
Maryam Sohrabi
Department of Anatomical Sciences, Faculty of Medicine, Hamadan University of Medical Sciences, Shahid Fahmideh Street, 65178-3-8736 Hamadan, Iran
LiveDNA: 98.16590
Mahnaz Hosseini
Department of Anatomical Sciences, Faculty of Medicine, Hamadan University of Medical Sciences, Shahid Fahmideh Street, 65178-3-8736 Hamadan, Iran
Sevinc Inan
Department of Histology and Embryology, Faculty of Medicine, Celal Bayar University, Manisa, Turkey
Zohreh Alizadeh
Department of Anatomical Sciences, Faculty of Medicine, Hamadan University of Medical Sciences, Shahid Fahmideh Street, 65178-3-8736 Hamadan, Iran
LiveDNA: 98.14007
Mehrangiz Vahabian
Department of Anatomical Sciences, Faculty of Medicine, Hamadan University of Medical Sciences, Shahid Fahmideh Street, 65178-3-8736 Hamadan, Iran
LiveDNA: 98.13739
Ali Asghar Vahidinia
Department of Biochemistry and Nutrition, Faculty of Medicine, Hamadan University of Medical Sciences, Shahid Fahmideh Street, 65178-3-8736 Hamadan, Iran
Hosein Lahoutian
Department of Anatomical Sciences, Faculty of Medicine, Hamadan University of Medical Sciences, Shahid Fahmideh Street, 65178-3-8736 Hamadan, Iran
Pakistan Journal of Biological Sciences
Year: 2017 | Volume: 20 | Issue: 6 | Page No.: 289-297







ABSTRACT
Background and Objective: Spermatogenesis is a process by which germ cells produce spermatozoa and can be disturbed at every level. Nitric Oxide Synthases (NOS), implicate in interactions with Oxidative Stress (OS) which is one of the main factors in the etiology of male infertility. The High Fat Diet (HFD) is a major factor of obesity which in turn is important for enhancing OS. Antioxidants and garlic could attenuate or reverse effects of HFD. The aim of the study was to investigate the effects of dietary antioxidants and garlic on testicular inducible NOS (iNOS) and endothelial NOS (eNOS) in Wistar albino rats fed on HFD. Materials and Methods: Groups (each n = 8) were: SD (100% access to standard diet), F-HFD, (100% access to HFD) and R-HFD (70% access to HFD), F-HFD +antioxidants, F-HFD+garlic and R-HFD+antioxidants. The HFD consisted of a 60% fatty diet in 3 forms: Without antioxidants, with antioxidants and with garlic. The testicular iNOS and eNOS were studied by immunohistochemical (IHC) method. Also used ANOVA, repeated measures ANOVA, t-tests and Tukey’s test (where necessary) to analyze the data (p<0.05). Results: The iNOS increased in the F-HFD and R-HFD+antioxidants groups. The eNOS increased in R-HFD,F-HFD and F-HFD+garlic groups. The H-E evaluation in R-HFD group showed a decrease in spermatogenesis score count and seminiferous tubules diameters (μm) in comparison with the SD and F-HFD groups. R-HFD+antioxidants group had lower score than F-HFD+antioxidants and F-HFD+garlic groups. Conclusion: Restricted fat diet consumption causes increase in weight and impairs spermatogenesis. Results of this study reveal that adding the antioxidants can’t improve histological changes of testis. The iNOS expression in seminiferous tubules in restricted fat diet along with antioxidants, suggest a potential role of iNOS in spermatogenesis and male infertility.
PDF Abstract XML References Citation
Received: November 05, 2016;
Accepted: April 24, 2017;
Published: May 15, 2017
Copyright: © 2017. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Maryam Sohrabi, Mahnaz Hosseini, Sevinc Inan, Zohreh Alizadeh, Mehrangiz Vahabian, Ali Asghar Vahidinia and Hosein Lahoutian, 2017. Effect of Antioxidants on Testicular iNOS and eNOS after High-Fat Diet in Rat. Pakistan Journal of Biological Sciences, 20: 289-297.
DOI: 10.3923/pjbs.2017.289.297
URL: https://scialert.net/abstract/?doi=pjbs.2017.289.297
DOI: 10.3923/pjbs.2017.289.297
URL: https://scialert.net/abstract/?doi=pjbs.2017.289.297
INTRODUCTION
Spermatozoa are produced by germ cells in spermatogenesis, a multi-phase process capable of being disrupted at any phase1. Magnusdottir et al.2, also showed the relationship between obesity and reduction of sperm count. Obesity can have genetic roots but based on some evidence it can also be attributed to environmental factors such as increase in the amount of fat in the diet of the world’s population in the recent decades3,4. Additionally, research has shown that High Fat Diet (HFD) is the main cause of obesity4.
