
Tanti Juwita
Department of Pharmacology and Clinical Pharmacy, Faculty of Pharmacy, Universitas Padjadjaran, Jl. Raya Bandung-Sumedang km 21 Jatinangor, West Java, Indonesia
Irma Melyani Puspitasari
Department of Pharmacology and Clinical Pharmacy, Faculty of Pharmacy, Universitas Padjadjaran, Jl. Raya Bandung-Sumedang km 21 Jatinangor, West Java, Indonesia
Jutti Levita
Department of Pharmacology and Clinical Pharmacy, Faculty of Pharmacy, Universitas Padjadjaran, Jl. Raya Bandung-Sumedang km 21 Jatinangor, West Java, Indonesia
LiveDNA: 62.1290
Pakistan Journal of Biological Sciences
Year: 2018 | Volume: 21 | Issue: 4 | Page No.: 151-165







ABSTRACT
In order to propose a prospective candidate for novel complementary phytopharmaceuticals, one of Zingiberaceae family plant, Etlingeraelatior or torch ginger, was being evaluated. The aim of this review was to provide a comprehensive literature research focused on the botanical aspects, nutritional quality, phytoconstituents and pharmacological activities of E. elatior. Researches on this particular plant were conducted in Malaysia (55.5%), Indonesia (33.3%), Thailand (8.3%) and Singapore (2.7%). This review article has revealed that the most prominent pharmacological activities were anti-microbial, anti-oxidant and anti-tumor activities in consistent with the dominated levels of flavonoids, terpenoids and phenols. However, extended and integrated research should be converged towards intensive investigations concerning to isolated phytoconstituents and its bioactivities, pharmacokinetics, bioavailability, molecular mechanism of its specific pharmacological activities, safety and efficacy studies for further development.
PDF Abstract XML References Citation
Copyright: © 2018. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Tanti Juwita, Irma Melyani Puspitasari and Jutti Levita, 2018. Torch Ginger (Etlingera elatior): A Review on its Botanical Aspects, Phytoconstituents and Pharmacological Activities. Pakistan Journal of Biological Sciences, 21: 151-165.
DOI: 10.3923/pjbs.2018.151.165
URL: https://scialert.net/abstract/?doi=pjbs.2018.151.165
DOI: 10.3923/pjbs.2018.151.165
URL: https://scialert.net/abstract/?doi=pjbs.2018.151.165
INTRODUCTION
The empirical use of medicinal herbs for hundreds of years indicates that Indonesia possesses an enormous opportunity to support the development of indigenous medication. One of the famous Indonesian original plant is Etlingera elatior (Jack) R. M. Smith (Zingiberaceae family).In Indonesia, the inflorescence bud is cultivated especially for culinary purpose to add flavor in traditional dishes and as a nutritional sources1. Interestingly, E. elatior is potentially used as a source of alternative medicine due to its afford ability, availability and compatibility as plant-derived drugs. This plant contains secondary metabolites such as phenols, flavonoids, glycosides, saponins, tannins, steroids, terpenoids, which revealed its bioactivities2,3. Therefore, E. elatior has been reported of its broad range of pharmacological activities including anti-hyperglycemic4, anti-hyperuricemic5, anti-inflammatory4, anti-microbials6, anti-oxidant7 and anti-tumor activity8. Moreover, for commercial used as natural ingredient in cosmetic formulation, inflorescence of torch ginger is also used for skin whitening9, anti-aging9 and lipstick preparation10.
This review article provides a comprehensive literature research focused on the botanical aspects, nutritional quality, phytoconstituents and pharmacological activities of E. elatior (Table 1).
Botanical aspects: The morphological characters of mature E. elatior were presented in Table 2.
The reproductive phase of this plant starts once the inflorescence shoot emerges from the rhizome. The major phases of the E. elatior developments are peduncle elongation, inflorescence occurrence, flowering and senescent stage. At the initial growth, peduncle elongation stage takes place 20 days to grow. The elongation of peduncle will slow down when the inflorescence bud is established during the blooming stage. After 30 days of shoots occurrence, the inflorescence bud starts to establish. E. elatior inflorescence consists of floral and involucral bracts. The floral bract of mature inflorescence at a full bloom may contain up to 20-25 layers, while the involucral bracts are only 3-4 layers. The seven development growth stages of inflorescence bracts are observed throughout the innermost layer (stage one to four) to the outermost layer (stage five to seven). The floral bracts are tender and smaller than involucral bracts. The purpose of involucral bracts is essentially for defense mechanism. The inflorescence proposes the flowering phase after 50 days of development. At the initial of the blooming phase, the outermost of involucral bract begins to disclose. By day 60, all of the involucral bracts will be completely disclosed and the bracts will be opened. During this phase, the first ring of the individual flowers will be opened gradually. From stage one to five, actual flowers were in the developing process. Through the 6th stage, the yellow margin of labellum appeared. Within stage seven, full blooming flowers arriving senesced stage and the labellum will be shut. The involucral bracts will dry slowly and the color become brown as the opening of true flowers continued17.
Phytoconstituents: E. elatior is widely used empirically as a traditional medicine because of its high level of secondary metabolites such as phenols, flavonoids, glycosides, saponins, tannins, steroids, terpenoids2,3. Total phenolic and flavonoids contents within E. elatior were found in high amounts in the leaves, flowers, stems and rhizome28. Flavonoids contained in E. elatior are quercetin, apigenin, kaempferol, luteolin, myricetin8. Kaempferol 3-glucuronide, quercetin 3-rhamnoside, quercetin 3-glucoside and quercetin 3-glucuronide were identified in the leaves37. Quercetin, kaempferol-3-O-glucoside and kaempferol were found in the flowers and stems16. The high content of kaempferol played a role as an anti-inflammatory agent and might help in decreasing risk of various types of brain disorders38. High level of anthocyanin was also found in this plant and had been shown to have cytotoxic activity against ovarian cancer cell lines31.However, anthocyanin is very susceptible to degradation due to increasing heating temperature39. Interestingly, anthocyanin in E. elatior extract had been proven to be more resistant to heating during food processing or pasteurization31.
Moreover, essential oils are found considerably rich in E. elatior, which had been identified in the leaves, stems, flowers and rhizome extracts. The essential oils were dominated by monoterpenes hydrocarbons whereas miscellaneous compounds were contained more in the flowers, oxygenated monoterpene derivates and oxygenated sesquiterpene derivatives were abundantly detected in the stems, while the sesquiterpene hydrocarbons were highly quantified in the leaves21. Based on the classification of essential oils, the most commonly compounds were β-pinene, caryophyllene, (E)-β-farnesene, 1,1-dodecanediol diacetate, (E) -5-dodecane, cyclododecane, 1-dodecanol, dodecanal, 17-pentatriacontene, dodecanoic acid, myrcene, α-humulene, camphene, β-pinene, Z-11-pentadecanol3,21,12,35.
