
Venkata Subba Reddy Gangireddygari
Department of Microbiology, Sri Krishnadevaraya University, 515 003 Anantapuramu, Andhra Pradesh, India
LiveDNA: 91.16213
DileepKumar Kanderi
Department of Microbiology, Sri Krishnadevaraya University, 515 003 Anantapuramu, Andhra Pradesh, India
Ramanjaneyulu Golla
Department of Microbiology, Sri Krishnadevaraya University, 515 003 Anantapuramu, Andhra Pradesh, India
Manjunatha Bangeppagari
Center for Biofluid and Biomimic Research,Pohang University of Science and Technology (POSTECH), 790-784 Pohang, South Korea
Vijay Anand Kumar Babu Gundi
Department of Biotechnology, Vikrama Simhapuri University, 524 001 Nellore, Andhra Pradesh, India
Khayalethu Ntushelo
Department of Agriculture and Animal Health, College of Agriculture and Environmental Sciences, Florida Science Campus, Corner Christiaan De Wet and Pioneer Avenue, Florida, University of South Africa, 1710 Johannesburg, Gauteng, South Africa
Rajasekhar Reddy Bontha
Department of Microbiology, Sri Krishnadevaraya University, 515 003 Anantapuramu, Andhra Pradesh, India
Pakistan Journal of Biological Sciences
Year: 2017 | Volume: 20 | Issue: 8 | Page No.: 410-422







ABSTRACT
Background and Objective: A widely used pesticide quinalphos (O, O-diethyl O-quinoxalin-2-yl phosphorothioate) may be an undesirable and persistent pollutant to non-target environments like rivers and other ecosystems. The objective of this study was to isolate a potential degradant bacterium of quinalphos from polluted soils and test its fitness under various culture conditions. Materials and Methods: A soil bacterium strain, capable of utilizing quinalphos as its sole source of carbon and energy was isolated from soil by enrichment method on a minimal salts medium (MSM). On the basis of morphological, biochemical and 16S rRNA gene sequence analysis the bacterium is a species of the genus Bacillus and it was closely related to Bacillus subtilis. Quinalphos degrading capabilities of this bacterium were assessed under different culture conditions. Quinalphos degradation data were analysed byusing a two-way ANOVA analysis with the Statistica v.10. Results: Bacillus subtilis grew on quinalphos with a generation time of 32.34 min or 0.54 h in the logarithmic phase. Maximum degradation of quinalphos was observed with an inoculum of 1.0 optical density, around pH-7.5 and at an optimum temperature of 35-37°C. Among the additional carbon and nitrogen sources, carbon source-glucose and nitrogen source-yeast extract marginally improved the rate of degradation of quinalphos. Gas chromatography-mass spectrometry (GC-MS) analysis of the culture of B. subtilis grown on quinalphos indicated the formation of one main metabolite-quinoxaline. Conclusion: The B. subtilis strain discovered in this study has a unique combination of abilities to degrade quinalphos and it is therefore suitable candidate bioremediator of quinalphos polluted environments.
PDF Abstract XML References Citation
Received: May 09, 2017;
Accepted: September 06, 2017;
Published: October 03, 2017
Copyright: © 2017. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Venkata Subba Reddy Gangireddygari, DileepKumar Kanderi, Ramanjaneyulu Golla, Manjunatha Bangeppagari, Vijay Anand Kumar Babu Gundi, Khayalethu Ntushelo and Rajasekhar Reddy Bontha, 2017. Biodegradation of Quinalphos by a Soil Bacterium-Bacillus subtilis. Pakistan Journal of Biological Sciences, 20: 410-422.
DOI: 10.3923/pjbs.2017.410.422
URL: https://scialert.net/abstract/?doi=pjbs.2017.410.422
DOI: 10.3923/pjbs.2017.410.422
URL: https://scialert.net/abstract/?doi=pjbs.2017.410.422
INTRODUCTION
Quinalphos (O,O-diethyl O-quinoxalin-2-yl phosphorothioate), is one of the widely used organophosphorus insecticides and acaricides. Being ranked moderately hazardous by the World Health Organization (WHO), quinalphos is either banned or restricted for usage in most nations. Quinalphos, which is classified as a yellow label (highly toxic) pesticide in India, is widely used on wheat, rice, groundnut, cotton, sugarcane, coffee and other ornamental crops. Andhra Pradesh tops the crop protection chemical business in India by contributing 20% of the total pesticides in this country. Among organophosphates, quinalphos is widely used in agriculture in Andhra Pradesh as reflected by the 5% of total sales of pesticides registered. However, only 1% of the pesticide, when applied, hits the target pest, while the remaining 99% of the pesticide drifts into the environment contaminating soil, water and biota1. In spite of the extensive usage of quinalphos in agriculture for pest control in crops, severe inhibition of acetylcholinesterase (AChE) in non-target organisms by quinalphos2-5 and adverse influence on blood and brain esterase activity in chickens6 and fertility efficiency in adult male rats7 have raised concerns about the lack of attention paid to various factors and in particular the biotic factors involved in the fate of quinalphos in natural resources8-12. A clear cut participation of factors, in particular biotic, in degradation of quinalphos in natural resources such as soil and water can only be demonstrated with the isolation of biotic agents with a degradation trait from natural resources. In this regard, the current study was designed to isolate bacterial species capable of degrading quinalphos in soil samples collected from agricultural fields and to further assess the extent of degradation by the biotic agent in liquid cultures under different conditions. This study aimed to advance the search for a candidate bioremediator of quinalphos polluted environments.
MATERIALS AND METHODS
Soils: Soil samples [organic matter-0.37(%), nitrogen-1.46(%) and pH-7.99] were collected from a vineyard in Akuthotapalli village, Anantapuramu district, Andhra Pradesh, India in 2010.
Chemicals: Technical-grade quinalphos purchased from Sigma-Aldrich (99.2% purity) was used as the sole source of carbon and energy in the bacterial growth culture. All the other chemicals and solvents used in the present study were of analytical reagent grade/high performance liquid chromatography (HPLC) grade and purchased from Sigma-Aldrich.