Human body is normally exposed to endogenous and exogenous free radicals. However exces sive production of free radicals and inability of the body to neutralize them as well as underproduction of antioxidant lead to Oxidative Stress (OS)5. Reactive Oxygen Species (ROS) including free radicals damage cell’s DNA in the process of oxidation and in its turn causes many disease5. Up to now, some clinical studies have been carried out on the correlation between OS and male infertility but few done on the destructive effect of HFD on spermatogenesis6,7. On the contrary, the impact of HFD on female fertility has been already researched8. Previous studies showed the obesity increase OS in men9 and male infertility is an adverse consequence of increasing OS7. The OS lead to the decline of leydig cells steroidogenic capacity and loss of germinal epithelium capacity to differentiate normal spermatozoa. Chronic hypoxia and normal oxygen supply had opposing effect on physiological mechanisms of spermatogenesis and male fertility. Chronic hypoxia induced a state of oligospermia and the normal oxygen supply resulted in the normalization of spermatogenesis. Ischemic reperfusion damage to the testis was observed due to the generation of ROS under the condition of OS which is considered the main cause of male infertility. Damage to the antioxidant mechanisms increase in obese patients and animal models are clinically documented10.
Supplying subjects with antioxidant prevents many HFD-related disease which increase OS in different tissues11. Antioxidant along with the main enzymes processing ROS, protect testes against oxidative damage. The antioxidants and a large number of free radical scavengers challenge ions. Spermatogenesis is reactivated in subjects taking dietary vitamins and semen quality is promoted in those receiving fat-and water soluble vitamins12,13. According to a study, OS resulting from lack of vitamin E and C disturbs spermatogenesis while a diet containing them reactivate the process14. Another study showed that vitamin E and C supplementation decreased testicular toxicity although it did not completely reverse the damage15. Garlic and many garlic preparations also have antioxidant properties. Garlic has been proved to be efficient in aiding endogenous antioxidant defense in different animals16. Previous study has reported contrary effects of garlic on spermatogenesis17. A report showed positive effect of garlic on the process while the other claimed that different doses of crude garlic in a month could reduce spermatogenesis18.
Nitric oxide (NO), a ubiquitous free radical and freely diffusible water and lipid-soluble gaseous molecule with a short half-life, is synthesized in all mammalian cells by nitric oxide synthases (NOS). With regard to the effect of both deficiency and excess of NO production in the dysfunction of cells and tissues, studying the role of diet in the regulation of NO synthesis in humans and animals to keep them healthy and protected against diseases is critical19. There are three isoforms of the NOS: nNOS (originally in neuronal tissue), iNOS (originally as being inducible by cytokines) and eNOS (originally in endothelial cells). In the testis, eNOS is expressed in leydig cells, sertoli cells, spermatocytes and spermatids while various cytokines inducing transcription of iNOS is required for its expression19. In the recent decades, previous study has shown the direct relationship of dietary factors such as cholesterol, fatty acids and vitamins with constitutive and inducible NO production in mammalian cells20.
Many assisted reproduction techniques have been recently devised to study spermatogenesis21. Based on a study, the role of obesity in infertility of men was 3 times more than idiopathic and female factors in the couples22. The aim of the present study was to measure immunohistochemical expression of testicular iNOS and eNOS after treatment with different types of diets: standard diet, high-fat diet, high-fat diet with crude garlic, high-fat diet with antioxidants, for a 12 weeks period. The study was designed to test the hypothesis that antioxidant supplementation could attenuate or partially reverse the effects of a high-fat diet on expression of iNOS and eNOS in testis.
MATERIALS AND METHODS
Experimental animals: Forty eight 2 months old Wistar Albino rats were utilized in this study. They were divided into five mice in each cage under standard conditions, i.e. 12 h light, 12 h dark, 60±5% humidity, 30±2°C temperature and providing with food and water for 1 week. Research ethics approval from the Ethics Committee of the School of Medicine, Hamadan University (Hamadan, Iran) was obtained. Then rats were divided into six (each n = 8) groups: 3 controls: SD (100% access to standard diet), F-HFD (100% access to HFD) and R-HFD (70% access to HFD) and 3 treatments:
| Table 1: | Holstein’s scores for spermatogenesis classification |
 | |
| Table 2: | Holstein’s normal spermatogenesis characteristics |
 | |
F-HFD+antioxidants, F-HFD+garlic and R-HFD+ antioxidants for 12 weeks. The received energy was measured weighing of all groups every day during the study.
High fat diet and antioxidants composition: The ingredients for every 100 g of HFD were milk fat (31.65 g), soya oil (3.22 g), casein (25.84 g), maltodextrin (16.14 g), sucrose (8.9 g), alpha (α)-cellulose (6.5 g), potassium citrate (2.1 g), dicalcium phosphate (1.7 g), L-cysteine (0.37 g), calcium carbonate (0.7 g), choline chloride (0.24 g), mixed vitamins (1.27 g) and mixed minerals (1.27 g), mixed minerals (1.25 g), mixed vitamins (1.29 g). Mixed vitamins composition was: 2 g kg–1 vitamin E, 2 g kg–1 vitamin C and 0.6 g kg–1 astaxanthin)23,24.