Eight isolates of secondary metabolites derivatives of steroids, diarylheptanoids and labdane diterpenoids had been identified and isolated from ethyl acetate, n-hexane, chloroform rhizome extracts, which revealed potent cytotoxic activity18.
| Table 1: | Most important results of the main E. elatior articles |
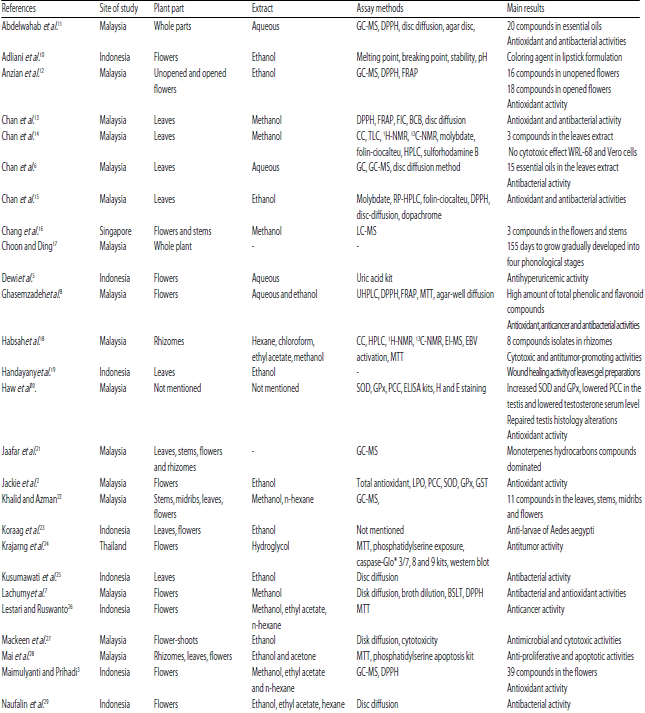 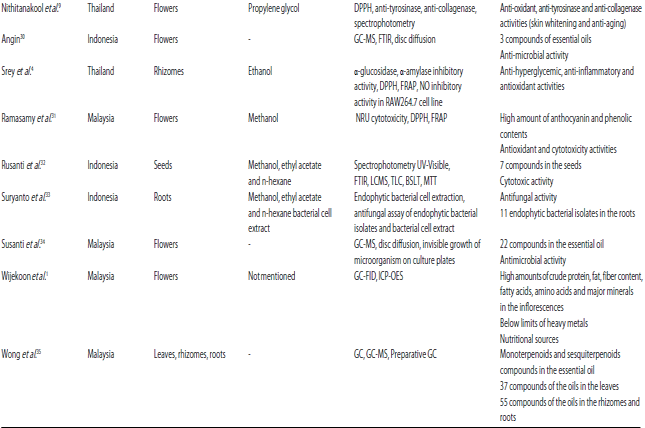 | |
*GC-MS: Gas chromatography mass spectrometry, DPPH: 2,2-diphenyl-1-picrylhydrazyl, FRAP: Ferric-reducing antioxidant power, FIC: Ferrous-ion chelating, BCB: $-carotene bleaching, CC: Column chromatography, TLC: Thin layer chromatography, H- and C-NMR: Nuclear magnetic resonance, HPLC: High-performance liquid chromatography, RP-HPLC: Reversed-phased HPLC, LC-MS: Liquid chromatography-mass spectrometry, MTT: 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazolium bromide, EI-MS: Electron ionization MS, EBV: Epstein-Barr virus, SOD: Superoxide dismutase, PCC: Protein carbonyl content, GPx: Glutathione peroxidase, GST: Glutathione S-transferase, H and E: Hematoxylin and Eosin, NO: Nitric oxide, BSLT: Brine shrimp lethality test, NRU: Neutral red uptakes, FTIR: Fourier-transform infrared spectroscopy, GC-FID: GC with flame ionization detector, inductively coupled plasma optical emission spectrophotometry (ICP-OES) | |
| Table 2: | Morphology and physiology of E. elatior36 |
 | |
Additionally, caffeoylquinic acids isomers, which are esters of caffeic and quinic acids were also isolated from E. elatior leaves due to the high amounts of chlorogenic acids (CGA) and caffeoylquinic acids (CQA)14. The phytoconstituents isolated from this plant are presented in Table 314.
Nutritional quality: In traditional dishes, E. elatior flower, inflorescence and fruit are widely used as a food ingredient because of the bright red shaped, distinctive taste and smell, which can increase the taste of the dishes. The consumption of E. elatior in foods is considered to have high nutritional value such as unsaturated fatty acids, proteins, amino acids and other mineral compounds as well as low contaminants of heavy metals. The composition of protein, fat and fiber in the dried flower were 12.6, 18.2 and 17%, respectively1. The high level of fiber content in this plant can potentially reduce blood cholesterol levels, hypertension, heart disease risk and constipation40. It was proven that the high content of saturated fatty acids were: Myristic acid>palmitic acid>stearic acid, whereas for the unsaturated were: Palmitoleic acid>linoleic acid>oleic acid1. Monounsaturated fatty acid and polyunsaturated acid contents were quantified up to 22.4 and 19.8%, respectively. In this basis, E. elatior could be used as an alternative source of fatty acids that is essential for various physiological processes and protection against various disease41.
Furthermore, the amino acid profiles of E. elatior inflorescence revealed the presence of non-essential amino acids higher than the essential amino acids, which supports the pharmacological activity to treat inflammation, as immune stimulating agents, anti-oxidant and anti-microbials agent42,43. The non-essential amino acids were dominated by glutamic acid and aspartic acid, while the essential amino acids were dominated by lysine, leucine, valine, threonine, isoleucine and phenylalanine. The inflorescence also contained high levels of major minerals like potassium, calcium, magnesium and phosphor, which are very fundamental for extracellular and intracellular roles and as building blocks of physical components in human body. Also interestingly, heavy metal contents are found at a lower concentration thus making it safer for daily diet consumption1.
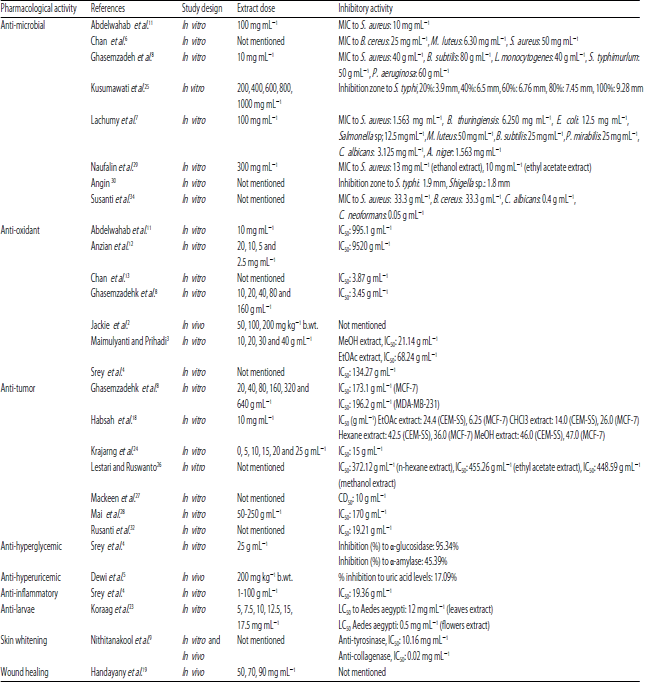
Pharmacological activities: Table 4 listed all of the pharmacological activities articles of E. elatior accompanied with the inhibitory activity, which are anti-microbial, anti-oxidant, anti-tumor, anti-hyperglycemic, anti-hyperuricemic, anti-inflammatory, anti-larvae, skin whitening and anti-aging and wound healing.