Culture medium and selective enrichment method: Mineral salts medium (MSM), formulated as follows (g L–1) 1.5 NH4NO3, 1.5 K2HPO4. 3H2O, 0.2 MgSO4.7H2O, 1.0 NaCl and 1 mL of trace elements stock solution which contained (g L–1):2.0 CaCl2.2H2O, 0.2 MnSO4.4H2O, 0.1 CuSO4.2H2O, 0.2 ZnSO4.H2O, 0.02 FeSO4.7H2O, 0.09 CoCl2.6H2O, 0.12 Na2MoO4.2H2O and 0.006 H3BO3 was used as the selective enrichment medium.
The selective enrichment involved incubation of 5 g samples of soil in MSM spiked with technical-grade quinalphos at 20 μg mL–1 of MSM in 250 mL Erlenmeyer flasks in an orbital shaker (Scigenics Biotech, Orbitek LE-IL Model, India). The temperature of incubation was 37°C and the rotation speed was 175 rpm. After 10 days of incubation, 5 mL of the culture was transferred to a fresh medium, amended by increasing the concentration of quinalphos up to 200 μg mL–1 of MSM in the Erlenmeyer flasks. The new cultures were incubated for further 10 days. After 5 more transfers, the culture was purified by the serial dilution transfer method and streak plated onto solidified MSM containing 20 μg mL–1 of quinalphos. Finally, a pure bacterial strain was obtained and designated as OP2.
Identification and characterization of bacterial isolate morphological, physiological and biochemical characterization: The isolated bacterial colony was observed under an optical compound microscope, its physiological and biochemical properties were determined following the procedures of Bergey’s Manual of Determinative Bacteriology13.
16S rRNA gene sequencing and phylogenetic tree analysis: Genomic DNA was extracted from the bacterial isolate (OP2) using the standard phenolic extraction procedure14 and amplification of the 16S rRNA gene was performed with the universal primers the 16 forward primer (sequence-5’-AGACTCAGGTTTGATCCTGG-3’) and 16 reverse primer(sequence-5’-ACGGCTACCTTGTTACGACTT-3’). Phylogenetic analysis of the 16S rRNA gene sequence was done according to the method used by Qin et al.15. The obtained 16S rRNA gene sequence was queried on the GenBank/EMBL database using BLAST16. The top BLAST hits were collected and aligned and a neighbor joining and a maximum-likelihood tree was constructed using the Robust Phylogenetic tree online tool17,18 to establish the phylogenetic relationship.
Measurement of bacterial growth kinetics on quinalphos: The bacterial isolate OP2 was grown overnight in 50 mL of MSM amended with 20 ppm of quinalphos per mL of MSM and yeast extract (0.1%) on an orbital shaker at 175 rpm at 37°C. Bacterial cells were harvested aseptically from the culture grown overnight (8000 rpm, 15 min, 4°C), thoroughly washed with MSM and suspended in sterile MSM to a get a suspension with the desired optical density (OD). For growth of the bacterial isolate on quinalphos, 50 mL of sterile MSM, spiked with quinalphos at a concentration of 20 μg mL–1, was dispensed into sterile 250 mL Erlenmeyer flasks. After inoculation with the bacterial culture to the final OD of 1.0 mL–1 of MSM, the flasks were incubated on an orbital shaker at 175 rpm at 37°C. Uninoculated flasks with fortified medium served as control. Five-milliliter aliquots of growing culture broth were withdrawn at 6 h intervals and the growth was monitored at wavelength 600 nm in a UV-visible spectrophotometer (Chemito-UV-2600, India). The count of viable bacterial colony-forming units was determined by serial dilution on nutrient agar. The specific growth rate of bacterial sp. OP2 was calculated in the logarithmic phase.
Biodegradation of quinalphos: The experiments on the biodegradation of quinalphos by the bacterial isolate were undertaken in 250 mL Erlenmeyer flasks in the same manner as is done for growth experiments (measurement of bacterial growth kinetics on quinalphos). Flasks containing quinalphos in MSM without inoculum served as controls. At regular intervals of 48 h, 10 mL of culture broth was aseptically withdrawn from flasks for growth measurements and residue analysis. Culture broth from the both uninoculated and inoculated flasks was processed for residue analysis and centrifuged at 8000 rpm for 15 min. The supernatants were recovered and an equal volume of dichloromethane was added, this was repeated 3 times with fresh lots of dichloromethane. The extracts were pooled together, dried over anhydrous sodium sulfate, filtered and allowed to dry at room temperature. The dried residue was dissolved in methanol for ultra-fast liquid chromatography (UFLC) [UFLC - LC 20 AD-Shimadzu, Japan] and gas-chromatography-mass spectrometry (GC-MS) [GC-MS-QP-5050 chromatograph (M/s. Shimadzu Instruments, Japan)] analysis.
Factors influencing the biodegradation of quinalphos: To assess the effect of various factors on the degradation of quinalphos by OP2, appropriate modifications were made to the supplementation of the nutrients in the MSM and to the growth conditions of the bacterial culture on quinalphos. For this purpose, the MSM was spiked with 20 mg L–1 of quinalphos and distributed in 250 mL flasks (100 mL flask). These new cultures were supplemented with an additional carbon source-(glucose or sodium acetate) or additional nitrogen sources - (NH4Cl, (NH4)2SO4, urea or yeast extract) to a final concentration of 0.01% (w/v). The flasks were inoculated with the bacterial suspension to an initial OD of 1.0 and flasks with no bacteria were the controls. The cultures were incubated at 37°C with a gentle rotation of 175 rpm and samples were drawn at 48 h intervals for residue analysis. The influence of the concentration of quinalphos on its degradation was assessed by growing the bacterial isolate on MSM containing different concentrations (20-200 ppm) of quinalphos. In another experiment, flasks containing MSM (pH-7.5) supplemented with 20 mg L–1 of quinalphos were inoculated with the bacterial cell suspension to an initial OD of 1.0 and incubated in a shaker at 175 rpm at different temperatures of (30-45°C) to study the influence of temperature on the degradation of quinalphos. In order to study the effect of pH on quinalphos degradation, the OP2 was cultured as described above and only the pH was varied from pH 5.5-8.5.
