
Alaadin El-Sayed El-Haddad
Department of Pharmacognosy, Faculty of Pharmacy, October 6 University, Giza, Egypt
LiveDNA: 20.29230
Amr Mohammed Saadeldeen
Department of Pharmacognosy, Faculty of Pharmacy, October 6 University, Giza, Egypt
Soad Zakaria El-Emam
Department of Pharmacology and Toxicology, Faculty of Pharmacy, October 6 University, Giza, Egypt
Pakistan Journal of Biological Sciences
Year: 2019 | Volume: 22 | Issue: 10 | Page No.: 502-509







ABSTRACT
Background and Objective: The waste products of the tamarind canning industry have been discarded; however, it has potential health benefits. Herein, the study was carried out HPLC profiling of phenolic constituents of Tamarindis indica pericarp and seeds. Furthermore, the cytotoxic activity against HUH-7 cells was evaluated and assessed with molecular docking study on angiogenesis-related VEGF kinase proteins in addition to evaluating the level of released VEGF in treated HUH-7 cells by ELISA. Materials and Methods: Folin-ciocalteu and AlCl3 assays were used for quantification of total phenolics (TPC) and total flavonoids (TFC) contents, respectively. Molecular docking study was done on VEGF kinase proteins. Results: TPC and TFC of pericarp and seeds were 0.35±0.02 g GAE g–1 DE and 0.12±0.009 g CE g–1 DE, 0.39±0.01 g GAE g–1 DE and 0.03±0.006 g CE g–1 DE, respectively. In pericarp, 8 phenolics were tentatively identified, where (+)-catechin was the major (27,386.04 μg g–1 DE) followed by gallic acid and naringenin (931.47, 500.42 μg g–1 DE) respectively. While in seeds, 11 phenolics were tentatively identified, where naringenin was the major (95,305.47 μg g–1 DE) followed by (+)-catechin and rutin (54,930.29, 15,361.66 μg g–1 DE) respectively. Aqueous and methanol seeds extracts exhibit cytotoxic effect with IC50 27.4±1.81 and 13.4±0.94 μg mL–1, respectively, it was more potent than aqueous and methanol pericarp extracts which had IC50 132±5.82 and 61.6±3.16 μg mL–1. The tested phenolics were fit on the active sites of VEGF kinase targets with varied degree of interactions. The cytotoxic and anti-angiogenic activities were confirmed in light of phenolics docking interactions. Conclusion: Results demonstrate for the first time that phenolics could inhibit angiogenesis via inhibiting kinase proteins, which could therefore be developed as antiangiogenic drugs.
PDF Abstract XML References Citation
Copyright: © 2019. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Alaadin El-Sayed El-Haddad, Amr Mohammed Saadeldeen and Soad Zakaria El-Emam, 2019. Anti-angiogenic Activity of Major Phenolics in Tamarind Assessed with Molecular Docking Study on VEGF Kinase Proteins. Pakistan Journal of Biological Sciences, 22: 502-509.
DOI: 10.3923/pjbs.2019.502.509
URL: https://scialert.net/abstract/?doi=pjbs.2019.502.509
DOI: 10.3923/pjbs.2019.502.509
URL: https://scialert.net/abstract/?doi=pjbs.2019.502.509
INTRODUCTION
The anticancer agents should target multiple pathways that involved in carcinogenesis. Angiogenesis is a necessary process in tumor growth and metastasis. The angiogenic factor, vascular endothelial growth factor (VEGF), is a cell signaling protein that initiates vasculogenesis (formation of new blood vessels) and its binding to the receptors (VEGFRs) stimulates endothelial cell survival, proliferation and migration1. A new tactic of anticancer drugs by inhibition of angiogenesis have been identified. VEGF signaling pathway has been considered as a promising target in controlling cancer progression2. Recently, inhibitors against VEGF pathway (e.g., sunitinib) that are released in the market elicited a range of adverse effects3. Thus, devising safe new anti-angiogenic drugs with selected anti-tumor targets is necessary. Dietary phytochemicals as chemopreventive agents are expected to be less toxic. Natural polyphenolics; catechin, naringenin and procyanidin are well-known to exhibit beneficial health effects including anti-inflammatory, anti-proliferative and antitumor activities. Procyanidin has numerous targets of intracellular signaling pathways, including pro-inflammatory mediators of cell survival, apoptosis and angiogenesis4. Currently, many reports suggest procyanidin as a promising compound for cancer prevention and treatment.
