
Khayalethu Ntushelo
Department of Agriculture and Animal Health, Science Campus, University of South Africa, Corner Christian De Wet and Pioneer Avenue, Private Bag X6, 1710 Florida, South Africa
LiveDNA: 27.15205
Pakistan Journal of Biological Sciences
Year: 2017 | Volume: 20 | Issue: 6 | Page No.: 278-288







ABSTRACT
Background and Objective: Salicylic acid is a signal molecule which activates plant defense against plant pathogens such as the soft rot enterobacterium Pectobacterium carotovorum subsp. carotovorum. The objectives of study were to determine bactericidal effects of salicylic acid on the growth of P. carotovorum subsp. carotovorum and secondly, assess chemical responses of P. carotovorum subsp. carotovorum to salicylic acid. Materials and Methods: Pectobacterium carotovorum subsp. carotovorum was grown in lysogeny broth amended with salicylic acid at concentrations of 0, 100, 200, 400, 800 and 1200 mg L–1. The P. carotovorum subsp. carotovorum cultures were incubated at 25°C and sampled at two time points, 0 h (sampled before incubation) and 24 h. Bacterial counts were done at the onset of the incubation (0 h) and after the 24 h incubation. The set which was incubated for 24 h was split into two, one subset was centrifuged and the other was not. From the centrifuged subset the supernatant was recovered and was, together with all the other samples (0 and 24 h not centrifuged), analyzed with1H nuclear magnetic resonance and gas chromatography. Results: Bacterial counts done before and after incubation showed that the lower concentrations of salicylic acid, 0, 100, 200 and 400 mg L–1, supported the growth of P. carotovorum subsp. carotovorum whereas the higher concentrations of 800 and 1200 mg L–1 inhibited the growth of the bacterium completely. Nuclear magnetic resonance results showed either slight or no differences in the metabolite profiles and gas chromatography showed different responses without a clearly defined pattern among the experimental treatments. However, methanethiol was detected by both nuclear magnetic resonance and gas chromatography in all the treatments and was probably formed as a result of the breakdown of lysogeny broth. Conclusion: From the results obtained it was concluded that salicylic acid promotes the growth of P. carotovorum subsp. carotovorum at lower concentrations of 0-400 mg L–1 but higher concentrations of salicylic acid of 800 and 1200 mg L–1 inhibit bacterial growth. All the tested salicylic acid concentrations (0-1200 mg L–1) cause only slight chemical shifts in the bacterial culture. Methanethiol was detected in all treatments and it is probably formed from the breakdown of lysogeny broth.
PDF Abstract XML References Citation
Received: February 16, 2017;
Accepted: April 06, 2017;
Published: May 15, 2017
Copyright: © 2017. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Khayalethu Ntushelo, 2017. Effect of Salicylic Acid on the Growth and Chemical Responses of Pectobacterium carotovorum subsp. carotovorum. Pakistan Journal of Biological Sciences, 20: 278-288.
DOI: 10.3923/pjbs.2017.278.288
URL: https://scialert.net/abstract/?doi=pjbs.2017.278.288
DOI: 10.3923/pjbs.2017.278.288
URL: https://scialert.net/abstract/?doi=pjbs.2017.278.288
INTRODUCTION
Bacteria in liquid cultures grow sigmoidally in three phases, the lag phase, the exponential phase and the equilibrium phase1-4. Liquid culture dynamics range from depletion of nutrients, accumulation of toxic metabolites and changes in ion equilibrium. It is assumed that bacteria in liquid cultures grow and produce adaptive compounds in response to their chemical environment. For the purposes of plant-microbe interactions, aspects of importance are the response of plant pathogens to compounds produced during pathogen invasion. Salicylic Acid (SA) is one of these compounds produced by the plant in its defense against pathogens5-9. Many studies have shown that this plant-produced phenolic compound serves as a key signal molecule for activating Pathogen-Associated Molecular Pattern (PAMP)-triggered immunity (PTI), Effector-Triggered Immunity (ETI) and Systemic Acquired Resistance (SAR) following infection by pathogens10,11. Because of its role in plant defense, studies have been conducted to investigate the role of exogenously applied SA against various pathogens. Czajkowski et al.12 found SA to inhibit infection by Dickeya solani of potato. Lagonenko et al.13 found that SA inhibits biofilm formation, motility and N-Acyl homoserine lactone production by Pectobacterium carotovorum and Pseudomonas syringae pv. syringae at sub inhibitory concentrations. Furthermore, it was found that SA induces biofilm formation by Pseudomonas corrugata while inhibiting its motility and had no effect on biofilm formation by Xanthomonas campestris pv. campestris but slightly increased its motility. In this same study, Erwinia amylovora was insensitive to SA treatment. Chow et al.14 found SA to affect motility of P. aeruginosa and Da et al.15 found SA to attenuate biofilm formation but not swarming in P. aeruginosa. Prithiviraj et al.16 found it to attenuate the virulence of P. aeruginosa on Arabidopsis thaliana as well as decrease gene expression of the bacterial fliC genes at growth inhibitory levels. Although these studies have generally shown the inhibitory effects of SA on plant pathogenic bacteria some critical aspects in this knowledge realm have been left out. Effect of SA on the growth of many plant pathogenic bacteria remains unknown and not much investigation has been done on chemical responses of bacteria exposed to SA. Prior to undertaking this study, two areas requiring investigation were identified. Firstly, effect of SA on the growth of the soft rot enterobacterium Pectobacterium carotovorum subsp. carotovorum (Pcc) has not been investigated. Secondly, it is not known if exposure to SA of Pcc causes chemical responses by this enterobacterium. The objectives of this study were therefore to assess the growth of Pcc exposed to different concentrations of SA as well as investigate chemical responses of the bacterium to exposure to SA. As a plant defence signalling compound and plant pathogen, SA and Pcc naturally come into contact during plant-pathogen interactions and therefore their interaction warrants investigation.