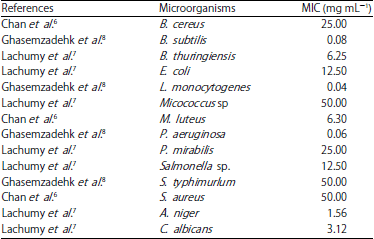
Anti-microbial activity: E. elatior has been reported to having significant capability as anti-bacterial agent in responding to the symptoms caused by pathogenic microbes. Leaves, rhizome and inflorescence extracts of this plant were found to have anti-bacterial and anti-fungal activity against various microbes (Table 5).
The highly potential anti-microbials activity possessed by E. elatior against microbial infections is due to its elevated contents of flavonoids, phenolic and terpenoids compounds. A study by Xu and Lee et al.44 showed inhibitory activity against MRSA by four flavonols such as datiscetin, kaempferol, quercetin, myricetin and two flavones such as flavone and luteolin44. Flavonoids mechanism in defeating microorganism pathogen is by targeting membrane cell wall due to its capability to composite with extracellular and soluble proteins45,46.
| Table 3: | Chemical compounds isolated from E. elatior |
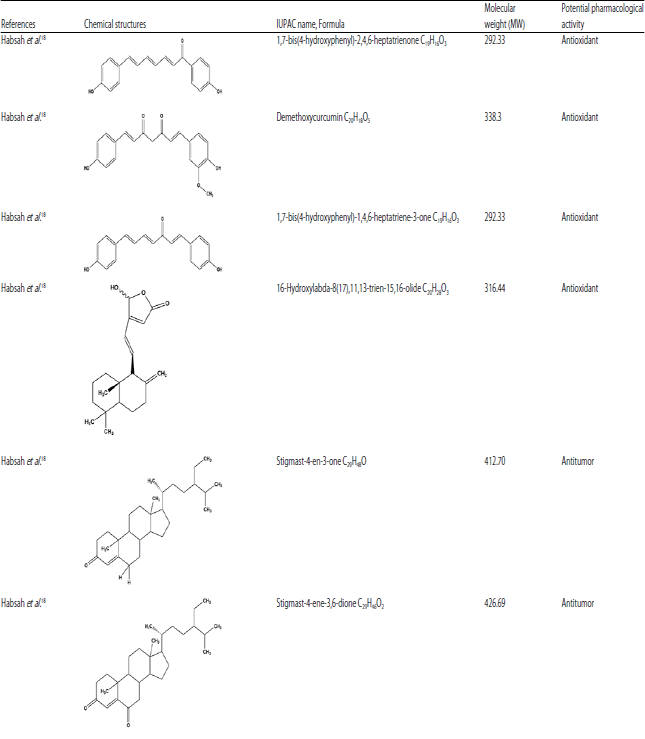 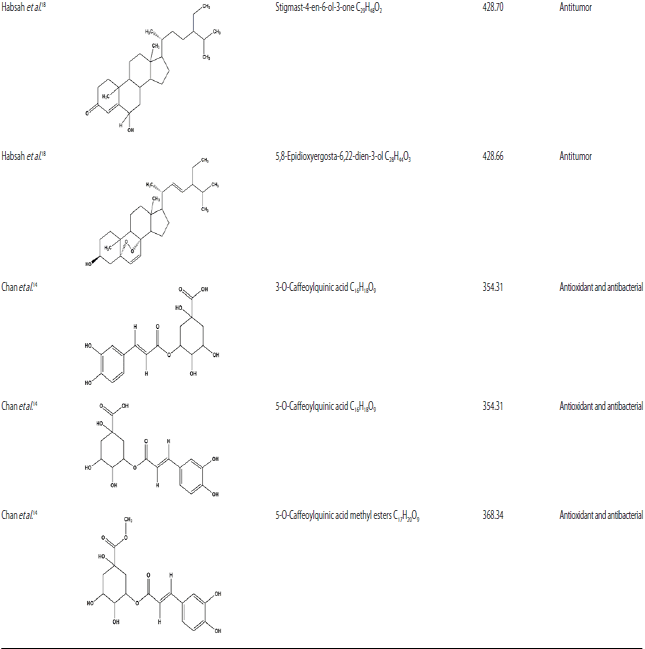 | |
| Table 4: | Pharmacological activity studies of E. elatior |
 | |
*S. aureus : Staphylococcus aureus , B. cereus: Bacillus cereus , M. luteus: Micrococcus luteus, B. subtilis: Bacillus subtilis, L. monocytogenes: Listeria monocytogenes , S. typhimurlum : Salmonella typhimurlum , P. aeruginosa : Pseudomonas aeruginosa , B. thuringiensis: Bacillus thuringiensis , E. coli: Escherichia coli , P. mirabilis : Proteus mirabilis , C. albicans: Candida albicans , A. niger : Aspergillus niger , C. neoformans: Candida neoformans, MeOH: Methanol, EtOAc: Ethyl acetate | |
| Table 5: | Antimicrobial activity (MIC) of E. elatior extract |
 | |
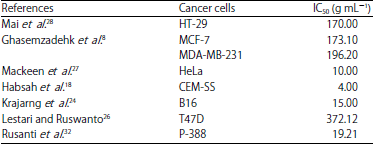
| Table 6: | Anti-tumor activity of E. elatior |
 | |
Furthermore, flavonoids with more hydroxyl group son the B-ring resulted greater destruction activity against microorganism membrane cell wall due to the more hydroxylation processes45,47.
Meanwhile, the effect of E. elatior as anti-fungal is predicted due to the high level of phenolic content especially by caffeic acid derivates which act on cell wall damage48, disrupting plasma membrane49, inhibition of isocitrate lyase enzyme activity50 and dimorphism inhibition49.
Caffeoylquinic acid isomers which are esters of caffeic and quinic acids, e.g., chlorogenic acid, are known to affect fungal cell membrane by forming pores and depriving potential electricity of the membrane and leading to the breakdown of membrane permeability barrier, ultimately causing cytoplasmic content leakage by releasing ions and other materials49. Chlorogenic acid derivatives will impair fungal cell wall by attacking1,3-β-D-glucan synthase, the essential enzyme in the forming of fungal cell wall48. Chlorogenic acid as antifungal can interrupt glyoxylate cycle with the isocitrate lyase enzyme inhibition, which in turn will disturb defense mechanism of fungal in nutrient-limited host niches50. Moreover, chlorogenic acid also shows an effect on fungal dimorphism by inhibiting and destructing the forming of hyphae or pseudohyphae49.