Tamarind (Tamarindis indica L., Caesalpinioideae) is widely consumed fresh or as juice and jams. Antioxidant, antihyperlipidemic5 and antimicrobial6 activities were reported for its fruits. The fruit extract was more active than that of the seeds, with greater ability to scavenge radicals and inhibit xanthine oxidase7. Phenolics and flavonoids viz; catechin, epicatechin and taxifolin were isolated from the fruits extract7. Tamarind seeds were reported to contain mainly procyanidin oligomers (up to eleven)7. Flavonoids can reduce lipid oxidation by lipoxygenase inhibition or by radicals scavenging8. Naringenin has been demonstrated to exhibit antioxidant and antiproliferative effects9.
A large amount of tamarind seeds wastes have been discarded from the industry. Tamarind seeds are a low-cost rich source of antioxidants phytochemicals such as catechin and naringenin10,11. The study’s goal was to highlight the potential health benefits of underutilized by-product. Herein, quantification and HPLC profiling of phenolic constituents of T. indica pericarp and seed extracts have proceed. Furthermore, in continued search for cytotoxic phytochemicals12, the cytotoxic activity was evaluated and assessed by molecular docking of major occurring phenolics in tamarind on VEGF kinase proteins.
MATERIALS AND METHODS
The study was proceed from May, 2017 till June, 2019 in Faculty of Pharmacy, October 6 University and HPLC profiling was done on National Research Centre, Cairo, Egypt. Gallic acid, (+)-catechin, Folin-Ciocalteu’s reagent, MTT (5 mg mL–1) in 0.9% NaCl solubilized in acidified isopropanol (0.04 N HCl in isopropanol) (Sigma-Aldrich Co., Germany). Authentic phenolics for HPLC profiling were kindly supplied by Food Technology Research Institute, Giza, Egypt. All other chemicals were of the highest purity.
Plant material and extraction process: Pods of T. indica L. were purchased from Egypt’s local market. The identity was kindly confirmed by Agriculture Research Center, Cairo, Egypt. Seeds were carefully separated from pericarp and were powdered separately. Powdered pericarp and seeds (100 g each) were separately defatted with n-hexane (3×1 L) then extracted in Soxhlet with methanol (99.5%, 500 mL, 30 min). In a parallel manner, another powdered pericarp and seeds (100 g each) were separately defatted and extracted in Soxhlet with water (500 mL, 30 min). Extracts (methanol and aqueous) were left to cool, filtered then were evaporated individually (Rotavapor® R-300, BÜCHI, Switzerland)13.
Total phenolics and flavonoids contents: Total phenolics content (TPC) as gallic acid equivalent of dried extract (GAE g–1 DE) and total flavonoids content (TFC) as catechin equivalent (CE g–1 DE) were carried out using folin-ciocalteu and AlCl3 assays, respectively. Dried powdered T. indica pericarp and seeds (1 g each) were separately defatted with n-hexane (10 mL, twice) followed by extraction by sonication with methanol (95%, 50 mL). Methanol was distilled and extracts volume were adjusted to 100 mL with distilled water for phenolics determination or with ethanol for flavonoids determination. By measuring the color intensity using spectrophotometer (Shimadzu Corporation, Japan), phenolics and flavonoids were complexed with folin-ciocalteu’s phenol and ALCl3, respectively, with reference to pre-established standard calibration curves14,15.