Analytical methods
Quinalphos residue analysis by ultra-fast liquid chromatography (UFLC): The residue of quinalphos extracted from the different experiments was dissolved in methanol and analyzed by UFLC equipped with a ternary gradient pump, programmable variable-wave length PDA detector, column oven, electric sample valve ODS-2 and C18, reverse phase column (4.6×250 mm×5 μm). The quinalphos residue analysis was conducted by using an isocratic mobile phase of methanol. The sample was injected at a volume of 20 μL, the mobile phase was programmed at a flow rate of 1 mL min–1 and quinalphos was detected at 254 nm wavelength under these operating conditions, with a retention time of 1.859 min.
Detection and identification of quinalphos metabolites by GC-MS analysis: MSM spiked with quinalphos at 20 μg mL–1 was distributed into 250 mL Erlenmeyer flasks at volumes of 50 mL flask in the same manner as mentioned earlier in the section measurement of bacterial growth kinetics on quinalphos. The flasks were divided into two main groups, one group receiving an inoculum of OP2 isolate and another devoid of inoculum (uninoculated/control). The two sets of flasks were incubated under optimal conditions at 37°C and 175 rpm for 10 days. At regular intervals, 10 mL aliquots of culture were withdrawn aseptically and the metabolites were extracted in the same manner as specified in the section biodegradation of quinalphos and analyzed in a GC-MS chromatograph with the column used as ZB-5 capillary column (25 m×0.32 mm). Toluene was used as an external standard for the quantification of the compounds.
Statistical analysis: All the parameters (carbon source, nitrogen source, size of inoculum, concentration of quinalphos, pH and temperature) were compared using a two-way ANOVA analysis. All were tested at the p<0.05 significance level and the Duncan multiple range test was used for separation between treatment means. Statistica v.10, StatSoft (USA) was used for all the statistical analyses19.
RESULTS
Identification and characterization of bacterial isolate: The bacterial strain (OP2) was isolated from a vineyard soil and cultured by a selective enrichment method and the procedures in the Bergey’s Manual of Determinative Bacteriology14 were followed to identify this strain. Microscopy revealed the bacterial strain is Gram-positive and rod-shaped, circular with a crenate margin. In addition, various biochemical tests were performed and recorded as follows: Indole test-negative, methyl red test-negative, VP test-negative, citrate utilization test-negative, glucose and lactose fermentation tests-negative, urease activity-negative, catalase activity-positive, nitrate-reductase activity-positive, starch hydrolysis-positive, casein hydrolysis-negative and gelatin liquefaction-positive. Based on these morphological and biochemical characteristics, the strain OP2 was considered to be homologous with Bacillus sp.
As the 16S rRNA gene sequence is a proven molecular and taxonomic tool used for the identification of bacteria isolated from the environment11,12,19-26, the same approach was adopted for the identification of bacterial isolate in the present study.
The 16S rRNA gene sequence was analyzed with the Robust phylogenetic tree online tool17,18 and a neighbor-joining dendrogram was constructed (Fig. 1).
Based on the dendrogram and the morphological and biochemical characteristics, OP2 showed 92% homology with Bacillus subtilis strain HPC20 and as a result was tentatively identified as Bacillus subtilis. The nucleotide sequence encoding the 16S rRNA of B. subtilis (1379 bases) was deposited in the GenBank database under the accession number KC540760.
Growth rate of Bacillus subtilis on quinalphos: The bacterial culture-B. subtilis was grown in MSM with a concentration of 20 μg mL–1 quinalphos. The culture was grown at 37°C with gentle shaking at 175 rpm. Viable cell counts of B. subtilis were done at regular intervals of 6 hours. The results of the viable cell counts are shown in Table 1.
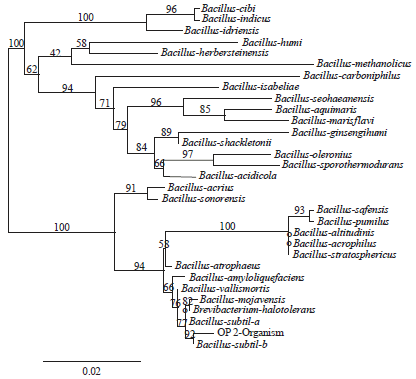 | |
| Fig. 1: | Phylogenetic tree based on the 16S rRNA gene sequences of strain OP2 |
Robust phylogenetic tree showing the phylogenetic relationship between strain OP2 and other related bacterial species based on the 16S rRNA gene sequences. Bootstrap values obtained with 1000 repetitions were indicated as percentages at all branches | |
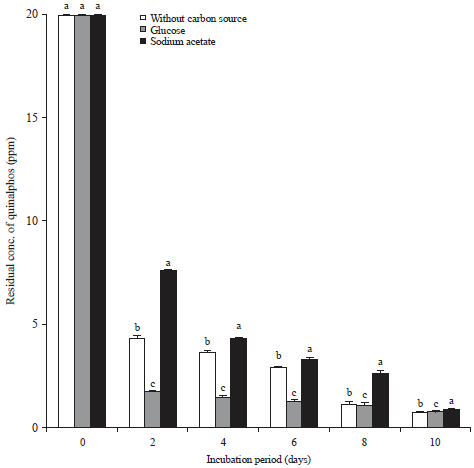 | |
| Fig. 2: | Influence of additional carbon source on biodegradation of quinalphos |
Values represented by bars with the same letter (a-c) are not statistically significant at p<0.05 and expressed in standard error mean (SEM) | |
| Table 1: | Growth of Bacillus subtilis on quinalphos |
 | |
The initial viable cell count with (20×108 CFU mL–1 of B. subtilis in the medium at the time of inoculation) rose to 130×108 CFU mL–1 within 6 h of incubation, then increased to 341×108 and 373×108 CFU mL–1 after 12 and 18 h of incubation respectively. During the log phase, the bacterial growth rate and generation time were calculated using the equation-K = log Nt-log No/log 2×t where Nt and No are the bacterial populations at times t and 0. The generation time of B. subtilis in the log phase was 32.34 min or 0.539 h.