MATERIALS AND METHODS
Assessment of bacterial growth in culture amended with salicylic acid: Thirty milliliters batches of lysogeny broth with concentrations of 0, 100, 200, 400, 800 and 1200 mg L–1 of salicylic acid were prepared. Into each of the six broth preparations a 1 mL culture with approximately 4×106 Pectobacterium carotovorum subsp. carotovorum (Pcc) cells was added. The experiment was replicated six times and the cultures were incubated at 25°C for 24 h. Another set of the cultures was frozen immediately after inoculation and was analysed as the time 0 h set which was compared with the set which was incubated for 24 h. After 24 h of incubation, bacterial counts were performed by plating or serial diluted culture samples. After the counts half of the treatments were centrifuged at 14000 rpm, the supernatant was recovered and kept for further analysis. With the remaining half of the treatments, complete culture contents were saved for further analysis. Details on the treatments are shown in Table 1.
1H nuclear magnetic resonance spectroscopy: 1H Nuclear Magnetic Resonance (NMR) spectroscopy was conducted to assess if Salicylic Acid (SA) causes Pcc to produce response metabolites in the lysogeny broth Pcc cultures which were amended with SA. This would be observed by chemical shifts in the various cultures which had different concentrations of SA. Secondary metabolites were extracted from 50 μL of the cultures of the various treatments. Information on samples and treatments is shown in Table 1. Secondary metabolites were extracted by adding 600 μL methanol-D4 and 600 μL buffer (deuterium oxide + potassium dihydrogen phosphate). The mixture was vortexed, sonicated for 20 min and centrifuged briefly at room temperature. The supernatant was dispensed into NMR tubes for analysis. Nuclear magnetic resonance spectroscopic analysis was done using a 600 MHz Varian NMR instrument to obtain 1D proton spectra of the samples.
| Table 1: | Grouping of samples by similarity of *1H nuclear magnetic resonance (NMR) spectra |
 | |
*Only slight differences in NMR spectra were caused by the salicylic acid treatments. Type of media did not influence secondary metabolite profiles so was time of sampling, Samples with the same letter (a, b, c, d, e, f, g) had similar NMR spectra | |
Phase correction and baseline correction of the spectra was done using ACD/NMR Processor. The NMR shift range visually inspected for spectral differences was between 0.5 and 4 ppm (Fig. 1) but only the range of the spectra from 0 to 6 ppm was bucketed for further analysis. From the bucketed range principal component analysis was done using SIMCA to ascertain treatment differences. Nuclear magnetic resonance spectra were queried on NMR Shift DB to identify the metabolites in the treatment samples. For each sample analysed with NMR, NMR Shift DB generated an output of 300 compounds represented by the spectrum and these compounds were matched with compound identities from the output of the gas chromatography analysis.
Gas chromatography analysis: Gas chromatography analysis was conducted to assess if SA causes Pcc to produce response metabolites (volatile organic compounds) in the Pcc lysogeny broth cultures which had different concentrations of SA. This would be observed by differences in the gas chromatography spectra in the various cultures which had different concentrations of SA. The list of samples is shown is Table 1, however, only supernatant solutions and no whole cell cultures were included in the gas chromatography analysis. When gas chromatography was performed the total growth inhibiting 1200 mg L–1 salicylic acid treatment was omitted because it was not expected to produce results different from the 800 mg L–1 treatment which also completely inhibited the growth of Pectobacterium carotovorum subsp. carotovorum cells. Small volumes (10 μL) of the supernatant samples were dispensed into gas chromatography vials. The samples were deproteinized by adding 40 μL of methanol and subsequently centrifuged for 10 min at 12000 g. The supernatant was dried under N2 and then derivatized with hydroxylamine chloride for 90 min at 40°C and MSTFA for 50 min at 40°C. The samples were injected into the gas chromatogram for analysis. Measurements of retention time were taken and similarity scores based on entries in the database were recorded for identification of the volatile compounds.