HPLC profiles of phenolics: Methanol extracts (1 g each) were separately sonicated with aqueous acetone (70%, 200 mL). After acetone removing, the residues were sonicated in methanol (10 mL, 5 min) then centrifuged (1000 rpm, 10 min). The supernatants were filtered (0.2 millipore membrane filter). Identification was performed using Hewlett Packard HPLC system (series 1050) equipped with a Lichrosorb RP-18 column (4.0×250 mm; 5 μm) and UV detector (280 and 330 nm for phenolics and flavonoids, respectively). Elution was carried out using methanol and acetonitrile (2:1, 1 mL min–1). Peaks assignments were confirmed by the injection of authentics. The retention time and peak area were used to calculate compound concentrations by Hewlett Packard software16,17.
In vitro cytotoxic activity and evaluating the level of VEGF release: Cytotoxic activity was valued by MTT cell proliferation assay18. In 96 well-microplates, human liver carcinoma (HUH-7) cells (0.5×105) were cultured. Cells were treated with tamarind extracts individually dissolved in DMSO (20 μL, 100-0.8 μg mL–1). After incubation of plates, the media were removed and MTT solution (40 μL/well) was added and the plates were shaken at room temperature (4 h). Absorbance was photometrically determined at 570 nm using ELISA reader (FLUOstar Omega, BMG, Labtech, Germany). The relative viability percentage was calculated by18:
![]()
For VEGF quantification, HUH-7 cells were cultured in RPMI media, treated with tamarind extracts (100 μg mL–1) and incubated (12 h). Briefly, after incubation, media and cells were collected and centrifuged. The cell lysate was prepared and passed through a 21-gauge needle to break up cell aggregates, then centrifuged (14,000 rpm, 15 min, 4°C). The supernatants were used for quantifying VEGF by Sandwich ELISA using a VEGF ELISA kit according to the manufacturer’s instructions (Cusabio Technology LLC)19. Data (as Mean±SD) were statistically analyzed by GraphPad Prism 6 (La Jolla, CA, USA), using one-way ANOVA followed by Tukey's Kramer Multiple Comparison Test.
Molecular docking study
Preparation of targets and molecular docking protocols: Molecular Operating Environment (MOE 2014.0901) software was used. The X-ray crystallographic structure of VEGF-1 (PDB ID: 3HNG), VEGF-2 (PDB ID: 2QU6, 2QU5, 2P2I, 3EWH, 1YWN) transferase (kinase) proteins were downloaded (https://www.rcsb.org). The enzymes were prepared by removal of unnecessary chains and/or ligands. Docking setup was validated by redocking of the co-crystallized ligands for predicting the ligand-receptor interactions. The enzymes and examined compounds were then 3D protonated and energy minimized with default options.
| Table 1: | Redocked conformations of co-crystalized ligands with VEGF kinase proteins |
 | |
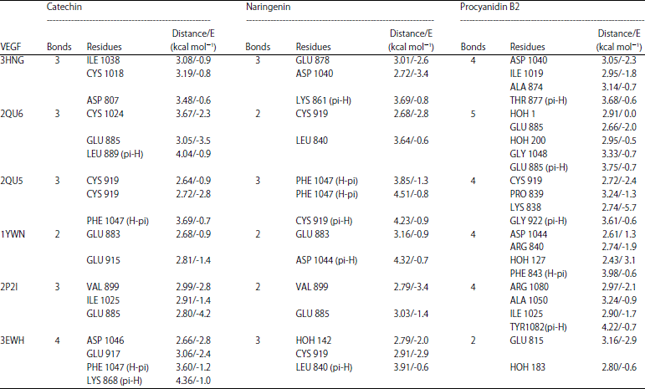
The protocol suitability was assured by low binding energy score (S), small RMSD value and the ability of the redocked pose to reproduce the key interactions accomplished by the co-crystallized ligand, (Table 1). The validated protocol was then used in predicting the ligand-receptor interactions at the binding site for catechin, naringenin and procyanidin B2.