Quinalphos was included in MSM as the sole source of carbon and energy for the cultivation of the bacterial culture in the present study. The proliferation of bacterial cells on quinalphos occurred in MSM up to 18 h, as reflected by the increase in the viable cell count (Table 1), which meant that quinalphos as the sole source of carbon was used by the bacteria.
Biodegradation of quinalphos by Bacillus subtilis : Variable factors such as additional carbon and nitrogen sources, the inoculum density, a medium pH, temperature and the concentration of quinalphos were examined to optimize the biodegradation of quinalphos by B. subtilis.
Influence of an additional carbon source on the biodegradation of quinalphos: In order to find out the influence an additional/supplementary carbon source on the biodegradation of quinalphos, the bacterial strain-B. subtilis was grown on quinalphos MSM with and without 0.01% glucose or sodium acetate. The concentrations of the quinalphos residues in the culture broth in the presence and absence of additional carbon were quantified and the results are presented in Fig. 2.
On the 2nd day of incubation, about 40% of the initially added quinalphos was recovered from the culture broth of B. subtilis with supplementation of sodium acetate, as opposed to the 8% recovered from the culture broth of B. subtilis grown in the presence of glucose. Twenty-three percent quinalphos was recovered from the culture broth of the same B. subtilis culture grown in the absence of additional carbon. Adding glucose to the culture medium marginally improved the degradation of quinalphos by B. subtilis.
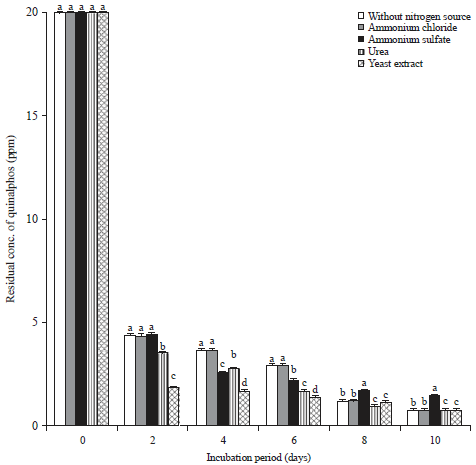 | |
| Fig. 3: | Influence of additional nitrogen source on biodegradation of quinalphos |
Values represented by bars with the same letter (a-d) are not statistically significant at p<0.05 and expressed in standard error mean (SEM) | |
Influence of an additional nitrogen source on the biodegradation of quinalphos: The effect of various nitrogen sources-(inorganic, in the form of ammonium chloride or ammonium sulfate and organic, in the form of urea and yeast extract) on the biodegradation of quinalphos by the bacterial isolate-B. subtilis was examined. The growth of B. subtilis on MSM without an additional nitrogen source (control) caused a 78.66% degradation of quinalphos within 2 days of incubation (Fig. 3). The addition of nitrogen sources-(ammonium chloride, ammonium sulfate, urea and yeast extract) to the MSM resulted in respective degradation of 78.37, 90.73, 82.44 and 92.8% of quinalphos by at the end of the 2 days incubation period. Analysis of these results in the present study therefore indicated that supplementation of the nitrogen with yeast extract marginally favored the quinalphos degradation by B. subtilis.
Influence of the inoculum density on the biodegradation of quinalphos: To assess the optimal inoculum density for the degradation of quinalphos, fortified MSM was tested with three different inoculum densities of the bacterial isolate, which is potentially B. subtilis. The cell suspension of the culture was diluted to different cell densities and added to MSM fortified with quinalphos to provide final bacterial cell densities of 0.5, 0.75 and 1.0 OD at A600 nm in the inoculated medium at 0 days. The culture was grown at 30°C in an orbital shaker and the residue of the parent compound (quinalphos) in the medium was quantified at 2 day intervals. The results of these tests are presented in Fig. 4. The corresponding figures of the quinalphos degradation in the culture of B. subtilis grown after inoculation at densities of 0.5, 0.75 and 1.0 OD at the end of 2 days intervals were 59.0, 68.75 and 78.6% respectively (Fig. 4). The highest cell density, used in the present study (i.e., 1.0 OD) caused the most degradation of quinalphos by B. subtilis. Quinalphos degradation occurred to a lesser extent at the lower inoculum densities of 0.5 and 0.75 OD of B. subtilis. It is therefore clear from the results of the present study that an inoculum density of 1.0 OD favored the degradation of quinalphos by B. subtilis.
Influence of the concentration of quinalphos on its biodegradation: To assess the degradation capacity of quinalphos by B. subtilis, MSM was fortified with quinalphos at different concentrations, which ranged from 20-200 mg L–1.
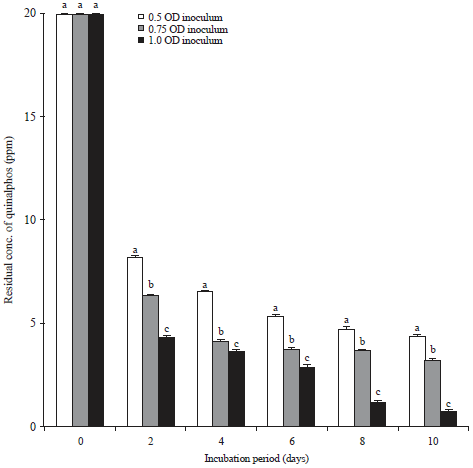 | |
| Fig. 4: | Influence of inoculum density on biodegradation of quinalphos |
Values represented by bars with the same letter (a-c) are not statistically significant at P < 0.05 and expressed in standard error mean (SEM) | |
| Table 2: | Influence of initial concentration of quinalphos degradation by Bacillus subtilis under sub-merged culture conditions |
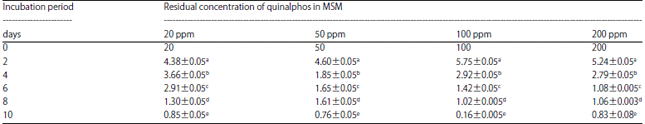 | |
Values labeled with the same letter (a-e) are not statistically significant at p<0.05. Range indicated in the table is standard error mean (SEM) | |
After inoculation, the culture was grown in MSM for 10 days with uninoculated controls. The quinalphos residue in the culture medium was quantified at 2 days intervals and the results, after subtraction of the control values, are presented in Table 2.
