 271-282.
CrossRefDirect Link - Xie, Y., D. Xu, W. Cui and W. Shen, 2012. Mutation of Arabidopsis HY1 causes UV-C hypersensitivity by impairing carotenoid and flavonoid biosynthesis and the down-regulation of antioxidant defence. J. Exp. Bot., 63: 3869-3883.
CrossRefDirect Link - Widyarini, S., M. Allanson, N.L. Gallagher, J. Pedley and G.M. Boyle et al., 2006. Isoflavonoid photoprotection in mouse and human skin is dependent on metallothionein. J. Invest. Dermatol., 126: 198-204.
CrossRefDirect Link - Parmar, N. and K.R. Dadarwal, 1999. Stimulation of nitrogen fixation and induction of flavonoid-like compounds by rhizobacteria. J. Applied Microbiol., 86: 36-44.
CrossRefDirect Link - Kataoka, I. and K. Beppu, 2004. UV irradiance increases development of red skin color and anthocyanins in 'Hakuho' peach. HortScience, 39: 1234-1237.
CrossRefDirect Link - Hemaiswarya, S. and M. Doble, 2013. Combination of phenylpropanoids with 5-fluorouracil as anti-cancer agents against human cervical cancer (HeLa) cell line. Phytomedicine, 20: 151-158.
CrossRefDirect Link - Alotaibi, R.M., H.R. Rezk, C.I. Juliana and C. Guure, 2018. Breast cancer mortality in Saudi Arabia: Modelling observed and unobserved factors. PLoS ONE, Vol. 13.
CrossRefDirect Link - Kabel, A.M., 2017. Tumor markers of breast cancer: New prospectives. J. Oncological Sci., 3: 5-11.
CrossRefDirect Link - Madkhali, N.A., O. Santin, H. Noble and J. Reid, 2017. Understanding breast health awareness in the Kingdom of Saudi Arabia: A systematic review of the literature. World J. Nurs. Sci., 3: 19-32.
Direct Link - Hsieh, Y.J., H.S. Huang, Y.L. Leu, K.C. Peng, C.J. Chang and M.Y. Chang, 2016. Anticancer activity of Kalanchoe tubiflora extract against human lung cancer cells in vitro and in vivo. Environ. Toxicol., 31: 1663-1673.
CrossRefDirect Link - Nikolaou, M., A. Pavlopoulou, A.G. Georgakilas and E. Kyrodimos, 2018. The challenge of drug resistance in cancer treatment: A current overview. Clin. Exp. Metastasis, 35: 309-318.
CrossRefDirect Link - Iswandana, R., B.T. Pham, W.T. van Haaften, T. Luangmonkong, D. Oosterhuis, H.A.M. Mutsaers and P. Olinga, 2016. Organ- and species-specific biological activity of rosmarinic acid. Toxicol. In vitro, 32: 261-268.
CrossRefDirect Link - Wu, C.F., A. Karioti, D. Rohr, A.R. Bilia and T. Efferth, 2016. Production of rosmarinic acid and salvianolic acid B from callus culture of Salvia miltiorrhiza with cytotoxicity towards acute lymphoblastic leukemia cells. Food Chem., 201: 292-297.
CrossRefDirect Link - Kim, K.H., V. Janiak and M. Petersen, 2004. Purification, cloning and functional expression of hydroxyphenylpyruvate reductase involved in rosmarinic acid biosynthesis in cell cultures of Coleus blumei. Plant Mol. Biol., 54: 311-323.
CrossRefDirect Link - Trocsanyi, E., Z. Gyorgy, K. Inotai, K. Szabo and Z. Pluhar et al., 2015. Enhanced rosmarinic acid accumulation and rosmarinic acid synthase gene expression under drought stress in thyme (Thymus vulgaris). Planta Medica, Vol. 81, No. 16.
CrossRefDirect Link - Petersen, M., 1997. Cytochrome P450-dependent hydroxylation in the biosynthesis of rosmarinic acid in Coleus. Phytochemistry, 45: 1165-1172.
CrossRefDirect Link - Ellis, B. and G.H.N. Towers, 1970. Biogenesis of rosmarinic acid in Mentha. Biochem. J., 118: 291-297.