RESULTS
Phenolics and flavonoids contents and HPLC profiling: In pericarp, TPC and TFC were 0.35±0.02 g GAE g–1 DE and 0.12±0.009 g CE g–1 DE, respectively. However those for seeds were 0.39±0.01 g GAE g–1 DE and 0.03±0.006 g CE g–1 DE, respectively (Table 2). This is similar to the levels reported previously8. The TPC differed between pericarp and seeds extracts as always TFC is lower than TPC20.
The methanol extracts obtained from the tamarind pericarp and seeds were analyzed by HPLC-UV. Relative concentrations were determined by peak areas (Fig. 1). In pericarp, eight phenolics were tentatively identified, where (+)-catechin was the major (27,386.04 μg g–1 DE) followed by gallic acid and naringenin (931.47, 500.42 μg g–1 DE) respectively, in addition to syringic acid, propyl gallate, daidzein (4`,7-dihydroxyisoflavone), quercetin and cinnamic acid (Table 3, Fig. 1a). While in seeds, eleven phenolics were tentatively identified, where naringenin was the major (95,305.47 μg g–1 DE) followed by (+)-catechin and rutin (54,930.29, 15,361.66 μg g–1 DE) respectively, in addition to gallic, coffeic, syringic, coumaric acids, vanillin, propyl gallate, quercetin and cinnamic acid (Table 3, Fig. 1b).
In vitro cytotoxic activity and evaluation of VEGF released by HUH-7 cells: Tamarind pericarp and seeds extracts were examined against HUH-7 cells. Aqueous and methanol seeds extracts exhibited cytotoxic effects with IC50 27.4±1.81 and 13.4±0.94 μg mL–1, respectively, it was more potent than those of pericarp extracts (IC50 of 132±5.82 and 61.6±3.16 μg mL–1, respectively) (Table 4). Treatment of HUH-7 cells with tamarind extracts significantly (p<0.05) reduced the level of VEGF when compared with untreated control cells.
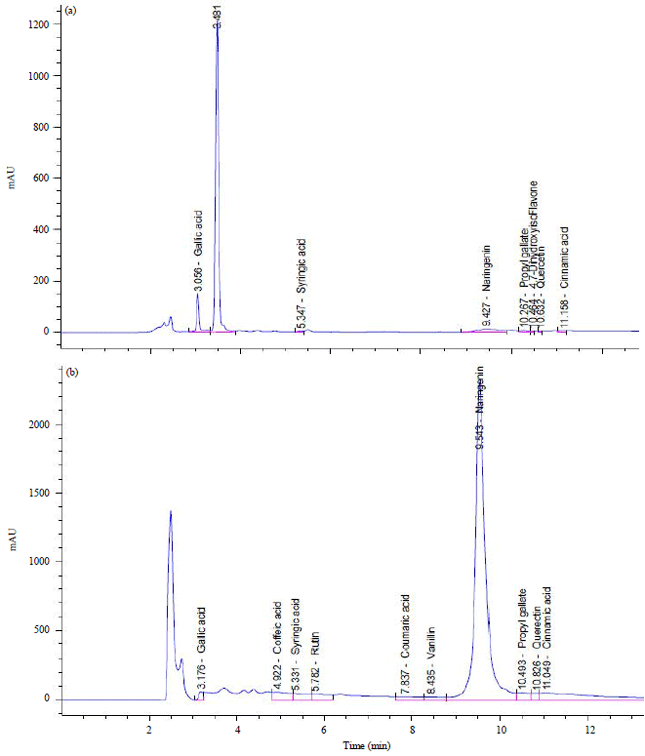 | |
| Fig. 1(a-b): | Analytical HPLC-UV chromatograms of (a) Tamarind pericarp and (b) Seeds methanol extracts |
Moreover, further inhibition was observed with seed extracts especially the methanol extract when compared with pericarp extracts (Fig. 2). Our results are in accordance with several studies showing the significance of phenolics in inhibiting angiogenesis via reducing VEGFR signaling4,21.