Endophytic microbes isolated from plants against various diseases have been proven in possessing pharmacological activities such as anti-cancer, anti-hyperglycemic, anti-oxidant, anti-inflammation, anti-microbials and anti-viral51-57. Moreover, endophytic bacteria isolated from E. elatior roots indicated anti-fungal activity against Culvularia sp., Fusarium oxysporum, Rigidoporusmicroporus, Rhizoctoniasolani and Saprolegnia sp. Eleven isolates of endophytic bacterial were obtained, in which seven of them are Gram-negative while the other four isolates were Gram-positive with diplococcus, streptobacillus and staphylococcus cell arrangements. The anti-fungal activity of endophytic bacterial isolated from E. elatior roots was predicted caused by alkaloid, saponin and terpene bioactive compounds33. Alkaloid works by disrupting (1,3)-β-glucan synthase, while saponin impairs to sterol in the cell membrane in increasing its permeability and protein denaturing of the cell membrane58. The mechanism of action of terpenes, i.e., monoterpenes and sesquiterpenes, as antifungal were reported by disrupting the membrane integrity and also destroying the fungal mitochondria59,60.
Anti-oxidant activity: E. elatior leaves, inflorescences and rhizomes exhibited diverse antioxidant activity level analyzed through ascorbic acid equivalent antioxidant capacity (AEAC) and gallic acid equivalent (GAE) in FRAP determination. Rhizomes were observed possessed the highest activity11. The scavenging effect against free radicals of E. elatior extract has often been revealed through the ability in reducing violet of DPPH to the yellow of DPPH-H and the ability to reduce ferric (Fe3+) tripyridyltriazine complex to ferrous (Fe2+), forming deep blue colour by FRAP assay determination12. The fresh whole plant extract of E. elatior proved potential antioxidant activity through DPPH assay11.The result of DPPH radical scavenging activity of methanol and ethanol extracts showed elevated percentage of DPPH by 76.26 and 63%, respectively7,8. Moreover, ethyl acetate fraction showed the lowest IC50 with 12.60 g mL–1 if compared to other solvents such as aqueous, ethanol, methanol, ethyl acetate, dichloromethane, butanol and hexane3,4,8.
Moreover, another study by Anzian et al.12 investigated the scavenging activity of three different drying methods (freeze-, oven- and sun-drying)of unopened E. elatior inflorescences by DPPH and FRAP assay and revealed that the freeze-drying method exerted the best free radical reduction with 89.24% of DPPH radical scavenging effect and 1973.01 mg/100 grams of the FRAP values and the IC50 obtained were 9.52 mg mL–1 12. The highest anti-oxidant activity of the freeze-drying method is probably due to the reduction of degradative enzyme activity, which leads to the absence of thermal degradation during the drying process. This particular method takes high extraction efficiency due to the forming of ice crystals within the plant matrix, leads to the rupture of cell structure and furthermore, causes cellular components leakage. High amount of solvent will enter the cells and ultimately give better extraction yield.61
Further study proved that E. elatior flower extract exhibited potent anti-oxidant activity by increasing the antioxidant enzyme levels such as SOD, GPx, GST and total anti-oxidants and also by lowering LPO and PCC proteins as biomarkers of protein oxidation due to oxidative stress2,20. It was supported by study investigations in lead acetate toxicity perturbation in inducing oxidative stress within rats serum levels2 and on a testicular functional damage and spermatogenesis capabilities of rats20.
The anti-oxidant activity of E. elatior revealed the potential effect of reducing free radicals in human physiology systems due to the high level of bioactive compounds such as phenolic and flavonoid8,11. The high amount of flavonoids content in E. elatior demonstrates various mechanisms in preventing injury caused by free radicals, which including direct scavenging activity against reactive oxygen (ROS), activation of anti-oxidant enzymes62, inhibition of oxidases63,64, metal chelating activity65, reduction of tocopheryl radicals63,66, increase in uric acid levels67, increase in anti-oxidant properties of low molecular antioxidants68, mitigation of oxidative stress caused by nitric oxide69. While, anti-oxidant mechanisms of phenolic compounds are based on the ability of donating hydrogen atom or electron to neutralize free radicals70, chelating metal ions70, acts as prooxidants71 and the ability of OH groups in phenolic to rehttps://docsdrive.com/images/ansinet/pjbs/2radical (ROO–)72.
Anti-tumor activity: Many studies have revealed the potent anti-tumor activity of E. elatior plant extracts against various cancer cells and have minimal effect against normal cells (Table 6). The antitumor activity of the aqueous extracts of E. elatior leaves showed potent activity against human breast cancer cell lines MCF-7 and MDA-MB-231 if compared to tamoxifen. Tamoxifen is highly toxic against normal cells, in which the concentration of tamoxifen inducing toxicity was more than 120.4 g mL–1, while the extract was not found to be toxic to normal cells8. Meanwhile, the combination of stigmasterol and -sitosterol isolated from E. elatior showed the highest potent anti-tumor activity with 85.1% of inhibition rate against Raji cell line18.
The acetone extract of E. elatior leaves indicated potent antiproliferative and apoptotic activity against colorectal carcinoma cells (HT-29) and did not interrupt any proliferation inhibition to the normal cells of Chinese Hamster Ovary (CHO) cells. Moreover, the acetone leaves extracts promoted the HT-29 cells death at the initial stage of apoptosis by PS apoptotic cells translocation from inner to the outer leaflet of cell membrane, while both caspase-3 activation and PS translocation took place at the later stage of apoptosis. The apoptosis activity against HT-29 cells was confirmed by ELISA assay with the extent of apoptosis and necrosis with 2.353-fold and 0.229-fold increases, respectively28.
A further study by Krajarng et al.24 proved the activity of E. elatior extract in inhibiting cancer cell progression on melanoma B16 cells through the caspase-independent pathway and the inhibition of ERK1/2, p38 and Akt signaling pathway, compared to Vero monkey cells24. The E. elatior extract has shown to induce early onset of apoptosis of cells through nuclear condensation, phosphatidylserin exposure and loss of mitochondrial membrane potential, which was acquired about 14% in the beginning of apoptosis stage at 4 and 8 h and the end stage of apoptosis increased by about 40% at 16 and 24 h, in which the apoptosis process appears when phosphatidylserine is transmitted from the inside to the outside of the cell membrane and will signal to the phagocytes to eat the cell. Based on the study, E. elatior extract did not enhance the expression of antiapoptosis and proapoptosis proteins such as of Bcl-2, Bax and caspase 3/7, 8 and 9 after extracts administration. However, Bim protein expression showed a significant improvement level, in which Bim is the proapoptotic protein subgroup of the third Bcl-2 family. To conclude, E. elatior extract of 15 μg mL–1 can only induce apoptosis through the expression of proapotosis protein of Bim and through the caspase-independent24. The mechanism of caspase-independent cell line mortality can be encouraged by factors liberated from mitochondria including endonuclease G (EndoG) and apoptosis-inducing factor (AIF), where either of these factors move to the cell nucleus and cause large-scale chromatin condensation and DNA fragmentation. The discharge of endoG from mitochondria is stimulated by BH3 proteins for instance tBid and Bim and inhibited by Bcl-273.