CrossRefDirect Link - Venkateshappa, S.M. and K.P. Sreenath, 2013. Potential medicinal plants of Lamiaceae. Am. Int. J. Res. Formal Applied Nat. Sci., 3: 82-87.
Direct Link - Tuan, P.A., W.T. Park, H. Xu, N.I. Park and S.U. Park, 2012. Accumulation of tilianin and rosmarinic acid and expression of phenylpropanoid biosynthetic genes in Agastache rugosa. J. Agric. Food Chem., 60: 5945-5951.
CrossRefDirect Link - Lu, X., L. Hao, F. Wang, C. Huang and S. Wu, 2013. Molecular cloning and overexpression of the tyrosine aminotransferase (TAT) gene leads to increased rosmarinic acid yield in Perilla frutescens. Plant Cell Tissue Org. Cult., 115: 69-83.
CrossRefDirect Link - Weitzel, C. and M. Petersen, 2011. Cloning and characterisation of rosmarinic acid synthase from Melissa officinalis L. Phytochemistry, 72: 572-578.
CrossRefDirect Link - Song, J. and Z. Wang, 2011. RNAi-mediated suppression of the phenylalanine ammonia-lyase gene in Salvia miltiorrhiza causes abnormal phenotypes and a reduction in rosmarinic acid biosynthesis. J. Plant Res., 124: 183-192.
CrossRefDirect Link - Murashige, T. and F. Skoog, 1962. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant., 15: 473-497.
CrossRefDirect Link - Lopez-Arnaldos, T., M. Lopez-Serrano, A.R. Barcelo, A.A. Calderon and J.M. Zapata, 1995. Spectrophotometric determination of rosmarinic acid in plant cell cultures by complexation with Fe2+ ions. Fresenius' J. Anal. Chem., 351: 311-314.
CrossRefDirect Link - Chomczynski, P., 1993. A reagent for the single-step simultaneous isolation of RNA, DNA and proteins from cell and tissue samples. Biotechniques, 15: 532-537.
PubMed - Kim, Y.B., Y. Shin, P.A. Tuan, X. Li, Y. Park, N.I. Park and S.U. Park, 2014. Molecular cloning and characterization of genes involved in rosmarinic acid biosynthesis from Prunella vulgaris. Biol. Pharmaceut. Bull., 37: 1221-1227.
CrossRefDirect Link - Verpoorte, R., R. van der Heijden and J. Memelink, 2000. Engineering the plant cell factory for secondary metabolite production. Transgenic Res., 9: 323-343.
CrossRefDirect Link - Verpoorte, R., A. Contin and J. Memelink, 2002. Biotechnology for the production of plant secondary metabolites. Phytochem. Rev., 1: 13-25.
CrossRefDirect Link - Rao, S.R. and G.A. Ravishankar, 2002. Plant cell cultures: Chemical factories of secondary metabolites. Biotechnol. Adv., 20: 101-153.
CrossRefPubMedDirect Link - Matkowski, A., 2008. Plant in vitro culture for the production of antioxidants—A review. Biotechnol. Adv., 26: 548-560.
CrossRefDirect Link - Amoah, S.K., L.P. Sandjo, J.M. Kratz and M.W. Biavatti, 2016. Rosmarinic acid-pharmaceutical and clinical aspects. Planta Medica, 82: 388-406.
CrossRefDirect Link - Doring, A.S. and M. Petersen, 2014. Production of caffeic, chlorogenic and rosmarinic acids in plants and suspension cultures of Glechoma hederacea. Phytochem. Lett., 10: 111-118.
CrossRefDirect Link - Bulgakov, V.P., Y.V. Inyushkina and S.A. Fedoreyev, 2012. Rosmarinic acid and its derivatives: Biotechnology and applications. Crit. Rev. Biotechnol., 32: 203-217.
CrossRefDirect Link - Park, W.T., M.V. Arasu, N.A. Al-Dhabi, S.K. Yeo and J. Jeon et al., 2016. Yeast extract and silver nitrate induce the expression of phenylpropanoid biosynthetic genes and induce the accumulation of rosmarinic acid in Agastache rugosa cell culture. Molecules, Vol. 21, No. 4.