Molecular docking of detected phenolics on VEGF kinase proteins: Molecular docking of catechin, naringenin and procyanidin B2 were performed for the first time on VEGF kinase enzyme. Tested phenolics were fitted on the active sites with varied degree of interactions. The active pocket of 3EWH is mainly composed of conserved ASP 1046, GLU 917, PHE 1047 and LYS 868. The main interactions include H-bonds and H-pi interactions between amino acids and ligand.
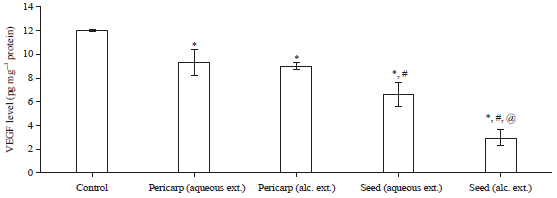 | |
| Fig. 2: | Level of VEGF released by HUH-7 cells after treatment with tamarind pericarp and seeds extracts |
*p<0.05 when compared to untreated control cells, #p<0.05 when compared to pericarp extracts treated cells, @p<0.05 when compared to seed aqueous extract treated cells | |
| Table 2: | Total phenolics (TPC) and total flavonoids (TFC) contents of tamarind pericarp and seeds |
 | |
| Table 3: | Phenolics contents of tamarind pericarp and seeds analyzed by HPLC-UV |
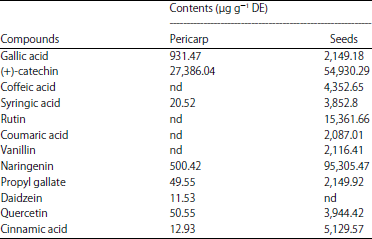 | |
nd: not detected | |
| Table 4: | IC50 of T. indica pericarp and seed extracts against HUH-7 cells |
 | |
Catechin interacts by four H-bonds between three hydroxyl groups and ASP 1046, GLU 917 and PHE 1047 moreover it also interacts by pi-H-bond with LYS 868 (Table 5, Fig. 3). However, in 3HNG, the active pocket is mainly composed of conserved GLU 878, ASP 1040 and LYS 861. Its main key interactions include H-bonds and pi-H-bond interactions. Naringenin interacts by two H-bonds, one is between the hydroxyl group and GLU 878 and the other is between the carbonyl group and ASP 1040, moreover it also interacts by pi-H-bond with LYS 861 (Table 5, Fig. 4). The binding pocket of 2QU6 is mainly composed of conserved GLU 885, GLY 1048, HOH 1 and HOH 200. The main key interactions include H-bonds and pi-H-bond interactions. Procyanidin B2 interacts by two H-bonds between hydroxyl groups and GLU 885 and GLY 1048. It also interacts by pi-H-bond with GLU 885, moreover it interacts through two hydroxyl groups with water-mediated H-bond with HIS 1026 and ILE 1025 (Table 5, Fig. 5). The cytotoxic and anti-angiogenic activities of tamarind extracts were confirmed in light of its phenolics docking interactions.
DISCUSSION
Phenolics and flavonoids are naturally-occurring phytochemicals, exhibited good antioxidant potential. To date, more than 6000 flavonoids have been recognized. Methanol, as a polar solvent, efficiently used in the extraction of phenolics22. RP-HPLC-UV analysis of the extracts revealed the presence of several peaks corresponding to phenolics and flavonoids including naringenin, (+)-catechin, rutin, gallic, coffeic, syringic, coumaric acids, vanillin, propyl gallate, quercetin and cinnamic acid (Table 3). Catechin and naringenin were the majors in pericarp and seeds extracts respectively. Sudjaroen et al.7 reported that the pericarp mainly contains catechin and naringenin, while the seeds are mainly dominated by proanthocyanidins and naringenin and lower levels of catechin.
All of the detected phenolics are known to have biological activities viz; (+)-catechin, has chemoprotective effects by reducing oxidative stress. The detected flavonoids could act synergistically in antioxidant activities. Result of in vitro anti-carcinogenesis assay showed that naringenin inhibited aflatoxin B1-induced carcinogenesis22,23.