 | |
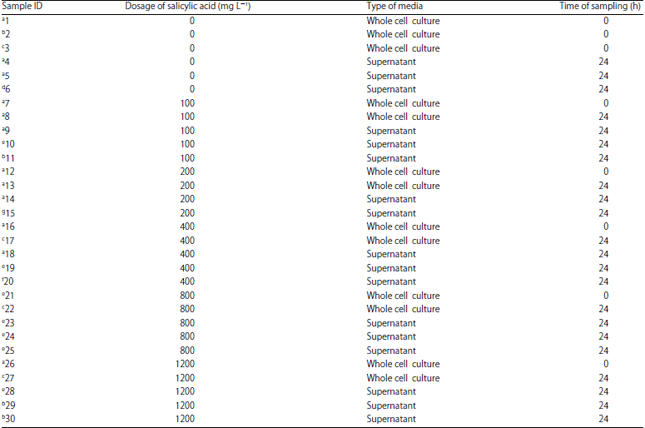
| Fig. 1: | 1H nuclear magnetic resonance (NMR) spectra of representative samples tested in the NMR analysis |
| The box is drawn to show some of the differences in the spectra | |
RESULTS
Assessment of bacterial growth in culture amended with salicylic acid: The assays were conducted in 50 mL tubes with a volume of 31 mL broth. The initial Pectobacterium carotovorum subsp. carotovorum (Pcc) bacterial count was approximately 4×106 per treatment. The assay lasted for 24 h after which final bacterial counts were conducted in each of the treatments.
 | |
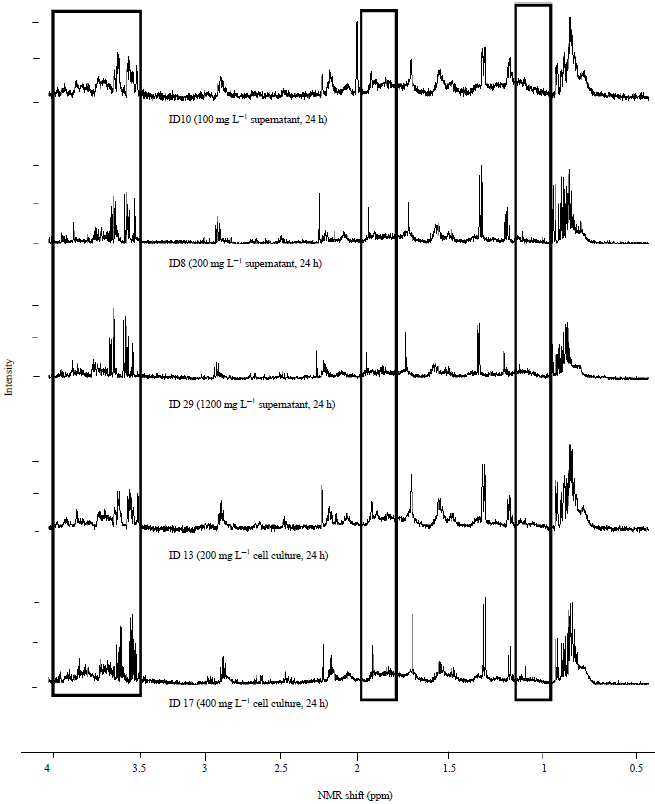
| Fig. 2: | Principal component analysis of all the observations in Table 1 |
The observations are labelled by the dose of salicylic acid (0, 100, 200, 400, 800 and1200 mg L–1). The Q2 value was 0.657, PCAReadyPccData9Until6.0-Copy.M1 (PCA-X), Colored according to Obs ID (Dose (mg L–1)) | |
The counts were as follows after the 24 h incubation: The treatment with 0 mg L–1 Salicylic Acid (SA) had a total count of approximately 3×109 bacterial cells, the treatment with 100 mg L–1 of SA also had approximately 3×109 cells, the count with 200 mg L–1 SA had approximately 4×109 cells, the treatment with 400 mg L–1 had about 1×109 bacterial cells and no bacterial growth was recorded in the 800 and 1200 mg L–1 treatments. The pH was seven for the initial media except for 800 and 1200 mg which had a pH slightly lower than seven. The pH at termination was also seven except for 400 and 800 mg L–1 samples which had a pH of six as well as 1200 mg L–1 sample which had a pH of 4. The pH changes did not form part of the study but pH was tested just to assess if the culture changed.