However, the effect of E. elatior extract in lowering the protein expression of extracellular signal-regulated kinase (ERK) and Akt has been reported. The expression of p38, ERK1/2, phosphorylated of p38 (P-p38) and phosphorylated of ERK1/2 (P-ERK1/2) decreased after the administration of E. elatior extract. Also, the extracts showed an inhibition of B16 melanoma cell through the suppression of Akt, which decreased total Akt activity and phosphorylated Akt (Ser473 and Thr308). ERK signaling can stimulate cell differentiation, proliferation and cell survival, in which c-Jun N-terminal kinase (JNK) and p38 are a stress-activated protein kinases24. The inhibition of ERK pathways in melanoma cells with BRAF inhibitors causes the cell cycle to stop and stimulate cell death by apoptosis74. The AKT pathway is a serine/threonine kinase protein that can stimulate cell growth, proliferation and survival. Phosphorylation Akt may inhibit the release of cytochrome c and apoptotic factors that would obstruct apoptosis and cancer cell development75.
The antiproliferative activity of E. elatior may be attributed to the high levels of flavonoids by the ortho-catechol moiety of a C2-C3 double bond and in ring B on flavones and flavanones. As to polymethoxylated-flavones, crucial contribution in providing high activity is given by the C3 hydroxyl and C8 methoxyl groups76. The flavonoids inhibition on tumor cells involved cell cycle arrest by interfering checking points regulation of both G1/S and G2/M and inhibiting all CDKs77-81 and also apoptosis promotion by inhibiting DNA topoisomerase I/II activity82-84, decreasing ROS85, regulating the expression of heat shock protein86, signaling pathways modulation87, down regulating of NF-kB, activating endonuclease and suppressing Mcl-1 protein73,86,88,89.
Anti-hyperglycemic activity: By inhibiting either α-glucosidase or α-amylase enzymes, it will further delay carbohydrate absorption and decrease sugar absorption after meals and eventually reduces glucose uptake and blood glucose89. E. elatior ethanol extract (25 μg mL–1) showed potent inhibition on α-glucosidase and α-amylase compared to acarbose inhibition4.
Anti-hyperuricemic activity: The anti-hyperuricemic activity of E. elatior flower extract in rats had been proven by Dewi et al.5 by determining uric acid level. The flower extract could suppress uric acid in blood serum up to 31.78%, while allopurinol was 45.65%. The potent anti-hyperuricemic activity was related to the high levels of polyphenols, flavonoids and saponin5. Flavonoids structure plays important role in inhibiting xanthine oxidase (XO) by controlling the overproduction of uric acid formation90. The double bond between C2 and C3 and also the hydroxyl groups at C5 and C7 were potentially exhibited inhibitory activity on XO64. The planar flavonols and flavones with a 7-hydroxyl group such as kaempferol, chrysin, quercetin, luteolin, isorhamnetin and myricetin had an elevated inhibition activity on XO by occupying the site of action competitively and by interacting with the enzyme either at a site or other than the active site90.
Anti-inflammatory activity: The study of anti-inflammatory activity of E. elatior plant has not been widely reported yet. The potent anti-inflammatory activity of E. elatior once reported by Srey et al.4 which conducted on the E. elatior rhizome extract inhibition to wards nitric oxide (NO) production in murine macrophage-like cell-line (RAW264.7). Rhizome extract at the concentration up to 100 g mL–1 could suppress the production of NO up to 91% with IC50 of 21 g mL–1 and compared to indomethacin with 85% of inhibition and 19 g mL–1 of the IC50. The anti-inflammatory activity of E. elatior plant was caused by the terpenoid aglycone4. For instance, several flavonoid and terpenoids groups have been reported to inhibit the expression of various enzymes related to the regulation of inflammation response such as inhibition of prostanoid biosynthesis (lipoxygenase, cyclooxygenase, inducible nitric oxide synthase), inhibition of histamine release, inhibition of phosphodiesterases, inhibition of protein kinases and transcription activation91. Water extract of red ginger could inhibit the rate of prostaglandin production in COX-1 and COX-292. Many phytoconstituents, e.g., flavonoids, diterpenoids, chalcones, interact with Arg120 and/or Tyr355 in COX-1 and with Tyr385 and/or Ser530 in COX-2, which are similar to the binding mode of NSAIDs. Nonetheless, based on the selectivity index calculation (cSI), only a few of the flavonoids and diterpenoids are categorized as selective COX-2 inhibitors93.
Toxicity study: E. elatior extract was found to be non-toxic based on the study of Lachumy et al.7 using brine shrimp lethality bioassay (LC50 = 2.52 mg mL–1) at 24 h. Thus, E. elatior extracts and the pharmaceutical properties contained can be categorized as biologically safe compounds7.
CONCLUSION AND FUTURE PERSPECTIVES
This review study on E. elatior, its phytoconstituents and pharmacological activities have revealed that the plant is potential to be utilized as phytopharmaceuticals. The most prominent pharmacological activities are anti-microbial, anti-oxidant and anti-tumor activities in consistent with the dominated levels of flavonoids and phenols. However, integrated research on the molecular mechanism of anti-inflammatory activity, particularly its inhibitory activity on NOS and modulation on NF-kappaB expression, as well as the level of flavonol synthase in the plant’s leaves, are being carried out.
REFERENCES
- Wijekoon, M.M.J.O., A.A. Karim and R. Bhat, 2011. Evaluation of nutritional quality of torch ginger (Etlingera elatior Jack.) inflorescence. Int. Food Res. J., 18: 1415-1420.
Direct Link - Jackie, T., N. Haleagrahara and S. Chakravarthi, 2011. Antioxidant effects of Etlingera elatior flower extract against lead acetate induced perturbations in free radical scavenging enzymes and lipid peroxidation in rats. BMC Res. Notes, Vol. 4.
CrossRefDirect Link - Maimulyanti, A. and A.R. Prihadi, 2015. Chemical composition, phytochemical and antioxidant activity from extract of Etlingera elatior flower from Indonesia. J. Pharmacogn. Phytochem., 3: 233-238.
Direct Link - Srey, C., C. Sontimuang, S. Thengyai, C. Ovatlarnporn and P. Puttarak, 2014. Anti α-glucosidase, anti α-amylase, anti-oxidation and anti-inflammation activities of Etlingera elatior rhizome. J. Chem. Pharm. Res., 6: 885-891.
Direct Link - Dewi, A.R., I. Nur’Aini, I.S. Bahri, H.N. Afifah, A. Fattah and W.A.S. Tunjung, 2016. Antihyperuricemic activity of ginger flower (Etlingera elatior Jack.) extract in beef broth-induced hyperuricemic rats (Rattus norvegicus). AIP Conf. Proc., Vol. 1755.
CrossRefDirect Link - Chan, E.W.C., Y.Y. Lim and M.A. Nor Azah, 2010. Composition and antibacterial activity of essential oils from leaves of Etlingera species (Zingiberaceae). Int. J. Adv. Sci. Arts, 1: 1-12.
Direct Link - Lachumy, S.J.T., S. Sasidharan, V. Sumathy and Z. Zuraini, 2010. Pharmacological activity, phytochemical analysis and toxicity of methanol extract of Etlingera elatior (torch ginger) flowers. Asian Pac. J. Trop. Med., 3: 769-774.