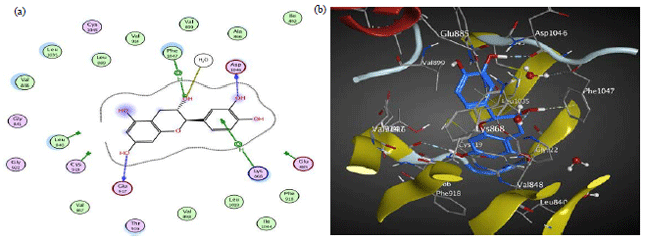 | |
| Fig. 3(a-b): | (a) 2D and (b) 3D diagram showing the binding mode of catechin (blue) with 3EWH VEGF kinase binding pocket |
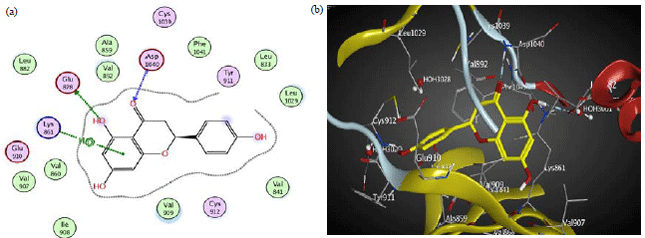 | |
| Fig. 4(a-b): | (a) 2D and (b) 3D diagram showing the binding mode of naringenin (yellow) with 3HNG VEGF kinase binding pocket |
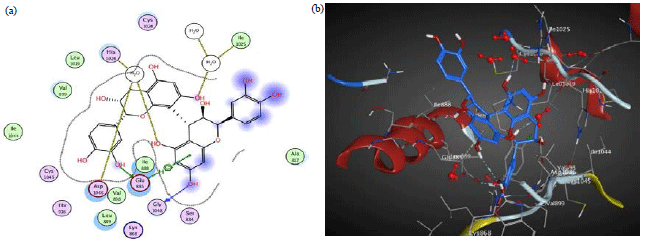 | |
| Fig. 5(a-b): | (a) 2D and (b) 3D diagram showing the binding mode of procyanidin B2 (blue) with 2QU6 VEGF kinase binding pocket |
| Table 5: | Docked conformations of catechin, naringenin and procyanidin B2 with VEGF kinase proteins |
 | |
Naringenin has a potentially useful inhibitory effect on tumor growth24. Recent studies have shown that procyanidin on human breast25, lung26, oral27, prostate cancers28 resulted in an inhibition of cell proliferation without a toxic effect on normal cells29. VEGF promotes angiogenesis via endothelial cells activation. Procyanidin B2 reduced the expression levels of VEGF and has been reported to alter carcinogen metabolism, inflammation, proliferation, apoptosis and angiogenesis30. Thus, VEGF is an ideal anticancer target. The interactions of VEGF-1 and VEGF-2 with catechin, naringenin and procyanidin B2 supported their usage against angiogenic molecular targets. The study demonstrates for the first time that catechin, naringenin and procyanidin B2 are betters in inhibiting angiogenesis via inhibiting VEGF.
CONCLUSION
Tamarind phenolics inhibit cell viability by decreasing angiogenesis. Incorporation of powder of tamarind seeds into the diet significantly increases the bioactive antioxidant and anticancer phytochemicals. Finally; the results may help the scientific community of medicine to carry out lab experiments considering the potential of tamarind seeds.
SIGNIFICANCE STATEMENT
This study highlighted the potential health benefits of the industrial underutilized by-product of T. indica that can be beneficial for consuming as a nutraceutical or food supplement. This study will help the researchers to uncover the critical areas of the anti-angiogenic effect of phenolics that many researchers were not able to explore.
REFERENCES
- Tabernero, J., 2007. The role of VEGF and EGFR inhibition: Implications for combining anti–VEGF and anti–EGFR agents. Mol. Cancer Res., 5: 203-220.