Histochemical evaluation: The procedure was ended 12 weeks after feeding the groups with the diets. Under sterile conditions, the animals were anesthetized by general method of intramuscular injection of ketamine and xylazine hydrochloride. All left testes were dissected and cranial halves were put in buffered formalin 10% fixative solution 72 h at +4°C for histological and immunohistological evaluation. Next, specimens were washed in 70% ethanol and immersed in a graded series of ethanol and cleared in xylene and then were embedded in paraffin. Approximately 5 μm thick sections were cut. Slides stained with Hematoxylin-eosin (HE) were used for morphological diagnosis and also the comparison of seminiferous tubule widths and scoring of spermatogenesis. Using an ocular micrometer, the mean diameter of seminiferous tubules was estimated in 10 randomly selected fields by measuring three diagonal diameters of 10 tubules (60 tubules in total) based on Holstein’s score Table 122.
To assess the stages of spermatogenesis quantitatively, abnormalities were evaluated for each testis using light microscopy and the spermatogonium, sertoli and interstitial cells have been considered. Afterwards, the characteristics of normal spermatogenesis were assayed adopting Holstein’s classification (Table 2)22. Diameter of the seminiferous tubule was 65 μ in the control group of the present study for rat (180 μ for human according to Holstein’s study)22 at the minimum.
Immunohistochemical evaluation: For immunohistochemical staining, sections (3 μm thick) were cut and then deparaffinization at 60°C during the night, immersion in PBS and treatment with 0.3% hydrogen peroxide in PBS for 10 min to block endogenous peroxidase activity was done successively. The sections were washed in PBS and exposed in 10% normal horse serum to suppress non-specific antigen. A mouse monoclonal iNOS antibody (Cat. 905-385 Neomarkers Inc. Fremont, CA, USA) and a polyclonal eNOS antibody (Cat. RB-1711-P, Neomarkers, Inc. Fremont, CA, USA) were used for immunohistochemistry. The incubation of the sections with eNOS (1:100) or iNOS (1:100) antibodies were done for the whole night at +4°C (To show specificity of the antibodies, normal IgG have been used instead of the primary antibodies for negative control sections). The sections were washed in PBS, then, the addition and incubation of appropriate biotinylated secondary antibodies (Cat.85-9043 Secondary kit Invitrogen Histostain plus kit, Broad spectrum, CA, USA) were performed for 30 min at room temperature and after that the avidin-biotin-peroxidase complex was added. The reagents of DAB kit (Spring Bioscience Inc. CA, USA) developed immunostaining. The sections were dehydrated and mounted (Surgipath, USA) after counter-staining with Hematoxylin Mayer (JJ Baker, Holland). The specimens were analyzed by B× 40 light microscope (Olympus, Tokyo). The staining scores evaluation were blindly performed by 2 observers separately. The iNOS and eNOS positive IHC stained cells in sections turn brown and their intensity was considered as negative (-), mild (+), moderate (++) and intense (+++).
Estimation of average expression of iNOS and eNOS (%) was carried out by counting positive cells (at least 500 cells in each testis) in 5 random medium-power fields (400×) including 100 cells and finally the total was divided by five.
Statistical analysis: Data was analyzed using Statistical Package for the Social Sciences (SPSS, Inc., Chicago, IL, USA) program, version 16.0. One-way analysis of variance (ANOVA), repeated measures ANOVA, t-tests and Tukey’s test were used to analyze the data. A p-value<0.05 indicated statistical significance25.
RESULTS
In this study animals were fed with one of the four types of HFDs including: Standard diet, high-fat diet with or without antioxidants and high-fat diet with crude garlic for 12 weeks.
Animal weight determinations are shown in Table 3. All rats gained weight significantly throughout the experimental period (p<0.001). However, R-HFD+antioxidants rats achieved a final lower body weight as compared with the other groups (Table 3).
The daily weight gain by the treated rats with highest value from F-HFD (1.81±0.54 g) followed by F-HFD+antioxidant in relation to the R-HFD and R-HFD+antioxidants groups, respectively. The feed efficiency ratio of rats fed with R-HFD increased significantly (p<0.05) compared to those with other groups. Interestingly, R-HFD+antioxidants have shown the lowest feed efficiency ratio among all groups (Table 4).
In the SD group, normal spermatogenesis was observed and the mean Holstein’s score was 10. In F-HFD and R-HFD groups, some sections of testes contained a few tubules with poor spermatogenesis and the score (9 and 8, respectively) were slightly lower than in the SD group. There were no differences between the scores in F-HFD+antioxidants and F-HFD+garlic groups (7 for both). The R-HFD+antioxidants group showed a more decrease in the mean Holstein’s score (6). The results of Holstein’s scoring are depicted in Table 1.