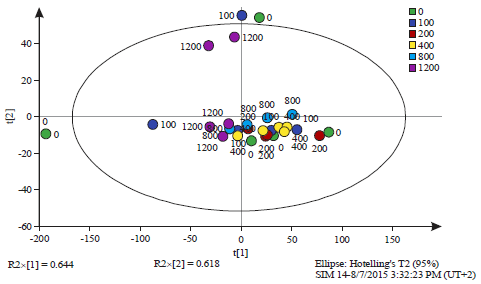
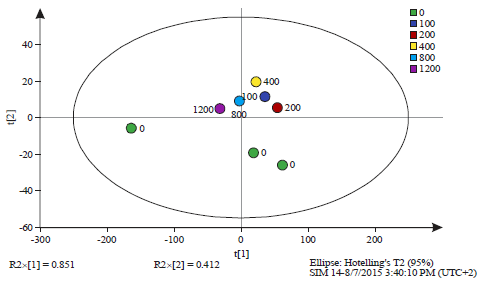
Analysis of 1H nuclear magnetic resonance data: When the NMR spectra was closely inspected some minor differences were observed between spectra according to the experimental treatments. The NMR spectra showed that the alkyl region 0.7-1.3 ppm was slightly different between the spectra. The methyl ketone region (2-2.4 ppm) also differed slightly as well as the alcohol ether region (3.5-4.5 ppm) (Fig. 1). The samples representing these differences were the supernatant of the dosage 100 mg L–1 of Salicylic Acid (SA) after 24 h of incubation (Sample ID 10), the supernatant of the dosage of 200 mg L–1 of SA after 24 h of incubation (Sample ID 8), the cell culture of the dosage of 1200 mg L–1 after 24 h of incubation (Sample ID 29), the cell culture of the dosage of 200 mg L–1 after 24 h of incubation (Sample ID 13) and the cell culture of the dosage of 400 mg L–1 after 24 h of incubation (Sample ID 17) (Fig. 1). Further comparison of samples based on NMR spectral differences is illustrated in Table 1. Principal Component Analysis (PCA) was not able to clearly discriminate between the samples (Fig. 2-6). However, when all the samples which were taken at time 0 h (before incubation) were analysed separately some grouping was evident. At 0 h sampling time point the difference between the samples was the amount of SA added as the bacterium had not had sufficient time in the culture to produce response chemicals. At the 0 h time point the 0 mg L–1 SA treatment samples were grouped together by PCA and the rest of the samples formed another single but distinct cluster (Fig. 3). This was not the case with the results of the separate analysis of samples taken at the 24 h sample point. No discrimination was evident among the samples (Fig. 4). The reasons for lack of discrimination amount the 24 h sample point could not be understood by just analyzing the data generated in this study and therefore additional experimentation and expanded analysis of the cultures are necessary. However, it could be that cell exudates produced during culturing and chemical products from the breakdown of lysogeny broth mask the PCA discrimination which was evident with just differences in SA concentration at time 0 h. Similarly, PCA was not able to discriminate among whole cell cultures sampled at both time 0 h and time 24 h (Fig. 5) and the reasons for this lack of discrimination could not be ascertained from analysing data generated from just this study. There was also no discrimination of supernatant samples taken at both the time 0 h and the time 24 h sampling points (Fig. 6).
 | |
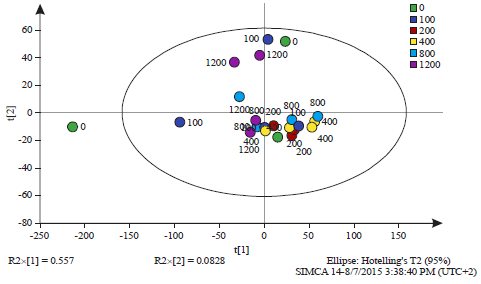
| Fig. 3: | :Principal component analysis of all observations at 0 h time interval (Table 1) |
The observations are labelled by the dose of salicylic acid (0, 100, 200, 400, 800 and 1200 mg L–1). The Q2 value was 0.793, PCAReadyPccData5Until6.0 ZeroHoursOnly.M1 (PCA-X), Colored according to Obs ID (Dose (mg L–1)) | |
 | |
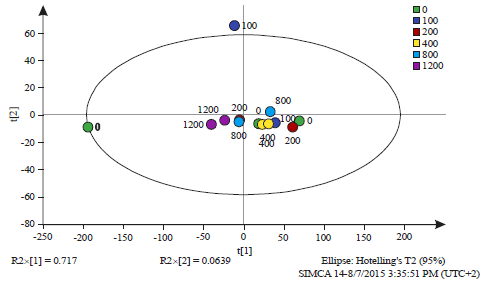
| Fig. 4: | Principal component analysis of all observations at the 24 h time interval (Table 1) |
The observations are labelled by the dose of salicylic acid (0, 100, 200, 400, 800 and 1200 mg L–1). The Q2 value was 0.546, PCAReadyPccData2Until6.0 24 h.M1 (PCA-X), Colored according to Obs ID (Dose (mg L–1)) | |
Gas chromatography: The production of volatile compounds by Pcc was expected because bacteria are known to produce volatile compounds17-19. In this study it could be ascertained that the volatile compounds detected uniquely in the different experimental treatments are produced by the bacterium and are not part of the medium since a different set of unique volatile compounds were found in each treatment (Fig. 7). The most frequently emitted volatile compounds by bacteria are, among others, furfural, butanoic acid, propanoic acid, 5-hydroxy-methyl-furfural, β-caryophyllene, geomycin, 2-methyl isoborneol, 1-octen-3-ol, α-pinene, camphene, camphor, methanol and actetyldehyde20-31. Of these, butanoic acid, was emitted in all the experimental treatments, propanoic acid by all the treatments except for 800 mg L–1 and acetaldehyde was emitted only in the 400 mg L–1 treatment. It could be assumed that propanoic acid was produced by the bacterium since it was only the culture (800 mg L–1) which did not support any bacterial growth which did not have propanoic acid. Unlike in the NMR analysis where a slight discrimination between lower salicylic acid (SA) dosages and higher dosages was observed on the spectra (Fig. 1, Table 1) no observable pattern could be seen in the spectra of volatile compounds produced in the different treatments.