The concentration of quinalphos had no influence on the degradation of quinalphos by B. subtilis in the present study (Table 2). Degradation of quinalphos occurred to the extent of 75% in the culture of B. subtilis grown on MSM with the concentration of quinalphos at 20 mgL–1 and 98% in the same culture with a concentration of 200 mg L–1 of quinalphos. No appreciable degradation of quinalphos (1%) was observed in respect of the uninoculated controls in the corresponding period.
Influence of medium pH on the biodegradation of quinalphos: One important factor, which influences the degrading ability of microorganisms, is the pH. It needs to be assessed critically because it significantly determines the survival and activity of the microorganisms. The quinalphos degrading ability of B. subtilis was studied at different pH conditions, namely at a pH of 5.5, 6.5, 7.5 and 8.5 in MSM fortified with 20 mg L–1 quinalphos, along with a control (Fig. 5).
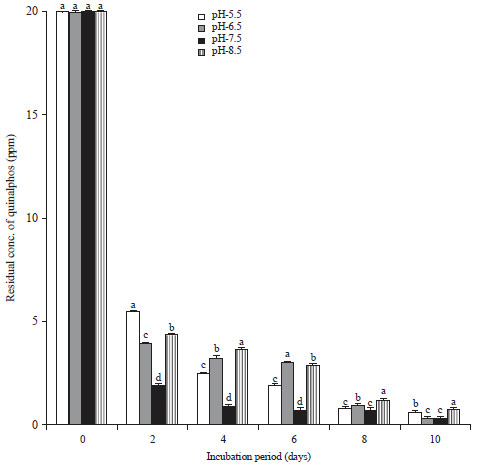 | |
| Fig. 5: | Influence of medium pH on biodegradation of quinalphos |
Values represented by bars with the same letter (a-d) are not statistically significant at p<0.05 and expressed in standard error mean (SEM) | |
The bacterial strain degraded quinalphos at all pH conditions, but to varying degrees. After 2 days of incubation, the quinalphos degradation had reached 73.1, 80.35, 90.31 and 78.6% in the culture of B. subtilis when the initial pH readings were 5.5, 6.5, 7.5 and 8.5, respectively. The pH 7.5 was evidently optimal for the maximum degradation of quinalphos by this bacterial strain.
Influence of temperature on the biodegradation of quinalphos: To assess the optimum temperature for the degradation of quinalphos, by the bacterial isolate-B. subtilis, the culture was cultivated in MSM fortified with 20 mg L–1 of quinalphos at different temperatures, i.e., 30, 35, 37, 40 and 45°C along with appropriate controls. Degradation occurred at all temperatures, but to varying degrees (Fig. 6): 78.6, 89.85, 91.08, 71.93 and 66.05% at the respective temperatures mentioned above, at the end of the 2 days interval. Based on these results, the temperature range of 35-37°C was considered optimal for the degradation of quinalphos by B. subtilis.
Identification of quinalphos metabolic intermediates through GC-MS analysis: In support of the data obtained from the growth of bacterial isolates and UFLC analysis, a further experiment was carried out to identify the metabolites, if any, formed during the degradation of quinalphos by the potential bacterial isolate B. subtilis. Cultivation of the bacterial isolate on MSM fortified with quinalphos was carried out under optimal conditions. Aliquots of the culture medium and uninoculated control were withdrawn at different time intervals and extracted with dichloromethane and the samples were analyzed through GC-MS as mentioned earlier in methodology section.
The GC-MS analysis indicated the presence of metabolites with a retention time of 23.057 min and different molecular ion [M+] distribution at m/z values of 298, 241, 157, 147, 130, 118 and 75 in a 48 h culture broth of B. subtilis grown on quinalphos (Fig. 7). One of the metabolites with an m/z value of 130 detected in the GC/MS analysis could be expected with the formation of quinoxaline during the degradation of quinalphos (Fig. 8). A tentative scheme of linkage to all the metabolites detected in the GC-MS analysis is shown in Fig. 8. Two metabolites 2-Hydroxyquinaxoline and diethyl phosphate in the degradation of quinalphos by Ochrobactrum sp. were identified by means of thin-layer chromatography, high-performance liquid chromatography (HPLC) and GC-MS11,25,26.
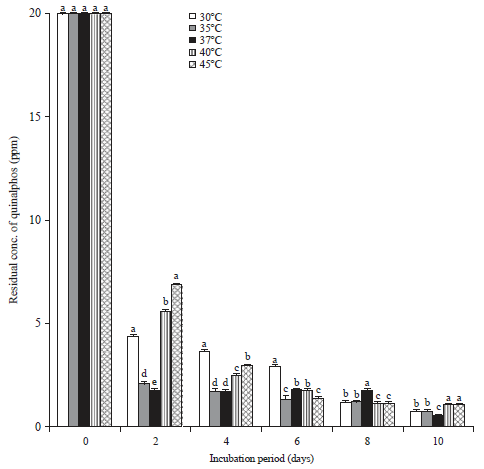 | |
| Fig. 6: | Influence of temperature on biodegradation of quinalphos |
Values represented by bars with the same letter (a-e) are not statistically significant at p<0.05 and expressed in standard error mean (SEM) | |
The formation of quinoxaline and other metabolites in the microbial metabolism of quinalphos needs to be further confirmed with authentic standards.