The results showed that in F-HFD group, the average diameter of seminiferous tubules was significantly lower than the SD and R-HFD groups (p<0.05). There was also significant reduction in the average diameter in R-HFD+antioxidants compared to F-HFD+antioxidants and F-HFD+garlic groups (p<0.05) (Table 5).
The presence of a continuous Basal Lamina (BM), spermatogonia in the basal compartment of seminiferous epithelium and spermatocytes I, spermatocytes II, spermatids and spermatozoa in the adluminal compartment in testes is shown in Fig. 1. The results from the immunohistochemical staining (Fig. 1) generally detected the eNOS and the iNOS in the cytoplasm of spermatogonia, spermatocyte I, spermatocyte II, spermatid and leydig cells in all experimental groups. There was, however, a considerable variability in the intensity of staining between testicular cells of them: (1) Intense immunoreactivity (+++) was shown for eNOS spermatocytes II and spermatids of F-HFD+garlic group and (2) iNOS in these cells were intense in F-HFD group.
| Table 3: | Body weight of rats after 12 weeks on the experimental diets |
 | |
1Delta body weight gain was calculated by subtracting the body weight of the rat the day of the final day from that of the first day of the experiment. The values are shown as Mean±SE. (*Paired t-test and **student t-test) (p<0.05), NS: Non-significant | |
| Table 4: | Mean weight gain, food consumption, daily received energy and feed efficiency ratio of rats after 12 weeks on the experimental diets |
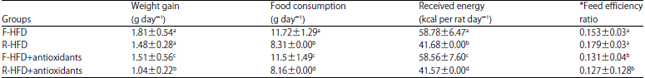 | |
*Feed efficiency ratio was determined as the ratio of body weight gained per unit (g) of feed consumed over a period of time (day), The values are shown as Mean±SE. Values with different superscripts within the column are significantly different (p<0.05) | |
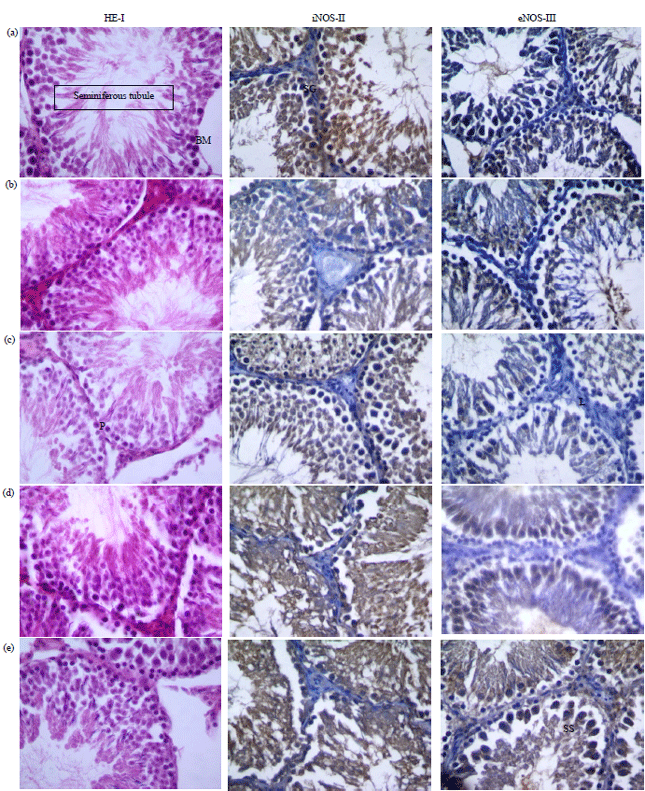 | |
| Fig. 1: | Photomicrographs of testis. HE-I: H and E staining showed intact basement membrane (BM), pycnotic nuclei cells (P) in the HFD+antioxidants group. iNOS-II and eNOS-III: iNOS and eNOS-III immunoreactivities, intense secondary spermatocytes (SS) and spermatids(S) in the R-HFD+antioxidants and the F-HFD+garlic groups, respectively. (a) F-HFD, (b) R-HFD, (C) F-HFD+antioxidants, (d) F-HFD+garlic and (e) R-HFD+antioxidants |
(3) Moderate immunoreactivity (++) was shown for eNOS in spermatocytes I of R-HFD, R-HFD+antioxidants and F-FHFD+garlic groups, spermatocytes II of F-HFD, R-HFD, F-FHFD+garlic and R-HFD+antioxidants groups, spermatids of F-HFD, R-HFD and R-HFD+antioxidants groups, (4) iNOS was moderate in spermatogonia of F-FHFD group, spermatocytes I of F-FHFD and R-HFD+antioxidants groups, spermatocytes II of R-HFD and F-FHFD+antioxidants and R-FHFD+antioxidants groups, spermatids of F-HFD+antioxidants and F-HFD+garlic groups.