 | |
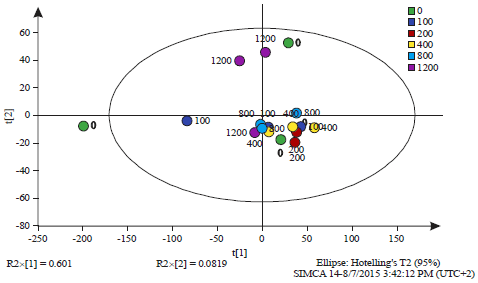
| Fig. 5: | Principal component analysis of all observations of whole cell cultures (Table 1) |
The observations are labelled by the dose of salicylic acid (0, 100, 200, 400, 800 and 1200 mg L–1). The Q2 value was 0.609, PCAReadyPccData3Until6.0 cells.M1 (PCA-X), Colored according to Obs ID (Dose (mg L–1)) | |
 | |
| Fig. 6: | Principal component analysis of all observations of the supernatant (Table 1) |
The observations are labelled by the dose of salicylic acid (0, 100, 200, 400, 800 and 1200 mg L–1). The Q2 value was 0.531, PCAReadyPccDataUntil6.0 Supernatant1-Copy.M1 (PCA-X), Colored according to Obs ID (Dose (mg L–1)) | |
However, the gas chromatography spectra were unique for each experimental treatment, to indicate that a unique set of volatile compounds is produced by Pcc in response to exposure to SA. Salicylic acid is produced by the plants during pathogen invasion. It is obvious that different volumes of SA may be produced by the plant and therefore the bacterial pathogen may produce different response secondary metabolites and volatile organic compounds as has been demonstrated by this study. The quantification of compounds produced during the plant-Pcc interaction falls outside the scope of this study and therefore it is not known how much SA concentration change is necessary to initiate a change in the profile of secondary metabolites and volatile organic compounds produced by Pcc. Methanethiol (CH4S) was detected in all the treatments and was probably produced as a result of the degradation of the lysogeny broth. It was detected with a similarity of 97.5% by NMR Shift DB and was also detected by gas chromatography. Out of the compounds detected either by gas chromatography or NMR it could be ascertained that methanethiol was found in those samples since it was detected by both NMR and gas chromatography.
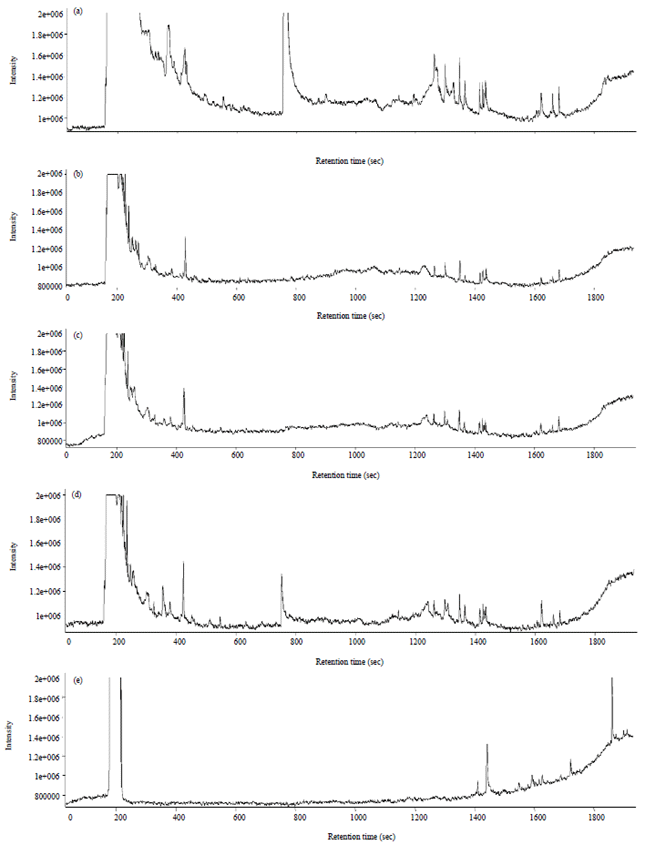 | |
| Fig. 7: | Gas chromatography spectra of supernatant solutions of bacterial cultures grown in different concentrations [(a) 800, (b) 400, (c) 200, (d) 100 and (E) 0 mg L–1] of salicylic acid |
DISCUSSION
Salicylic Acid (SA) promoted the growth of Pectobacterium carotovorum subsp. carotovorum (Pcc) at lower concentrations (0, 100, 200, 400 mg L–1) but higher concentrations of SA (800 and 1200 mg L–1) inhibited the growth of this bacterium. Only slight shifts in both nuclear magnetic resonance and gas chromatography profiles occurred when this bacterium was exposed to concentrations up to 1200 mg L–1. Methanethiol was found in all the cultures (all the treatments) and it was probably formed from the breakdown of lysogeny broth. Concentrations of SA chosen were 0, 100, 200, 400, 800 and 1200 mg L–1 and this selection was guided by the study of Wu et al.32, who found that biomass, colony diameter, number of conidium germination and conidium production of Fusarium oxysporum f. sp. niveum were decreased by 52.0, 25.7, 100 and 100% at concentrations of 800 mg L–1 and it was therefore estimated that, for the purposes of this study, the effective concentration against Pcc would not exceed 1200 mg L–1. That lower concentrations of SA (0, 100, 200, 400 mg L–1) supported the growth of Pcc and higher concentrations (800 and 1200 mg L–1) inhibited bacterial growth was not surprising but it was expected that there would also be chemical shifts (detected either by NMR or gas chromatography or both) due to exposure to SA. The finding that SA inhibits the growth of Pcc was concurrent with previous findings12-16. However, inhibition of growth occurred only in the two highest concentrations of 800 and 1200 mg L–1 and Pcc cells multiplied at the lower concentrations during the 25°C incubation over 24 h. The lower concentrations stimulated Pcc growth in a near-dose dependant manner. The boost of growth of Pcc by the lower concentrations of SA was probably due to the use of SA as a carbon source. Bacteria have the catechol and gentisate pathways as part of their metabolism and use these pathways to degrade SA33. It was therefore assumed that Pcc employed these pathways to support its growth in the lower dosages of SA. Furthermore, SA has another nutritive function in bacteria, it facilitates iron incorporation into iron depleted bacterial cells34 and this nutritional function must have been utilised by Pcc at the lower concentrations of SA. The results of this study showed very little chemical modification of the culture environment by the Pcc. Both NMR and gas chromatography profiles did not detect major shifts between Pcc treated with the various doses of SA whether before incubation or after and whether the supernatant or whole culture cells were analysed. From the observed NMR data, it was concluded that bacterial cells and SA have very little contribution to the chemical composition of the culture with little influence on the metabolite shifts. Only 0 mg L–1 concentration of SA was shown to differ from the other concentrations