CrossRefDirect Link - Ghasemzadehk, A., H.Z.E. Jaafar, A. Rahmat and S. Ashkani, 2015. Secondary metabolites constituents and antioxidant, anticancer and antibacterial activities of Etlingera elatior (Jack) R.M.Sm grown in different locations of Malaysia. BMC Complementary Altern. Med., Vol. 15.
CrossRefDirect Link - Nithitanakool, S., V. Teeranachaideekul, L. Ponpanich, N. Nopporn, T. Junhunkit, P. Wanasawas and M. Chulasiri, 2014. In vitro and in vivo skin whitening and anti-aging potentials of hydroglycolic extract from inflorescence of Etlingera elatior. J. Asian Assoc. Sch. Pharm., 3: 314-325.
Direct Link - Adliani, N., Nazliniwaty and D. Purba, 2012. Lipstick formulation using natural dye from Etlingera elatior (Jack) R.M.Sm. extract. J. Pharm. Pharmacol., 1: 87-94.
CrossRefDirect Link - Abdelwahab, S.I., F.Q. Zaman, A.A. Mariod, M. Yaacob, A.H.A. Abdelmageed and S. Khamis, 2010. Chemical composition, antioxidant and antibacterial properties of the essential oils of Etlingera elatior and Cinnamomum pubescens Kochummen. J. Sci. Food Agric., 90: 2682-2688.
CrossRefPubMedDirect Link - Anzian, A.B., S. Rashidah, N. Saari, C.W. Sapawi and A.S.B.M. Hussin, 2017. Chemical composition and antioxidant activity of torch ginger (Etlingera elatior) flower extract. Food Applied Biosci. J., 5: 32-49.
Direct Link - Chan, E.W.C., Y.Y. Lim and M. Omar, 2007. Antioxidant and antibacterial activity of leaves of Etlingera species (Zingiberaceae) in peninsular Malaysia. Food Chem., 104: 1586-1593.
CrossRefDirect Link - Chan, E.W.C., Y.Y. Lim, S.K. Ling, S.P. Tan, K.K. Lim and M.G.H. Khoo, 2009. Caffeoylquinic acids from leaves of Etlingera species (Zingiberaceae). LWT-Food Sci. Technol., 42: 1026-1030.
CrossRefDirect Link - Chan, E.W.C., Y.Y. Lim and S.P. Tan, 2011. Standardised herbal extract of chlorogenic acid from leaves of Etlingera elatior (Zingiberaceae). Pharmacogn. Res., 3: 178-184.
CrossRefDirect Link - Chang, Y.Q., S.N. Tan, J.W.H. Yong and L. Ge, 2012. Determination of flavonoids in Costus speciosus and Etlingera elatior by liquid chromatography-mass spectrometry. Anal. Lett., 45: 345-355.
CrossRefDirect Link - Habsah, M., A.M. Ali, N.H. Lajis, M.A. Sukari, Y.H. Yap, H. Kikuzaki and N. Nakatani, 2005. Antitumour-promoting and cytotoxic constituents of Etlingera elatior. Malays. J. Med. Sci., 12: 6-12.
PubMedDirect Link - Handayany, G.N., Mukhriani and R.M. Halim, 2015. The wound healing effect of torch ginger ethanol extract (Etlingera elatior) in gel preparation to rabbits (Oryctolagus cuniculus). J. Farmasi FIK Univ. Islam Alauddin Makasar, 3: 54-58.
Direct Link - Haw, K.Y., S. Chakravarthi, N. Haleagrahara and M. Rao, 2012. Effects of Etlingera elatior extracts on lead acetate-induced testicular damage: A morphological and biochemical study. Exp. Ther. Med., 3: 99-104.
CrossRefDirect Link - Jaafar, F.M., C.P. Osman, N.H. Ismail and K. Awang, 2007. Analysis of essential oils of leaves, stems, flowers and rhizomes of Etlingera elatior (Jack) R.M. Smith. Malays. J. Anal. Sci., 11: 269-273.
Direct Link - Khalid, S.A. and A.N Azman, 2016. Identification of mosquito repellent in Etlingera elatior (Torch ginger). ARPN J. Eng. Applied Sci., 11: 6182-6185.
Direct Link - Krajarng, A., M. Chulasiri and R. Watanapokasin, 2017. Etlingera elatior extract promotes cell death in B16 melanoma cells via down-regulation of ERK and Akt signaling pathways. BMC Complementary Altern. Med., Vol. 17.
CrossRefDirect Link - Lestari, T. and Ruswanto, 2015. Anticancer potential of torch ginger flower extract with various of polarity to T47D cells. J. Kesehat Bakti Tunas Husada, 14: 8-11.
Direct Link - Mackeen, M.M., A.M. Ali, S.H. El-Sharkawy, M.Y. Salleh, N.H. Lajis and K. Kawazu, 1997. Antimicrobial and cytotoxic properties of some Malaysian traditional vegetables (Ulam). Int. J. Pharmacol., 35: 174-178.
CrossRefDirect Link - Naufalin, R., B.S.L. Jenie, F. Kusnandar, M. Sudarwamto and H. Rukmini, 2005. Antibacterial activity of kecombrang flower extract toward pathogenic and food spoilage bacteria. J. Teknoli Industri Pangan, 16: 119-125.
Direct Link - Susanti, D., N.A. Awang, H. Qaralleh, H.I.S. Mohamed and N. Attoumani, 2013. Antimicrobial activity and chemical composition of essential oil of Malaysian Etlingera elatior (Jack) RM smith flowers. J. Essent. Oil Bear. Plants, 16: 294-299.
CrossRefDirect Link - Wong, K.C., Y. Sivasothy, P.L. Boey, H. Osman and B. Sulaiman, 2010. Essential oils of Etlingera elatior (Jack) R.M. Smith and Etlingera littoralis (Koenig) Giseke. J. Essent. Oil Res., 22: 461-466.
CrossRefDirect Link - Williams, C.A. and J.B. Harborne, 1977. The leaf flavonoids of the Zingiberales. Biochem. Syst. Ecol., 5: 221-229.
CrossRefDirect Link - Jung, C.H., H.M. Seog, I.W. Choi and H.Y. Cho, 2005. Antioxidant activities of cultivated and wild Korean ginseng leaves. Food Chem., 92: 535-540.
CrossRefDirect Link - Boranbayeva, T., F. Karadeniz and E. Yilmaz, 2014. Effect of storage on anthocyanin degradation in black mulberry juice and concentrates. Food Bioprocess Technol., 7: 1894-1902.
CrossRefDirect Link - Ishida, H., H. Suzuno, N. Sugiyama, S. Innami, T. Tadokoro and A. Maekawa, 2000. Nutritive evaluation on chemical components of leaves, stalks and stems of sweet potatoes (Ipomoea batatas Poir). Food Chem., 68: 359-367.
CrossRefDirect Link - Benatti, P., G. Peluso, R. Nicolai and M. Calvani, 2004. Polyunsaturated fatty acids: Biochemical, nutritional and epigenetic properties. J. Am. Coll. Nutr., 23: 281-302.