CrossRefDirect Link - Ebos, J.M.L. and R.S. Kerbel, 2011. Antiangiogenic therapy: Impact on invasion, disease progression and metastasis. Nat. Rev. Clin. Oncol., 8: 210-221.
CrossRefDirect Link - Lee, W.J., J.L. Lee, S.E. Chang, M.W. Lee and Y.K. Kang et al., 2009. Cutaneous adverse effects in patients treated with the multitargeted kinase inhibitors sorafenib and sunitinib. Br. J. Dermatol., 161: 1045-1051.
CrossRefDirect Link - Lee, Y., 2017. Cancer chemopreventive potential of procyanidin. Toxicol. Res., 33: 273-282.
CrossRefDirect Link - Martinello, F., S.M. Soares, J.J. Franco, A.C. Santos and A. Sugohara et al., 2006. Hypolipemic and antioxidant activities from Tamarindus indica L. pulp fruit extract in hypercholesterolemic hamsters. Food Chem. Toxicol., 44: 810-818.
CrossRefPubMedDirect Link - Tril, U., J. Fernandez-Lopez, J.A.P. Alvarez and M. Viuda-Martos, 2014. Chemical, physicochemical, technological, antibacterial and antioxidant properties of rich-fibre powder extract obtained from tamarind (Tamarindus indica L.). Ind. Crops Prod., 55: 155-162.
CrossRefDirect Link - Sudjaroen, Y., R. Haubner, G. Wurtele, W.E. Hull and G. Erben et al., 2005. Isolation and structure elucidation of phenolic antioxidants from Tamarind (Tamarindus indica L.) seeds and pericarp. Food Chem. Toxicol., 43: 1673-1682.
CrossRefDirect Link - Maisuthisakul, P., M. Suttajit and R. Pongsawatmanit, 2007. Assessment of phenolic content and free radical-scavenging capacity of some thai indigenous plants. Food Chem., 100: 1409-1418.
CrossRefDirect Link - Galati, G., M.Y. Moridani, T.S. Chan and P.J. O'Brien, 2001. Peroxidative metabolism of apigenin and naringenin versus luteolin and quercetin: Glutathione oxidation and conjugation. Free Radical Biol. Med., 30: 370-382.
CrossRefDirect Link - Tsuda, T., M. Watanabe, K. Ohshima, A. Yamamoto, S. Kawakishi and T. Osawa, 1994. Antioxidative components isolated from the seed of tamarind (Tamarindus indica L.). J. Agric. Food Chem., 42: 2671-2674.
CrossRefDirect Link - Natukunda, S., J.H. Muyonga and I.M. Mukisa, 2016. Effect of tamarind (Tamarindus indica L.) seed on antioxidant activity, phytocompounds, physicochemical characteristics and sensory acceptability of enriched cookies and mango juice. Food Sci. Nutr., 4: 494-507.
CrossRefDirect Link - Emam, M., M.A. El Raey, W.H. Eisa, A.E. El-Haddad, S.M. Osman, M.A. El-Ansari and A.G.M. Rabie, 2017. Green synthesis of silver nanoparticles from Caesalpinia gilliesii (Hook) leaves: Antimicrobial activity and in vitro cytotoxic effect against BJ-1 and MCF-7 cells. J. Applied Pharm. Sci., 7: 226-233.
CrossRefDirect Link - Boshra, S.A. and A.E. El-Haddad, 2018. The protective effects of MPLC isolated Glycyrrhizin and mangiferin against brain toxicity in rats. Med. Chem. Res., 27: 1449-1459.
CrossRefDirect Link - Dewanto, V., X. Wu, K.K. Adom and R.H. Liu, 2002. Thermal processing enhances the nutritional value of tomatoes by increasing total antioxidant activity. J. Agric. Food Chem., 50: 3010-3014.