DISCUSSION
Bacillus subtilis degraded quinalphos to yield quinoxaline, the generation time was 32.34 min or 0.54 h in logarithmic phase and maximum degradation was observed with the inoculum of 1.0 optical density, around pH (7.5) and optimum temperature of 35-37°C. Among the additional carbon and nitrogen sources, carbon source-glucose and nitrogen source-yeast extract marginally improved the rate of degradation of quinalphos by B. subtilis. The attributes of B. subtilis to degrade quinalphos with this shorter generation time under these experimental conditions was probably due to its adaptations to the conditions of enrichment culture method. Similar studies conducted to investigate the degradation of various pesticides with Bacillus found results that were comparable to this study.
Similar to this study, which found that B. subtilis grew more rapidly with a shorter generation time, it was also found that B. thuringiensis cultured in a quinalphos-containing medium grew rapidly12.
Bacillus pumilus C2A1 degraded 86, 92 and 100% of chlorpyrifos within 5 days of incubation with the addition of yeast extract, nutrient broth and glucose, respectively to the culture medium27. In the presence of all three the nutrients, namely-yeast extract, nutrient broth and glucose, nearly 50% of the chlorpyrifos was degraded within 24 h of incubation. Glucose enhanced the degradation rate of B. pumilus C2A1, as it completely degraded the chlorpyrifos within 3 days of incubation. However, the results of the present study revealed that the supplementation of carbon had a different effect on the degradation of quinalphos by B. subtilis. The utilization of quinalphos by B. subtilis in the presence of glucose was only marginally improved. This might be due to the effect of the glucose on the growth of the bacterium.
The addition of yeast extract, nutrient broth and glucose to MSM enhanced the degradation of chlorpyrifos by Bacillus pumilus27 but degradation in the presence of yeast extract and the nutrient broth was less than in glucose. The recent report referred to earlier on, confirm that yeast extract as an, additional nitrogen source improve the degradation of quinalphos by Bacillus thuringiensis12. In the present study, supplementation of nitrogen did not improve the degradation of quinalphos, except when yeast extract was used.
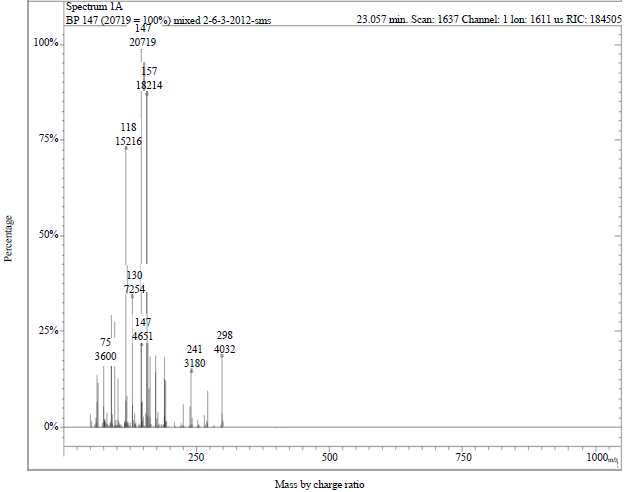 | |
| Fig. 7: | GC-MS chromatogram of B. subtilis culture broth after 48 h interval |
Bhadbhade28 reported that the monocrotophos degrading bacterial isolates–Arthrobacter atrocyaneus, Bacillus megaterium and Pseudomonas mendicona degraded monocrotophos within 48 h at an inoculum density of 108-109 cells mL–1 in a synthetic medium under aerated conditions. Three bacterial cultures, Brevundimonas sp., Bacillus sp. and Klebsiella oxytoca showed dimethoate degradation of up to 1000 mg L–1 in a culture medium with an inoculum of 107 cells mL–1. This was equivalent to 96, 94 and 71% of degradation within 12 days29. Anwar et al.27 assessed the influence of inoculum densities on the degradation of chlorpyrifos by the B. pumilus C2A1 strain. In cultures of B. pumilus C2A1 grown with lower inoculum densities, namely 105 and 107 CFU mL–1 there were longer lag phases, because the degradation of chlorpyrifos began after 1 and 2 days of incubation and resulted in 50 and 72% degradation of the pesticide respectively, after 5 days of incubation. In the same bacterial culture grown with 109 CFU mL–1 chlorpyrifos, degradation began rapidly, apparently without a lag phase and more than 80% degradation was achieved after 5 days of incubation. The lag phases were probably caused by the need for the microorganisms to adapt and multiply to reach the critical population size to secrete sufficient degradative enzymes so as to cause noticeable degradation. This may be the reason for the prolonged lag phase when the bacteria are grown in media with low densities of cells30. The recent report also found that the greater the size of the inoculum, the higher the degradation rate of quinalphos by Bacillus thuringiensis12.
Bhadbhade28 reported that three bacterial isolate cultures, Arthrobacter atrocyaneus, Bacillus megaterium and Pseudomonas mendicona-degraded a monocrotophos (MCP) concentration in the range of 100-500 mg L–1 in a synthetic medium under aerated conditions within 48 h. These three strains were able to tolerate MCP up to a concentration of 2500 mg L–1. This observation was in agreement with the results of the present study.