by Principal Component Analysis (PCA) of NMR intensity values (Fig. 3). The difference between the samples analysed by PCA in Fig. 3 was the concentration of SA. Microbial metabolomic procedures aim at quantifying microbial substrates and products either outside the cells (extracellular) or inside the cells (intracellular). For the present study only excreted metabolites could be detected because the cells were not lysed before chemical analysis and therefore any within-cell changes induced by SA could not be detected. Because this study aimed at understanding bacterial cell responses when Pcc is inside the plant, it was necessary not to lyse the cells as it was assumed that at no point bacterial cells are lysed during plant invasion and proliferation within the plant. Generally, extracellular metabolites are quantified in the cell free supernatant obtained either by filtration or centrifugation at low temperatures. This was the first set for analysis in this study and in the other set no centrifugation to recover the supernatant was done and the culture with the bacterial cells suspended was analysed (Table 1). This was the nearest point at which the endo metabolome (the total complement of metabolites inside bacterial cells) could be understood. Lack of detection of chemical changes in the Pcc cultures treated with different dosages of SA could be as a result of the low level of sensitivity of NMR and the fact that the volatile compounds (and compounds which could be derivatized) were mostly those in the growth medium and not as a result of response compounds produced by the bacterium. Holistic analytical metabolomics platforms must therefore be supplemented with more advanced hyphenated techniques to delineate even minute components of the culture. However, a basic gas chromatography instrument was able to provide analysis of what was the requirement in this study. Gas chromatography was suitable for use to achieve the aims of this study based on its validation done in a previous study35. To test the suitability of gas chromatography for analysis of bacterial cultures an extensive study was done using different microorganisms, i.e., Bacillus subtilis, Propionibacterium freudenreichii and Escherichia coli35. Escherichia coli was analysed at different growth stages of the culture to assess detection of culture differences. Many metabolite classes could be analyzed, alcohols, aldehydes, amino acids, amines, fatty acids, (phospho-) organic acids, sugars, sugar acids, (acyl-) sugar amines, sugar phosphate, purines, pyrimidines and aromatic compounds. The derivatization reaction proved to be efficient with more than 50% of compounds transferred to the volatile state. Linearity for most metabolites was satisfactory with regression coefficients better than 0.996 and the sensitivity was satisfactory with quantification limits at 40-500 pg on-column or 0.1-0.7 mmol g–1 of microbial cells (dry weight). Generally, intrabatch precision (repeatability) and inter batch precision (reproducibility) for the analysis of metabolites in cell extracts was better than 10 and 15%, respectively35. The GC has proven to be advantageous with its high sensitivity, the simultaneous quantification of many different metabolites (glycolysis, tricarboxylic acid cycle and pentose phosphate pathway) and the small sample volumes (10 μL) required for analysis with a detection limit in the picomole range36-38. This reliability of the GC supported the use of this instrument for analysing the cultures of Pcc which were amended with SA to produce different concentrations of this plant hormone in the culture. With the GC analysis it was possible to detect methanethiol which was also detected by NMR. Detection of methanethiol by the two detection methods improved the confidence that this compound was present in the cultures. Because methanethiol was present in all the cultures analysed by GC (all supernatant cultures) it could therefore be concluded that it was formed from the breaking down of lysogeny broth. To conclude, lower concentrations of SA support the growth of Pcc and higher concentrations of 800 and 1200 mg L–1 of SA completely inhibit the growth of Pcc. Very little chemical response was induced by the tested concentrations of SA on Pcc.
CONCLUSION
It is concluded that salicylic acid promotes the growth of Pectobacterium carotovorum subsp. carotovorum at lower concentrations but at higher concentrations of SA inhibit growth of this bacterium. Only slight shifts in both nuclear magnetic resonance and gas chromatography profiles occur when this bacterium is exposed to concentrations up to 1200 mg L–1. Methanethiol is a common compound in lysogeny broth cultures of P. carotovorum subsp. carotovorum and it is probably formed from the breakdown of lysogeny broth. With these observations it could also be concluded that plants which produce high levels of SA may inhibit the growth of a bacterial pathogen like P. carotovorum subsp. carotovorum.