CrossRefDirect Link - El-Gizawy, H.A., A.S. Alazzouni and A.E. El-Haddad, 2018. Pharmacognostical and biological studies of Delonix regia growing in Egypt: HPLC profiles. Pharmacogn. Commun., 8: 125-131.
Direct Link - Goupy, P., M. Hugues, P. Boivin and M.J. Amiot, 1999. Antioxidant composition and activity of barley (Hordeum vulgare) and malt extracts and of isolated phenolic compounds. J. Agric. Food Chem., 79: 1625-1634.
CrossRefDirect Link - Mattila, P., J. Astola and J. Kumpulainen, 2000. Determination of flavonoids in plant material by HPLC with diode-array and electro-array detections. J. Agric. Food Chem., 48: 5834-5841.
CrossRefPubMedDirect Link - Mosmann, T., 1983. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods, 65: 55-63.
CrossRefPubMedDirect Link - Hornig, C., T. Behn, W. Bartsch, A. Yayon and H.A. Weich, 1999. Detection and quantification of complexed and free soluble human vascular endothelial growth factor receptor-1 (sVEGFR-1) by ELISA. J. Immunol. Methods, 226: 169-177.
CrossRefDirect Link - Pietta, P.G., 2000. Flavonoids as antioxidants. J. Nat. Prod., 63: 1035-1042.
CrossRefPubMedDirect Link - Mao, J.T., Q.Y. Lu, B. Xue, P. Neis and F.D. Zamora et al., 2019. A pilot study of a grape seed procyanidin extract for lung cancer chemoprevention. Cancer Prev. Res., 12: 557-566.
CrossRefPubMedDirect Link - Erlund, I., 2004. Review of the flavonoids quercetin, hesperetin and naringenin. Dietary sources, bioactivities, bioavailability and epidemiology. Nutr. Res., 24: 851-874.
CrossRefDirect Link - Peter, F. and D.H. Kim, 1990. In vitro inhibition of dihydropyridine oxidation and aflatoxin B1 activation in human liver microsomes by naringenin and other flavonoids. Carcinogenesis, 11: 2275-2279.
CrossRefDirect Link - Kanno, S.I., A. Tomizawa, T. Hiura, Y. Osanai and A. Shouji et al., 2005. Inhibitory effects of naringenin on tumor growth in human cancer cell lines and sarcoma S-180-implanted mice. Biol. Pharmaceut. Bull., 28: 527-530.
CrossRefDirect Link - Avelar, M.M. and C.M.C.P. Gouvea, 2012. Procyanidin B2 cytotoxicity to MCF-7 human breast adenocarcinoma cells. Indian J. Pharm. Sci., 74: 351-355.
CrossRefPubMedDirect Link - Chatelain, K., S. Phippen, J. McCabe, C.A. Teeters, S. O'Malley and K. Kingsley, 2011. Cranberry and grape seed extracts inhibit the proliferative phenotype of oral squamous cell carcinomas. Evidence-Based Complementary Altern. Med.
CrossRef - Tyagi, A., R. Agarwal and C. Agarwal, 2003. Grape seed extract inhibits EGF-induced and constitutively active mitogenic signaling but activates JNK in human prostate carcinoma DU145 cells: possible role in antiproliferation and apoptosis. Oncogene, 22: 1302-1316.
CrossRefDirect Link - Yamakoshi, J., M. Satio, S. Kataoka and M. Kikuchi, 2002. Safety evaluation of proanthocyanidin-rich extract from grape seeds. Food Chem. Toxicol., 40: 599-607.
CrossRefPubMedDirect Link - Lewandowska, U., K. Szewczyk, K. Owczarek, Z. Hrabec, A. Podsędek, D. Sosnowska and E. Hrabec, 2013. Procyanidins from evening primrose (Oenothera paradoxa) defatted seeds inhibit invasiveness of breast cancer cells and modulate the expression of selected genes involved in angiogenesis, metastasis and apoptosis. Nutr. Cancer, 65: 1219-1231.
CrossRefDirect Link