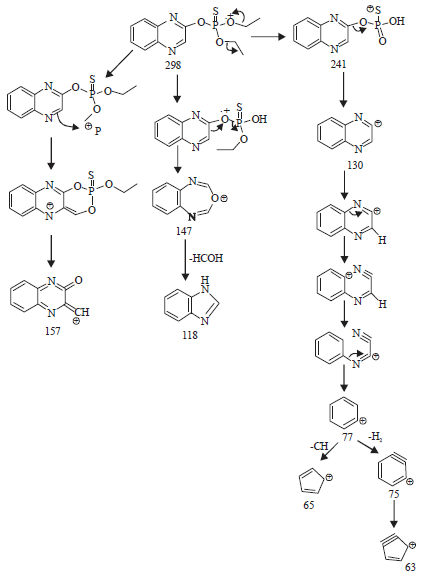 | |
| Fig. 8: | Tentative degradation pathway of quinalphos by Bacillus subtilis |
According to Deshpande29, three bacterial cultures-Brevundimonas sp., Bacillus sp. and Klebsiella oxytoca were able to degrade dimethoate in a culture medium at a concentration of upto 1000 mg L–1. These three strains respectively degraded 96, 94 and 71% of the dimethoate within 12 days. Anwar et al.27, found B. pumilus C2A1 to degrade chlorpyrifos at even higher concentrations. At 100, 200 and 300 mg L–1 concentrations, pesticide degradation started after 1, 2 and 3 days of incubation respectively and 73, 83 and 87% of the added chlorpyrifos was degraded within 10 days. At even higher concentrations of 500 and 1000 mg L–1, chlorpyrifos degradation began after 7 and 10 days of incubation and 88 and 89% of the pesticide was degraded after 2 weeks, in the respective instances. The recent report of Gangireddygari et al.12, demonstrated the degradation of quinalphos by Bacillus thuringiensis up to a concentration of 200 mg L–1. The prolonged lag phase in the degrading of chlorpyrifos at higher concentrations could be due to the time takes for the microorganism to produce sufficient degradative enzymes30.
Bhadbhade28 reported that the cultures of three bacterial isolates–Arthrobacter atrocyaneus, Bacillus megaterium and Pseudomonas mendicona degraded monocrotophos optimally at a pH of 7-8 in a synthetic medium under aerated conditions within 48 h. According to Deshpande29, three bacterial strains-Brevundimonas sp., Bacillus sp. and Klebsiella oxytoca degrade 96, 94 and 71% of dimethoate within 12 days, at the optimum pH of 7.
Anwar et al.27 reported that at an acidic pH B. pumilis C2A1 showed degradation of chlorpyrifos of nearly 50%, with a longer lag phase. At a relatively higher pH, no lag phase was observed and the degradation of chlorpyrifos was more efficient. Also, at as a neutral pH, more than 80% of the added pesticide was degraded. Talwar et al.11, reported that Ochrobactrum sp. strain HZM degraded quinalphos optimally at a pH of 7.0. Gangireddygari et al.12, most recently found that the optimum pH for the degradation of quinalphos by B. thuringiensis was 6.5-7.5. This observation is consistent with the result of the present study, namely that a pH of 7.5 is optimal for the degradation of this compound. In the present study, maximum degradation was achieved at a high pH. It is possible that some key enzyme(s) responsible for quinalphos degradation have their optimum enzymatic activity in high pH conditions.
Bhadbhade28 reported that cultures of three bacterial isolates, Arthrobacter atrocyaneus, Bacillus megaterium and Pseudomonas mendicona, degraded monocrotophos optimally at temperatures 30-35°C in a synthetic medium, with 77-88% of the degradation taking place under aerated conditions within 48 h. Cultures of Brevundimonas sp. and Klebsiella oxytoca showed maximum degradation of dimethoate at 30°C, whereas the Bacillus sp. displayed maximum degradation of dimethoate at 40°C29. Maximum quinalphos degradation by Bacillus thuringiensis occurred at temperatures of 30-45°C with the optimum temperatures of 30-37°C by Gangireddygari et al.12.
CONCLUSION
The bacterial isolate from soil samples of grapevine garden was identified as Bacillus subtilis through the selective culture enrichment method, morphological and biochemical analysis as well as sequence analysis of the 16Sr RNA gene. The optimum environmental conditions for growth and degradation of quinalphos were determined in shaking conditions and recorded as the inoculum density of (1.0.OD), pH (7.5), 35-37°C temperature and high concentration of quinalphos (200 ppm). Additional carbon and nitrogen sources, carbon source-glucose and nitrogen source-yeast extract marginally improved the rate of degradation of quinalphos. GC-MS analysis indicated the formation of quinoxaline in the pathway of quinalphos degradation by B. subtilis. From this research, this B. subtilis strain demonstrated superior qualities of quinalphos degradation. From these results, the authors could conclude that the Bacillus strain can be used to bioremediate quinalphos contaminated environments.
SIGNIFICANCE STATEMENT
This study presents the discovery of a Bacillus subtilis strain which is capable of degrading (utilize as a sole carbon source) the highly toxic pesticide active ingredient, quinalphos. The rate of degradation of quinalphos by B. subtilis was influenced by culture conditions e.g. temperature of incubation, pH, supplementation with additional carbon and nitrogen source, inoculum density and concentration of quinalphos. It was also discovered that 2-Hydroxyquinoxaline is a product of degradation of quinalphos by B. subtilis. With the findings of this study,this discovered B. subtilis can be recommended for depolluting the soil of quinalphos.
ACKNOWLEDGMENTS
This study was funded by University Grants Commission (UGC) (F.No. F. 33-205/2007 (SR) and Council of Scientific and Industrial Research-Senior Research Fellowship (CSIR-SRF) sanctioned (Lr. No. 09/383(0048)/2012-EMR-I), New Delhi.
REFERENCES
- Ramakrishnan, B., M. Megharaj, K. Venkateswarlu, R. Naidu and N. Sethunathan, 2010. The impacts of environmental pollutants on microalgae and cyanobacteria. Crit. Rev. Environ. Sci. Technol., 40: 699-821.
CrossRefDirect Link - Kopecka, J., A. Rybakowas, J. Barsiene and J. Pempkowiak, 2004. AChE levels in mussels and fish collected off Lithuania and Poland (Southern Baltic). Ocenologia, 46: 405-418.
Direct Link - Silman, I. and J.L. Sussman, 2005. Acetylcholinesterase: 'Classical' and 'non-classical' functions and pharmacology. Curr. Opin. Pharmacol., 5: 293-302.