SIGNIFICANCE STATEMENTS
This study provided insights into the fate of Pectobacterium carotovorum subsp. carotovorum (Pcc) when it comes to contact with Salicylic Acid (SA) in the plant during plant invasion. Plants equipped with production of SA to higher concentrations may be able to ward of Pcc. In addition to its role as a defence signalling molecule, SA may also play the role of a bactericide in the plant.
ACKNOWLEDGMENTS
I thank Ms. Mpho Alice Kotoane for assistance with gas chromatography analysis. Financial assistance was provided by the University of South Africa under the Emerging Researchers Support Programme (grant number 354500).
REFERENCES
- Gompertz, B., 1825. On the nature of the function expressive of the law of human mortality and on a new mode of determining the value of life contingencies. Philos. Trans. R. Soc. London, 115: 513-585.
CrossRefDirect Link - Richard, F.J., 1959. A flexible growth function for empirical use. J. Exp. Biot., 10: 290-300.
CrossRefDirect Link - Schnute, J., 1981. A versatile growth model with statistically stable parameters. Can. J. Fish. Aquat. Sci., 38: 1128-1140.
CrossRefDirect Link - Stannard, C.J., A.P. Williams and P.A. Gibbs, 1985. Temperature/growth relationships for psychrotrophic food-spoilage bacteria. Food Microbiol., 2: 115-122.
CrossRefDirect Link - Chen, Z., H. Silva and D.F. Klessig, 1993. Active oxygen species in the induction of plant systemic acquired resistance by salicylic acid. Science, 262: 1883-1886.
CrossRefDirect Link - Rasmussen, J.B., R. Hammerschmidt and M.N. Zook, 1991. Systemic induction of salicylic acid accumulation in cucumber after inoculation with Pseudomonas syringae pv syringae. Plant Physiol., 97: 1342-1347.
CrossRefDirect Link - Malamy, J., J.P. Carr, D.F. Kessig and I. Raskin, 1990. Salicylic acid: A likely endogenous signal in the resistance response of tobacco to viral infection. Science, 250: 1002-1004.
CrossRefDirect Link - Metraux, J.P., H. Signer, J. Ryals, E. Ward and M. Wyss-Benz et al., 1990. Increase in salicylic acid at the onset of systemic acquired resistance in cucumber. Science, 250: 1004-1006.
CrossRefPubMedDirect Link - Mills, P.R. and R.K.S. Wood, 1984. The effects of polyacrylic acid, acetylsalicylic acid and salicylic acid on resistance of cucumber to Colletotrichum lagenarium. J. Phytopathol., 111: 209-216.
CrossRefDirect Link - An, C. and Z. Mou, 2011. Salicylic acid and its function in plant immunity. J. Integr. Plant Biol., 53: 412-428.
CrossRefDirect Link - Vlot, A.C., D.M.A. Dempsey and D.F. Klessig, 2009. Salicylic acid, a multifaceted hormone to combat disease. Annu. Rev. Phytopathol., 47: 177-206.
CrossRefDirect Link - Czajkowski, R., J.M. van der Wolf, A. Krolicka, Z. Ozymko, M. Narajczyk, N. Kaczynska and E. Lojkowska, 2015. Salicylic acid can reduce infection symptoms caused by Dickeya solani in tissue culture grown potato (Solanum tuberosum L.) plants. Eur. J. Plant Pathol., 141: 545-558.
CrossRefDirect Link - Lagonenko, L., A. Lagonenko and A. Evtushenkov, 2013. Impact of salicylic acid on biofilm formation by plant pathogenic bacteria. J. Biol. Earth Sci., 3: B176-B181.
Direct Link - Chow, S., K. Gu, L. Jiang and A. Nassour, 2011. Salicylic acid affects swimming, twitching and swarming motility in Pseudomonas aeruginosa, resulting in decreased biofilm formation. J. Exp. Microbiol. Immunol., 15: 22-29.
Direct Link - Da, M.L., A.K. Heroux, Z. Pakzad and K.F.E.S. Schiffmacher, 2010. Salicylic acid attenuates biofilm formation but not swarming in Pseudomonas aeruginosa. J. Exp. Microbiol. Immunol., 14: 69-73.
Direct Link - Prithiviraj, B., H.P. Bais, T. Weir, B. Suresh and E.H. Najarro et al., 2005. Down regulation of virulence factors of Pseudomonas aeruginosa by salicylic acid attenuates its virulence on Arabidopsis thaliana and Caenorhabditis elegans. Infect. Immun., 73: 5319-5328.
CrossRefPubMedDirect Link - Schulz, S. and J.S. Dickschat, 2007. Bacterial volatiles: The smell of small organisms. Nat. Prod. Rep., 24: 814-842.
CrossRefPubMedDirect Link - Kai, M., M. Haustein, F. Molina, A. Petri, B. Scholz and B. Piechulla, 2009. Bacterial volatiles and their action potential. Applied Microbiol. Biotechnol., 81: 1001-1012.