CrossRefDirect Link - Bhat, R., K.R. Sridhar, C.C. Young, A.A. Bhagwath and S. Ganesh, 2008. Composition and functional properties of raw and electron beam‐irradiated Mucuna pruriens seeds. Int. J. Food Sci. Technol., 43: 1338-1351.
CrossRefDirect Link - Bernal, J., J.A. Mendiola, E. Ibanez and A. Cifuentes, 2011. Advanced analysis of nutraceuticals. J. Pharmaceut. Biomed. Anal., 55: 758-774.
CrossRefPubMedDirect Link - Xu, H.X. and S.F. Lee, 2001. Activity of plant flavonoids against antibiotic-resistant bacteria. Phytother. Res., 15: 39-43.
PubMedDirect Link - Cowan, M.M., 1999. Plant products as antimicrobial agents. Clin. Microbiol. Rev., 12: 564-582.
CrossRefPubMedDirect Link - Tsuchiya, H., M. Sato, T. Miyazaki, S. Fujiwara and S. Tanigaki et al., 1996. Comparative study on the antibacterial activity of phytochemical flavanones against methicillin resistant Staphylococcus aureus. J. Ethnopharmacol., 50: 27-34.
CrossRefDirect Link - Sato, M., S. Fujiwara, H. Tsuchiya, T. Fujii, M. Iinuma, H. Tosa and Y. Ohkawa, 1996. Flavones with antibacterial activity against cariogenic bacteria. J. Ethnopharmacol., 54: 171-176.
CrossRefDirect Link - Ma, C.M., T. Abe, T. Komiyama, W. Wang, M. Hattori and M. Daneshtalab, 2010. Synthesis, anti-fungal and 1,3-β-d-glucan synthase inhibitory activities of caffeic and quinic acid derivatives. Bioorg. Med. Chem., 18: 70009-70014.
CrossRefDirect Link - Sung, W.S. and D.G. Lee, 2010. Antifungal action of chlorogenic acid against pathogenic fungi, mediated by membrane disruption. Pure Applied Chem., 82: 219-226.
CrossRefDirect Link - Cheah, H.L., V. Lim and D. Sandai, 2014. Inhibitors of the glyoxylate cycle enzyme ICL1 in Candida albicans for potential use as antifungal agents. PloS One, Vol. 9.
CrossRef - Swarnalatha, Y. and B. Saha, 2016. Enhanced cytotoxic activity of endophytic bacterial extracts from Adhatoda beddomei leaves in A549 lung cancer cell lines. J. Cancer Res. Ther., 12: 1284-1290.
CrossRefDirect Link - Habbu, P.V., B.S. Patil, M.V. Salagare, S.D. Madagundi, R.V. Vanakudri, S.T. Shukla and V.H. Kulkarni, 2014. Antidiabetic potential of endophytic bacterial fraction of Murraya koenigii (L.) spreng. in rats. Spatula DD, 4: 139-150.
CrossRefDirect Link - Rahman, L., Z.K. Shinwari, I. Iqrar, L. Rahman and F. Tanveer, 2017. An assessment on the role of endophytic microbes in the therapeutic potential of Fagonia indica. Ann. Clin. Microbiol. Antimicrob., Vol. 16.
CrossRefDirect Link - Joe, M.M., S. Devaraj, A. Benson and T. Sa, 2016. Isolation of phosphate solubilizing endophytic bacteria from Phyllanthus amarus Schum & Thonn: Evaluation of plant growth promotion and antioxidant activity under salt stress. J. Applied Res. Med. Aromatic Plants, 3: 71-77.
CrossRefDirect Link - Padmanabhan, S.A., W.J. Wyson, S. Marimuthu, P. Saravanan and D. Anand, 2017. Isolation of endophytic bacteria, bioactive compounds and its antiviral activity against herpes simplex virus type-1. Int. J. Pharm. Sci. Drug. Res., 9: 113-117.
Direct Link - Pretsch, A., M. Nagl, K. Schwendinger, B. Kreiseder and M. Wiederstein et al., 2014. Antimicrobial and anti-inflammatory activities of endophytic fungi Talaromyces wortmannii extracts against acne-inducing bacteria. PloS One, Vol. 9.
CrossRef - Deba, F., T.D. Xuan, M. Yasuda and S. Tawatu, 2008. Chemical composition and antioxidant, antibacterial and antifungal activities of the essential oils from Bidens pilosa Linn. var. Radiata. Food Control, 19: 346-352.
CrossRefDirect Link - Tian, J., X. Ban, H. Zeng, J. He, Y. Chen and Y. Wang, 2012. The mechanism of antifungal action of essential oil from dill (Anethum graveolens L.) on Aspergillus flavus. PloS ONE, Vol. 7.
CrossRefDirect Link - Tian, J., B. Huang, X. Luo, H. Zeng, X. Ban, J. He and Y. Wang, 2012. The control of Aspergillus flavus with Cinnamomum jensenianum Hand.-Mazz essential oil and its potential use as a food preservative. Food Chem., 130: 520-527.
CrossRefDirect Link - Asami, D.K., Y.J. Hong, D.M. Barrett and A.E. Mitchell, 2003. Comparison of the total phenolic and ascorbic acid content of freeze-dried and air-dried marionberry, strawberry and corn grown using conventional, organic and sustainable agriculture practices. J. Agric. Food Chem., 51: 1237-1241.
CrossRefDirect Link - Nijveldt, R.J., E. van Nood, D.E.C. van Hoorn, P.G. Boelens, K. van Norren and P.A.M. van Leeuwen, 2001. Flavonoids: A review of probable mechanisms of action and potential applications. Am. J. Clin. Nutr., 74: 418-425.
CrossRefDirect Link - Heim, K.E., A.R. Tagliaferro and D.J. Bobilya, 2002. Flavonoid antioxidants: Chemistry, metabolism and structure-activity relationships. J. Nutr. Biochem., 13: 572-584.
CrossRefDirect Link - Cos, P., L. Ying, M. Calomme, J.P. Hu and K. Cimanga et al., 1998. Structure-activity relationship and classification of flavonoids as inhibitors of xanthine oxidase and superoxide scavengers. J. Nat. Prod., 61: 71-76.
CrossRefPubMedDirect Link - Ferrali, M., C. Signorini, B. Caciotti, L. Sugherini, L. Ciccoli, D. Giachetti and M. Comporti, 1997. Protection against oxidative damage of erythrocyte membrane by the flavonoid quercetin and its relation to iron chelating activity. FEBS Lett., 416: 123-129.
CrossRefPubMedDirect Link - Hirano, R., W. Sasamoto, A. Matsumoto, H. Itakura, O. Igarashi and K. Kondo, 2001. Antioxidant ability of various flavonoids against DPPH radicals and LDL oxidation. J. Nutr. Sci. Vitamonol., 47: 357-362.
PubMedDirect Link - Lotito, S.B. and B. Frei, 2006. Consumption of flavonoid-rich foods and increased plasma antioxidant capacity in humans: Cause, consequence, or epiphenomenon? Free Radic. Biol. Med., 41: 1727-1746.