CrossRefDirect Link - Vig, K., D.K. Singh and P.K. Sharma, 2006. Endosulfan and quinalphos residues and toxicity to soil microarthropods after repeated applications in a field investigation. J. Environ. Sci. Health Part B, 41: 681-692.
Direct Link - Chebbi, S.G. and M. David, 2009. Neurobehavioral responses of the freshwater teleost, Cyprinus carpio (Linnaeus.) under quinalphos intoxication. Biotechnol. Anim. Husbandry, 25: 241-249.
Direct Link - Sarkar, R., K.P. Mohanakumar and M. Chowdhury, 2000. Effects of an organophosphate pesticide, quinalphos, on the hypothalamo-pituitary-gonadal axis in adult male rats. J. Reprod. Fertil., 118: 29-38.
PubMedDirect Link - Goncalves, C., A. Dimou, V. Sakkas, M.F. Alpendurada and T.A. Albanis, 2006. Photolytic degradation of quinalphos in natural waters and on soil matrices under simulated solar irradiation. Chemosphere, 64: 1375-1382.
CrossRefDirect Link - Gupta, B., M. Rani, R. Kumar and P. Dureja, 2011. Decay profile and metabolic pathways of quinalphos in water, soil and plants. Chemosphere, 85: 710-716.
CrossRefDirect Link - Talwar, M.P., S.I. Mulla and H.Z. Ninnekar, 2014. Biodegradation of organophosphate pesticide quinalphos by Ochrobactrum sp. strain HZM. J. Applied Microbiol., 117: 1283-1292.
CrossRefDirect Link - Gangireddygari, V.S.R., P.K. Kalva, K. Ntushelo, M. Bangeppagari, A.D. Tchatchou and R.R. Bontha, 2017. Influence of environmental factors on biodegradation of quinalphos by Bacillus thuringiensis. Environ. Sci. Eur., Vol. 29.
CrossRefDirect Link - Qin, Q.L., D.L. Zhao, J. Wang, X.L. Chen and H.Y. Dang et al., 2007. Wangia profunda gen. nov., sp. nov., a novel marine bacterium of the family Flavobacteriaceae isolated from Southern Okinawa Trough deep-sea sediment. FEMS Microbiol. Lett., 271: 53-58.
CrossRefDirect Link - Altschul, S.F., W. Gish, W. Miller, E.W. Myers and D.J. Lipman, 1990. Basic local alignment search tool. J. Mol. Biol., 215: 403-410.
CrossRefPubMedDirect Link - Dereeper, A., S.Audic, J.M. Claverie and G. Blanc, 2010. BLAST-EXPLORER helps you building datasets for phylogenetic analysis. BMC Evol. Biol., Vol. 10.
CrossRefDirect Link - Dereeper, A., V. Guignon, G. Blanc, S. Audic and S. Buffet et al., 2008. Phylogeny.fr: Robust phylogenetic analysis for the non-specialist. Nucl. Acids Res., 36: W465-W469.
Direct Link - Nemutanzhela, M.V., D.M. Modise, K.J. Siyoko and S.A. Kanu, 2017. Assessment of growth, tuber elemental composition, stomatal conductance and chlorophyll content of two potato cultivars under irrigation with fly ash-treated acid mine drainage. Am. J. Potato Res., 94: 367-378.
CrossRefDirect Link - Yang, L., Y.H. Zhao, BX. Zhang, C.H. Yang and X. Zhang, 2005. Isolation and characterization of a chlorpyrifos and 3,5,6-Trichloro-2-pyridinol degrading bacterium. FEMS Microbiol. Lett., 251: 67-73.
CrossRefPubMedDirect Link - Pakala, S.B., G. Purushotham, B.P. Aleem, K.K. Ravi and B. Rajasekhar et al., 2007. Biodegradation of methyl parathion and p-nitrophenol: Evidence for the presence of a p-nitrophenol 2-hydroxylase in a Gram-negative Serratia sp. strain DS001. Applied Microbiol. Biotechnol., 73: 1452-1462.
CrossRefPubMedDirect Link - Yang, C., M. Dong, Y. Yuan, Y. Huang, X. Guo and C. Qiao, 2007. Reductive transformation of parathion and methyl parathion by Bacillus sp. Biotechnol. Lett., 29: 487-493.
CrossRefDirect Link - Caceres, T.P., M. Megharaj, S. Malik, M. Beer and R. Naidu, 2009. Hydrolysis of fenamiphos and its toxic oxidation products by Microbacterium sp. in pure culture and groundwater. Bioresour. Technol., 100: 2732-2736.
CrossRefDirect Link - Silambarasan, S. and J. Abraham, 2013. Kinetic studies on enhancement of degradation of chlorpyrifos and its hydrolyzing metabolite TCP by a newly isolated Alcaligenes sp. JAS1. J. Taiwan Inst. Chem. Eng., 4: 438-445.
CrossRefDirect Link - Reddy, G.V., B.R. Reddy and M.G. Tlou, 2014. Biodegradation of 2-Hydroxyquinoxaline (2-HQ) by Bacillus sp. J. Hazard. Mater., 278: 100-107.
CrossRefPubMedDirect Link - Reddy, G.S., M.M. Rafi, S.R. Kumar, N. Khayalethu and D.M. Rao et al., 2016. Optimization study of 2-Hydroxyquinoxaline (2-HQ) biodegradation by Ochrobactrum sp. HQ1. 3 Biotech, Vol. 6.
CrossRefDirect Link - Anwar, S., F. Liaquat, Q.M. Khan, Z.M. Khalid and S. Iqbal, 2009. Biodegradation of chlorpyrifos and its hydrolysis product 3,5,6-Trichloro-2-pyridinol by Bacillus pumilus strain C2A1. J. Hazard. Mater., 168: 400-405.
CrossRefDirect Link - Jilani, S. and M.A. Khan, 2004. Isolation, characterization and growth response of pesticides degrading bacteria. J. Biol. Sci., 4: 15-20.
CrossRefDirect Link