| Table 5: | Seminiferous tubular diameter of rat after 12 weeks on the experimental diets |
 | |
| Values are shown as Mean±SE (p<0.05) | |
| Table 6: | Evaluation of staining intensity scores (by two observers blinded to the study information independently) |
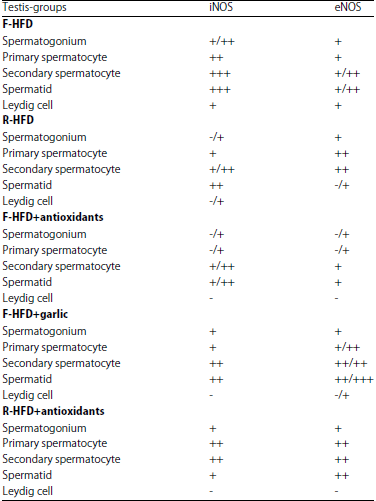 | |
| Intensity was graded as, -: Negative, +: Mild, ++: Moderate and +++: Intense | |
(5) Mild immunoreactivity (+) was shown for eNOS in spermatogonia of F-HFD, R-HFD, F-HFD+antioxidants, F-HFD+garlic and R-HFD+antioxidants groups, spermatocytes I of F-HFD, F-HFD+antioxidants groups, spermatocytes II of F-FHFD+antioxidants group, spermatids of F-HFD+antioxidants and R-HFD+antioxidants groups, leydig cells of F-HFD and F-HFD+garlic groups. (6) iNOS was mild in spermatogonia of R-HFD, F-HFD+antioxidants, F-HFD+garlic and R-HFD+antioxidants groups spermatocytes I of R-HFD, F-FHFD+antioxidants F-HFD+garlic groups, spermatids of R-HFD+antioxidants groups and leydig cells of F-HFD and R-HFD groups (7). The expression of eNOS was negative (-) in leydig cells of R-HFD, F-HFD+antioxidants and R-HFD+antioxidants groups. (8) iNOS was negative in leydig cells of F-HFD+antioxidants, R-HFD+antioxidants and F-HFD+garlic groups. There were two rats in R-HFD and one rat in F-HFD+antioxidants groups with hypogonadism. In the rats that were fed with HFD after 12 weeks, the evaluation of staining intensity scores is shown in Table 6 and differences between expression of iNOS and eNOS were compared in all experimental groups (Table 7). Expression rate of iNOS was compared with eNOS (Table 8).
DISCUSSION
The results of this study revealed that spermatogenesis function was disturbed in rats fed with a HFD, characterized as decreased body weight, decreased seminiferous tubule diameter, reduced spermatogenesis score and increased iNOS expression. Based on some evidence in the recent decades has of the world’s population, obesity can be attributed by environmental factors such as increase in the amount of fat in the diet, High Fat Diet (HFD), is the main cause of obesity3,4. Environmental factors, some diseases and different nutritive substances may directly or indirectly influence spermatogenesis.
| Table 7: | Comparison with expression of iNOS and eNOS intensity of rats that were fed with HFD after 12 weeks |
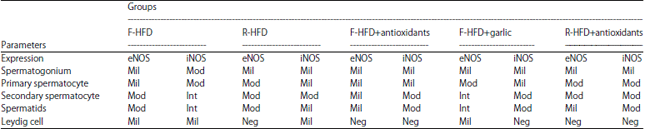 | |
| Intensity was graded as Neg: Negative, Mil: Mild (+), Mod: Moderate (++), Int: Intense (+++) | |
| Table 8: | Evaluation of expression rate of iNOS and eNOS in rat that were fed with HFD after 12 weeks |
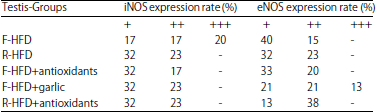 | |
Intensity was graded as, -: Negative, +: Mild, ++: Moderate and +++: Intense | |
In the present study, body weight increased in F-HFD more than R-HFD group. These results demonstrate the positive effect of calorie restriction on prevention of body weight raise and support the results obtained by Hall et al.26. Hariri et al.27, Park et al.28 and Mu et al.29 also observed overweight in animals fed with a high fat diet, regardless of the lipid quality. Unexpected results of present study showed that antioxidants treatment cannot improve increased body weight. These results were in disagreement with the Mu et al.29.
One of the parameters used for the evaluation of spermatogenesis is the seminiferous tubule diameter30. In present study, the seminiferous tubule diameter was lower in restricted HFD than the free HFD group. Campos-Silva et al.31 also reported that long-term administration diet rich in saturated fatty acids alters the testicular morphology with reductions of seminiferous tubule diameter and cell proliferation which could be related to a disturbance of spermatogenesis.