CrossRefDirect Link - Kim, Y.B., J.K. Kim, M.R. Uddin, H. Xu and W.T. Park et al., 2013. Metabolomics analysis and biosynthesis of rosmarinic acid in Agastache rugosa Kuntze treated with methyl jasmonate. PLoS ONE, Vol. 8, No. 5.
CrossRefDirect Link - Mizukami, H., Y. Tabira and B. Ellis, 1993. Methyl jasmonate-induced rosmarinic acid biosynthesis in Lithospermum erythrorhizon cell suspension cultures. Plant Cell Rep., 29: 706-709.
CrossRefDirect Link - Karwatzki, B., M. Petersen and A.W. Alfermann, 1989. Transient activity of enzymes involved in the biosynthesis of rosmarinic acid in cell suspension cultures of Coleus blumei. Planta Medica, 55: 663-664.
Direct Link - El-Naggar, H.M. and P.E. Read, 2010. PAL gene activity and rosmarinic acid production in rosemary genotypes. J. Herbs Spices Med. Plants, 16: 83-89.
CrossRefDirect Link - Yang, R. and K. Shetty, 1998. Stimulation of rosmarinic acid in shoot cultures of oregano (Origanum vulgare) clonal line in response to proline, proline analogue and proline precursors. J. Agric. Food Chem., 46: 2888-2893.
CrossRefDirect Link - Li, H., Y. Fu, H. Sun, Y. Zhang and X. Lan, 2017. Transcriptomic analyses reveal biosynthetic genes related to rosmarinic acid in Dracocephalum tanguticum. Scient. Rep., Vol. 7, No. 1.
CrossRefDirect Link - Li, Y., J.I. Kim, L. Pysh and C. Chapple, 2015. Four isoforms of Arabidopsis 4-coumarate: CoA ligase have overlapping yet distinct roles in phenylpropanoid metabolism. Plant Physiol., 169: 2409-2421.
CrossRefDirect Link - Ata, N., N.A. Yusuf, B.C. Tan, A. Husaini, Y.M. Yusuf, N.A. Majid and N. Khalid, 2015. Expression profiles of flavonoid-related gene, 4 coumarate: Coenzyme A ligase and optimization of culturing conditions for the selected flavonoid production in Boesenbergia rotunda. Plant Cell Tissue Org. Cult., 123: 47-55.
CrossRefDirect Link - Raes, J., A. Rohde, J.H. Christensen, Y. van de Peer and W. Boerjan, 2003. Genome-wide characterization of the lignification toolbox in Arabidopsis. Plant Physiol., 133: 1051-1071.
CrossRefDirect Link - Rohde, A., K. Morreel, J. Ralph, G. Goeminne and V. Hostyn et al., 2004. Molecular phenotyping of the pal1 and pal2 mutants of Arabidopsis thaliana reveals far-reaching consequences on phenylpropanoid, amino acid and carbohydrate metabolism. Plant Cell, 16: 2749-2771.
CrossRefDirect Link - Nile, S.H., A.S. Nile and Y.S. Keum, 2017. Total phenolics, antioxidant, antitumor and enzyme inhibitory activity of Indian medicinal and aromatic plants extracted with different extraction methods. 3 Biotech, Vol. 7, No. 1.
CrossRefDirect Link - Abaza, M.S.I., K.Y. Orabi, E. Al-Quattan and R.J. Al-Attiyah, 2015. Growth inhibitory and chemo-sensitization effects of naringenin, a natural flavanone purified from Thymus vulgaris, on human breast and colorectal cancer. Cancer Cell Int., Vol. 15.
CrossRefDirect Link - Oliviero, M., I. Romilde, M.M. Beatrice, V. Matteo and N. Giovanna et al., 2016. Evaluations of thyme extract effects in human normal bronchial and tracheal epithelial cell lines and in human lung cancer cell line. Chemico-Biol. Interact., 256: 125-133.
CrossRefDirect Link - Al-Seragy, I.M., K.R. Kharat and A.S. Dhabe, 2019. Cell cycle arrest and induction of apoptosis in human breast cancer cells (T-47D) by Annona squamosa L. and Thymus vulgaris L. ethanolic extract. J. Biol. Act. Prod. Nat., 9: 47-56.
CrossRefDirect Link