CrossRefDirect Link - Lemfack, M.C., J. Nickel, M. Dunkel, R. Preissner and B. Piechulla, 2014. mVOC: A database of microbial volatiles. Nucleic Acids Res., 42: D744-D748.
CrossRefDirect Link - Stahl, P.D. and T.B. Parkin, 1996. Microbial production of volatile organic compounds in soil microcosms. Soil Sci. Soc. Am. J., 60: 821-828.
CrossRefDirect Link - Li, H.F., T. Imai, M. Ukita, M. Sekine and T. Higuchi, 2004. Compost stability assessment using a secondary metabolite: Geosmin. Environ. Technol., 25: 1305-1312.
CrossRefDirect Link - Muller, T., R. Thiβen, S. Braun, W. Dott and G. Fischer, 2004. (M)VoC and composting facilities part 1: (M)VOC emissions from municipal biowaste and plant refuse. Environ. Sci. Pollut. Res., 11: 91-97.
CrossRefDirect Link - Muller, T., R. Thiβen, S. Braun, W. Dott and G. Fischer, 2004. (M)VOC and composting facilities part 2: (M)VOC dispersal in the environment. Environ. Sci. Pollut. Res., 11: 152-157.
CrossRefDirect Link - Leff, J.W. and N. Fierer, 2008. Volatile Organic Compound (VOC) emissions from soil and litter samples. Soil Biol. Biochem., 40: 1629-1636.
CrossRefDirect Link - Gray, C.M., R.K. Monson and N. Fierer, 2010. Emissions of volatile organic compounds during the decomposition of plant litter. J. Geophys. Res.: Biogeosci., Vol. 115.
CrossRefDirect Link - Insam, H. and M.S.A. Seewald, 2010. Volatile Organic Compounds (VOCs) in soils. Biol. Fertil. Soils, 46: 199-213.
CrossRefDirect Link - Ramirez, K.S., C.L. Lauber and N. Fierer, 2010. Microbial consumption and production of volatile organic compounds at the soil-litter interface. Biogeochemistry, 99: 97-107.
CrossRefDirect Link - Wenke, K., M. Kai and B. Piechulla, 2010. Belowground volatiles facilitate interactions between plant roots and soil organisms. Planta, 231: 499-506.
CrossRefDirect Link - Perl, T., M. Junger, W. Vautz, J. Nolte, M. Kuhns, M. Borg-von Zepelin and M. Quintel, 2011. Detection of characteristic metabolites of Aspergillus fumigatus and Candida species using ion mobility spectrometry-metabolic profiling by volatile organic compounds. Mycoses, 54: e828-e837.
CrossRefDirect Link - Junger, M., W. Vautz, M. Kuhns, L. Hofmann and S. Ulbricht et al., 2012. Ion mobility spectrometry for microbial volatile organic compounds: A new identification tool for human pathogenic bacteria. Applied Microbiol. Biotechnol., 93: 2603-2614.
CrossRefDirect Link - Sundberg, C., D. Yu, I. Franke-Whittle, S. Kauppi and S. Smars et al., 2013. Effects of pH and microbial composition on odour in food waste composting. Waste Manage., 33: 204-211.
CrossRefDirect Link - Wu, H.S., W. Raza, J.Q. Fan, Y.G. Sun and W. Bao et al., 2008. Antibiotic effect of exogenously applied salicylic acid on in vitro soilborne pathogen, Fusarium oxysporum f. sp. niveum. Chemosphere, 74: 45-50.
CrossRefDirect Link - Tomas-Gallardo, L., H. Gomez-Alvarez, E. Santero and B. Floriano, 2014. Combination of degradation pathways for naphthalene utilization in Rhodococcus sp. strain TFB. Microb. Biotechnol., 7: 100-113.
CrossRefDirect Link - Meyer, J.M., P. Azelvandre and C. Georges, 1992. Iron metabolism in Pseudomonas: Salicylic acid, a siderophore of Pseudomonas fluorescens CHAO. BioFactors, 4: 23-27.
PubMedDirect Link - Koek, M.M., B. Muilwijk, M.J. van der Werf and T. Hankemeier, 2006. Microbial metabolomics with gas chromatography/mass spectrometry. Anal. Chem., 78: 1272-1281.
CrossRefDirect Link - Wunschel, D.S., K.F. Fox, A. Fox, M.L. Nagpal, K. Kim, G.C. Stewart and M. Shahgholi, 1997. Quantitative analysis of neutral and acidic sugars in whole bacterial cell hydrolysates using high-performance anion-exchange liquid chromatography-electrospray ionization tandem mass spectrometry. J. Chromatogr. A, 776: 205-219.
CrossRefDirect Link - Van Dam, J.C., M.R. Eman, J. Frank, H.C. Lange, G.W.K. van Dedem and S.J. Heijnen, 2002. Analysis of glycolytic intermediates in Saccharomyces cerevisiae using anion exchange chromatography and electrospray ionization with tandem mass spectrometric detection. Anal. Chim. Acta, 460: 209-218.
CrossRefDirect Link - Villas-Boas, S.G., S. Mas, M. Akesson, J. Smedsgaard and J. Nielsen, 2005. Mass spectrometry in metabolome analysis. Mass Spectrom. Rev., 24: 613-646.
CrossRefDirect Link