Antioxidants administration in the both F-HFD and R-HFD groups caused more reduction in seminiferous tubule diameter. With respect to the spermatogenic cell layers and mature sperms and spermatogenesis score RHFD+antioxidants and HFD+antioxidants groups also showed decrease compared with the R-HFD and F-HFD groups respectively, indicating a decrease in spermatozoa production. Some studies revealed the protective effect of antioxidants against sperm damage in rats fed with HFD which contradicts present results. It seems unexpected results regarding to antioxidents effect in the present study were due to that the effects of antioxidants on the oxidation of foods are dependent on their concentration32, polarity and the medium33,34 and also the presence of other antioxidants35. Moreover when there are 2 or more antioxidants together, interaction occurs such as synergism, antagonism and simple addition36. Results of Adewoyin et al.37 showed normal functioning of the spermatozoa may require a balance between ROS and antioxidant.
Nitric oxide synthases (NOS) isoforms producing nitric oxide (NO) that is a free radical involved in physiologic and pathologic interactions with ROS11. Expression of eNOS and iNOS showed in leydig cells, primary spermatocytes and spermatids by IHC method in the testis of pigs. Seemingly, these NOS isoforms contribute to the process of spermatogenesis14.
In the present study, eNOS immunoreactivity was mild in spermatogonia of F-HFD group. Moderate in primary spermatocytes and secondary spermatocytes of the R-HFD and RHD+antioxidants groups, spermatids in F-HFD, F-HFD+antioxidants and R-HFD+antioxidants, leydig cells of F-HFD+antioxidants and R-HFD+antioxidants groups, intense secondary spermatocytes and spermatids were in the R-HFD+antioxidants groups. It was shown that normal germ cells were not eNOS stained, it was detectable in some degenerating germ cells, also presence of eNOS protein was shown at all stages of spermatogenesis. The eNOS was localized in leydig cells, sertoli cells and degenerating or apoptotic intraepithelial germ cells. Moreover, eNOS overexpression happened in prematurely shed spermatocytes and spermatids14. Bayatli et al.38 showed that oxidative stress induced by torsion gives rise to serious damage in testis and increases in eNOS expression level. Grape seed proanthocyanidin extract as a potent antioxidant agent decreased eNOS expression and improved testicular morphology. Present study were agreed with these findings.
The results of iNOS immunoreactivity in this study were evaluated as following: mild in spermatogonia of F-HFD group, moderate in primary spermatocytes and secondary spermatocytes of the R-HFD and RHD+antioxidants groups, spermatids of F-HFD, F-HFD+antioxidants and R-HFD+antioxidants, leydig cells of F-HFD+antioxidants and R-HFD+antioxidants groups, intense in secondary spermatocytes and spermatids of the R-HFD+antioxidants group. The over expression of iNOS has reported in other tissues in the HF diet condition. Park et al.28 showed liver iNOS expression pattern in the HF group.
These findings were in accordance with Uzun et al.15 study that up-regulation of iNOS occurred in the damaged seminiferous epithelium resulting from LPS-induced inflammation. Atta et al.39 also reported that thymoquinone treatment exerted a protective effect against reproductive dysfunction induced by diabetes not only through its powerful antioxidant and hypoglycemic effects but also through its down regulation of testicular iNOS expression levels in diabetic rats.
The iNOS was a critical factor in harming spermatogenesis in adult rats16. In idiopathic varicocele, over expression of iNOS was observed in leydig cells17. The results of Asgharzade et al.40 study also showed that iNOS may play a functional role in spermatogenesis via apoptosis.
The present study showed that restricted HFD+antioxidants had higher disturbing effect on spermatogenesis than free HFD access. Restriction of high fat diet and adding antioxidants seems have not therapeutic effect for disturbing changes in testis. It is suggested to do research on basic causes of oxidative stress in the reproductive tract of male subjects and also prepare optimized antioxidants to treat conditions due to an imbalance in the redox status of testis. This could develop the knowledge about male fertility in obese men.
CONCLUSION
Results of this study revealed that restricted fat diet consumption caused increase in weight, although the restriction in high fat diets a beneficial way in weight loss. In this study impaired spermatogenesis could not improve by adding antioxidants. HFD could induce testicular iNOS expression. Since the intense iNOS was expressed in secondary spermatocytes and spermatids of seminiferous tubules in R-HFD+antioxidants group, confirms a potential role of iNOS in disturbing changes of spermatogenic layer and possible roles in male fertility.
SIGNIFICANCE STATEMENT
High and restricted fat diet consumption impairs spermatogenesis and could not improve by adding antioxidants. Over expression of iNOS in seminiferous tubules in restricted fat diet+antioxidants condition, confirms a potential role of iNOS in destructive changes of testis and its possible role in male fertility.
ACKNOWLEDGMENT
The authors are obliged to thank the Department of Histology and Embryology, School of Medicine, Celal Bayar University, Manisa, Turkey for IHC excellent technical assistance and Emad Sohrabi for statistical comparison among animal group. The study was funded by Vice-chancellor for Research and Technology, Hamadan University of Medical Sciences (grant No. 87071695658).