CrossRefPubMedDirect Link - Yeh, S.L., W.Y. Wang, C.H. Huang and K.L. Hu, 2005. Pro-oxidative effect of β-carotene and the interaction with flavonoids on UVA-induced DNA strand breaks in mouse fibroblast C3H10T1/2 cells. J. Nutr. Biochem., 16: 729-735.
CrossRefDirect Link - Van Acker, S.A., M.N. Tromp, G.R. Haenen, W.J. van der Vijgh and A. Bast, 1995. Flavonoids as scavengers of nitric oxide radical. Biochem. Biophys. Res. Commun., 214: 755-759.
CrossRefPubMedDirect Link - Bravo, L., 1998. Polyphenols: Chemistry, dietary sources, metabolism and nutritional significance. Nutr. Rev., 56: 317-333.
CrossRefPubMedDirect Link - Lee, J., N. Koo and D.B. Min, 2004. Reactive oxygen species, aging and antioxidative nutraceuticals. Compreh. Rev. Food Sci., 3: 21-33.
CrossRefDirect Link - Jacobo-Velazquez, D.A. and L. Cisneros-Zevallos, 2009. Correlations of antioxidant activity against phenolic content revisited: A new approach in data analysis for food and medicinal plants. J. Food Sci., 74: R107-R113.
CrossRefDirect Link - Konig, A., G.K. Schwartz, R.M. Mohammad, A. Al-Katib and J.L. Gabrilove, 1997. The novel cyclin-dependent kinase inhibitor flavopiridol downregulates Bcl-2 and induces growth arrest and apoptosis in chronic B-cell leukemia lines. Blood, 90: 4307-4312.
Direct Link - Jiang, C.C., F. Lai, K.H. Tay, A. Croft and H. Rizos et al., 2010. Apoptosis of human melanoma cells induced by inhibition of B-RAFV600E involves preferential splicing of bimS. Cell Death Dis., Vol. 1, No. 9.
CrossRefDirect Link - Balmanno, K. and S.J. Cook, 2009. Tumour cell survival signalling by the ERK1/2 pathway. Cell Death Different., 16: 368-377.
CrossRefDirect Link - Kawaii, S., Y. Tomono, E. Katase, K. Ogawa and M. Yano, 1999. Antiproliferative activity of flavonoids on several cancer cell lines. Biosci. Biotechnol. Biochem., 63: 896-899.
CrossRefDirect Link - Senderowicz, A.M., 1999. Flavopiridol: The first cyclin-dependent kinase inhibitor in human clinical trials. Investig. New Drugs, 17: 313-320.
CrossRefDirect Link - Zi, X., D.K. Feyes and R. Agarwal, 1998. Anticarcinogenic effect of a flavonoid antioxidant, silymarin, in human breast cancer cells MDA-MB 468: Induction of G1 arrest through an increase in Cip1/p21 concomitant with a decrease in kinase activity of cyclin-dependent kinases and associated cyclins. Clin. Cancer Res., 4: 1055-1064.
Direct Link - Choi, J.A., J.Y. Kim, J.Y., Lee, C.M. Kang and H.J. Kwon et al., 2001. Induction of cell cycle arrest and apoptosis in human breast cancer cells by quercetin. Int. J. Oncol., 19: 837-844.
CrossRefDirect Link - Casagrande, F. and J.M. Darbon, 2001. Effects of structurally related flavonoids on cell cycle progression of human melanoma cells: Regulation of cyclin-dependent kinases CDK2 and CDK1. Biochem. Pharmacol., 61: 1205-1215.
CrossRefDirect Link - Wang, H.K., 2000. The therapeutic potential of flavonoids. Expert Opin. Invest. Drugs, 9: 2103-2119.
CrossRefDirect Link - Wang, I.K., S.Y. Lin-Shiau and J.K. Lin, 1999. Induction of apoptosis by apigenin and related flavonoids through cytochrome c release and activation of caspase-9 and caspase-3 in leukaemia HL-60 cells. Eur. J. Cancer, 35: 1517-1525.
CrossRefDirect Link - Bailly, C., 2000. Topoisomerase I poisons and suppressors as anticancer drugs. Curr. Med. Chem., 7: 39-58.
CrossRefDirect Link - Darwanto, A., M. Tanjung and M.O. Darmadi, 2000. Cytotoxic mechanism of flavonoid from Temu Kunci (Kaempferia pandurata) in cell culture of human mammary carcinoma. Clin. Hemorheol. Microcirc., 23: 185-190.
Direct Link - Lee, W.R., S.C. Shen, H.Y. Lin, W.C. Hou, L.L. Yang and Y.C. Chen, 2002. Wogonin and fisetin induce apoptosis in human promyeloleukemic cells, accompanied by a decrease of reactive oxygen species and activation of caspase 3 and Ca2+-dependent endonuclease. Biochem. Pharmacol., 63: 225-236.
CrossRefDirect Link - Rong, Y., E.B. Yang, K. Zhang and P. Mack, 2000. Quercetin-induced apoptosis in the monoblastoid cell line U937 in vitro and the regulation of heat shock proteins expression. Anticancer Res., 20: 4339-4345.
Direct Link - Yin, F., A.E. Giuliano and A.J. van Herle, 1999. Signal pathways involved in apigenin inhibition of growth and induction of apoptosis of human anaplastic thyroid cancer cells (ARO). Anticancer Res., 19: 4297-4303.
PubMedDirect Link - Iwashita, K., M. Kobori, K. Yamaki and T. Tsushida, 2000. Flavonoids inhibit cell growth and induce apoptosis in B16 melanoma 4A5 cells. Biosci. Biotechnol. Biochem., 64: 1813-1820.
CrossRefPubMedDirect Link - Tundis, R., M.R. Loizzo and F. Menichini, 2010. Natural products as α-amylase and α-glucosidase inhibitors and their hypoglycaemic potential in the treatment of diabetes: An update. Mini Rev. Med. Chem., 10: 315-331.
CrossRefDirect Link - Nagao, A., M. Seki and H. Kobayashi, 1999. Inhibition of xanthine oxidase by flavonoids. Biosci. Biotechnol. Biochem., 63: 1787-1790.
CrossRefPubMedDirect Link - Rathee, P., H. Chaudhary, S. Rathee, D. Rathee, V. Kumar and K. Kohli, 2009. Mechanism of action of flavonoids as anti-inflammatory agents: A review. Inflamm. Allergy - Drug Targets, 8: 229-235.
CrossRefDirect Link - Fikri, F., N.M. Saptarini, J. Levita, A. Mutalib and S. Ibrahim, 2016. The inhibitory activity on the rate of prostaglandin production by Zingiber officinale var. Rubrum. Pharmacol. Clin. Pharm. Res., 1: 33-41.
CrossRefDirect Link - Levita, J., M.R. Rositama, N. Alias, N. Khalida, N.M. Saptarini and S. Megantara, 2017. Discovering COX-2 inhibitors from flavonoids and diterpenoids. J. Applied Pharm. Sci., 7: 103-110.
CrossRefDirect Link