REFERENCES
- Magnusdottir, E.V., T. Thorsteinsson, S. Thorsteinsdottir, M. Heimisdottir and K. Olafsdottir, 2005. Persistent organochlorines, sedentary occupation, obesity and human male subfertility. Hum. Reprod., 20: 208-215.
CrossRefDirect Link - Lissner, L. and B.L. Heitman, 1995. Dietary fat and obesity: Evidence from epidemiology. Eur. J. Clin. Nutr., 49: 79-90.
PubMedDirect Link - Popkin, B.M. and C.M. Doak, 1998. The obesity epidemic is a worldwide phenomenon. Nutr. Rev., 56: 106-114.
CrossRefDirect Link - Brownlee, M., 2001. Biochemistry and molecular cell biology of diabetic complications. Nature, 414: 813-820.
CrossRefPubMedDirect Link - Pasquali, R., C. Pelusi, S. Genghini, M. Cacciari and A. Gambineri, 2003. Obesity and reproductive disorders in women. Hum. Reprod. Update, 9: 359-372.
CrossRefDirect Link - Agarwal, A., S. Gupta and S. Sikka, 2006. The role of free radicals and antioxidants in reproduction. Curr. Opin. Obst. Gynecol., 18: 325-332.
CrossRefPubMedDirect Link - Wang, Y., X.P. Liu, D.N. Qin, S. Chen and Y.S. Li, 2007. Diet-induced obesity affects testis development in pubertal rats. Nat. J. Androl., 13: 514-519.
PubMedDirect Link - Urakawa, H., A. Katsuki, Y. Sumida, E.C. Gabazza and S. Murashima et al., 2003. Oxidative stress is associated with adiposity and insulin resistance in men. J. Clin. Endocrinol. Metab., 88: 4673-4676.
CrossRefDirect Link - Campion, J., F.I. Milagro, D. Fernandez and J.A. Martinez, 2008. Vitamin C supplementation influences body fat mass and steroidogenesis-related genes when fed a high-fat diet. Int. J. Vitamin Nutr. Res., 78: 87-95.
CrossRefDirect Link - Sreekumar, R., J. Unnikrishnan, A. Fu, J. Nygren and K.R. Short et al., 2002. Impact of high-fat diet and antioxidant supplement on mitochondrial functions and gene transcripts in rat muscle. Am. J. Physiol. Endocrinol. Metab., 282: E1055-E1061.
CrossRefDirect Link - Audet, I., J.P. Laforest, G.P. Martineau and J.J. Matte, 2004. Effect of vitamin supplements on some aspects of performance, vitamin status and semen quality in boars. J. Anim. Sci., 82: 626-633.
PubMedDirect Link - Johnson, F.C. and H.M. Sinclair, 1979. The antioxidant vitamins. CRC Crit. Rev. Food Sci. Nutr., 11: 217-309.
CrossRefDirect Link - Ten, J., F.J. Vendrell, A. Cano and J.J. Tarin, 1997. Dietary antioxidant supplementation did not affect declining sperm function with age in the mouse but did increase head abnormalities and reduced sperm production. Reprod. Nutr. Dev., 37: 481-492.
Direct Link - Uzun, F.G., S. Kalender, D. Durak, F. Demir and Y. Kalender, 2009. Malathion-induced testicular toxicity in male rats and the protective effect of vitamins C and E. Food Chem. Toxicol., 47: 1903-1908.
CrossRefPubMedDirect Link - Borek, C., 1993. Molecular mechanisms in cancer induction and prevention. Environ. Health Perpect., 101: 237-245.
Direct Link - Nikravesh, M.R. and M. Jalali, 2010. The effect of crude aqueous extract of the Allium sativum on spermatogenesis in balb/C mice. Iran. J. Pharm. Res., 3: 58-58.
Direct Link - Hammami, I., A. Nahdi, C. Mauduit, M. Benahmed and M. Amri et al., 2008. The inhibitory effects on adult male reproductive functions of crude garlic (Allium sativum) feeding. Asian J. Androl., 10: 593-601.
CrossRefPubMedDirect Link - Saraswathi, V., G. Wu, M. Toborek and B. Hennig, 2004. Linoleic acid-induced endothelial activation role of calcium and peroxynitrite signaling. J. Lipid Res., 45: 794-804.
CrossRefDirect Link - Cheng, C.Y. and D.D. Mruk, 2010. The biology of spermatogenesis: The past, present and future. Philos. Trans. R. Soc. B: Biol. Sci., 365: 1459-1463.
CrossRefDirect Link - Hammoud, A.O., M. Gibson, C.M. Peterson, B.D. Hamilton and D.T. Carrell, 2006. Obesity and male reproductive potential. J. Androl., 27: 619-626.
CrossRefDirect Link - Holstein, A.F., W. Schulze and M. Davidoff, 2003. Understanding spermatogenesis is a prerequisite for treatment. Reprod. Biol. Endocrinol., Vol. 1.
CrossRef - Furnes, M.W., C.M. Zhao and D. Chen, 2009. Development of obesity is associated with increased calories per meal rather than per day. A study of high-fat diet-induced obesity in young rats. Obesity Surg., Vol. 19.
CrossRef - Sohrabi, M., A.M. Roushandeh, Z. Alizadeh, A. Vahidinia, M. Vahabian and M. Hosseini, 2015. Effect of a high fat diet on ovary morphology, in vitro development, in vitro fertilisation rate and oocyte quality in mice. Singapore Med. J., 56: 573-579.
Direct Link - Hall, K.D., T. Bemis, R. Brychta, K.Y. Chen and A. Courville et al., 2015. Calorie for calorie, dietary fat restriction results in more body fat loss than carbohydrate restriction in people with obesity. Cell Metab., 22: 427-436.
CrossRefDirect Link - Hariri, N., R. Gougeon and L. Thibault, 2010. A highly saturated fat-rich diet is more obesogenic than diets with lower saturated fat content. Nut. Res., 30: 632-643.
CrossRefDirect Link - Park, S., N.Y. Park, G. Valacchi and Y. Lim, 2012. Calorie restriction with a high fat diet effectively attenuated inflammatory response and oxidative stress related markers in obese tissues of the high diet fed rats. Mediators Inflamm.
CrossRefDirect Link - Mu, Y., W.J. Yan, T.L. Yin and J. Yang, 2016. Curcumin ameliorates high‑fat diet‑induced spermatogenesis dysfunction. Mol. Med. Rep., 14: 3588-3594.
CrossRefDirect Link - Volkmann, J., D. Muller, C. Feuerstacke, S. Kliesch, M. Bergmann, C. Muhlfeld and R. Middendorff, 2011. Disturbed spermatogenesis associated with thickened lamina propria of seminiferous tubules is not caused by dedifferentiation of myofibroblasts. Hum. Reprod., 26: 1450-1461.
CrossRefDirect Link - Campos-Silva, P., A. Furriel, W.S. Costa, F.J. Sampaio and B.M. Gregorio, 2015. Metabolic and testicular effects of the long-term administration of different high-fat diets in adult rats. Int. Braz. J. Urol., 41: 569-575.
CrossRefDirect Link - Frankel, E.N., S.W. Huang, E. Prior and R. Aeschbach, 1996. Evaluation of antioxidant activity of rosemary extracts, carnosol and carnosic acid in bulk vegetable oils and fish oil and their emulsions. J. Sci. Food Agric., 72: 201-208.
CrossRefDirect Link - Cheng, Z., J. Ren, Y. Li, W. Chang and Z. Chen, 2003. Establishment of a quantitative structure-activity relationship model for evaluating and predicting the protective potentials of phenolic antioxidants on lipid peroxidation. J. Pharm. Sci., 92: 475-484.
CrossRefDirect Link - Samotyja, U. and M. Malecka, 2007. Effects of blackcurrant seeds and rosemary extracts on oxidative stability of bulk and emulsified lipid substrates. Food Chem., 104: 317-323.
CrossRefDirect Link - Perron, N.R. and J.L. Brumaghim, 2009. A review of the antioxidant mechanisms of polyphenol compounds related to iron binding. Cell Biochem. Biophys., 53: 75-100.
CrossRefPubMedDirect Link - Choe, E. and D.B. Min, 2009. Mechanisms of antioxidants in the oxidation of foods. Comprehensive Rev. Food Sci. Food Saf., 8: 345-358.
CrossRefDirect Link - Adewoyin, M., M. Ibrahim, R. Roszaman, M.L.M. Isa, N.A.M. Alewi, A.A.A. Rafa and M.N.N. Anuar, 2017. Male infertility: The effect of natural antioxidants and phytocompounds on seminal oxidative stress. Diseases, Vol. 5.
CrossRef - Bayatli, F., D. Akkus, E. Kilic, R. Saraymen and M.F. Sonmez, 2013. The protective effects of grape seed extract on MDA, AOPP, apoptosis and eNOS expression in testicular torsion: An experimental study. World J. Urol., 31: 615-622.
CrossRefDirect Link - Atta, M.S., E.A. Almadaly, A.H. El-Far, R.M. Saleh, D.H. Assar, S.K. Al Jaouni and S.A. Mousa, 2017. Thymoquinone defeats diabetes-induced testicular damage in rats targeting antioxidant, inflammatory and aromatase expression. Int. J. Mol. Sci., Vol. 18.
CrossRef - Asgharzade, S., M. Rafieian-Kopaei, A. Mirzaeian, S. Reiisi and L. Salimzadeh, 2015. Aloe vera toxic effects: Expression of inducible nitric oxide synthase (iNOS) in testis of Wistar rat. Iran. J. Basic Med. Sci., 18: 967-973.
Direct Link