
Yanwirasti
Department of Anatomy, Faculty of Medicine, Andalas University, Padang, Indonesia
Wirsma A. Harahap
Division of Surgical Oncology, Medical School of M. Djamil Hospital, Andalas University, Padang, Indonesia
Dessy Arisanty
Department of Biochemistry, Faculty of Medicine, Andalas University, Padang, Indonesia
Pakistan Journal of Biological Sciences
Year: 2017 | Volume: 20 | Issue: 4 | Page No.: 189-196







ABSTRACT
Background: Abnormal expression of several microRNAs (miRNAs) has been demonstrated in many types of cancer tumor tissue. The miR-10b and miR-21 are an oncogenic miRNAs which play role in proliferation and invasion of Breast Cancer (BC) tumorigenesis. The aim of this study was to evaluate the miR-21 and miR-10b expression in BC in West Sumatran women, Indonesia. Materials and Methods: A total of 40 samples, consisting of 30 samples of breast cancer tissues (BC) and 10 samples of fibroadenoma tissues (FATs) as control and non-cancerous were analyzed. The miR-21 and miR-10b expression of each sample were investigated by using realtime PCR, followed by universal Reverse Transcription (RT) then real-time PCR amplification with specific primers. Hsa-miR-16-5p LNA PCR primer was used as an endogenous control. Results: The results showed that the expression level of miR-21 was more than 4 times higher in BC than in FATs. The expression level of miR-10b was lower in BC than FATs, by a factor of 3.34 fold. Both these differences were statistically different (p = 0.001). Conclusion: In this study it was concluded that for this sample of West Sumatran Women miR-21 expression in BC was higher than in FATs, whereas miR-10b was lower in BC than in FATs.
PDF Abstract XML References Citation
Received: November 07, 2016;
Accepted: January 23, 2017;
Published: March 15, 2017
Copyright: © 2017. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Yanwirasti, Wirsma A. Harahap and Dessy Arisanty, 2017. Evaluation of MiR-21 and MiR-10b Expression of Human Breast Cancer in West Sumatera. Pakistan Journal of Biological Sciences, 20: 189-196.
DOI: 10.3923/pjbs.2017.189.196
URL: https://scialert.net/abstract/?doi=pjbs.2017.189.196
DOI: 10.3923/pjbs.2017.189.196
URL: https://scialert.net/abstract/?doi=pjbs.2017.189.196
INTRODUCTION
It is estimated that almost 1.7 million cases of female Breast Cancer (BC) were diagnosed worldwide during 2012, corresponding to a rate of 43 per 100,000. Close to a quarter (24%) of all breast cancers were diagnosed within the Asia-Pacific region (approximately 404,000 cases at a rate of 30 per 100,000), with the greatest number of those occurring in China (46%), Japan (14%) and Indonesia (12%). It has been predicted that the international incidence of female breast cancer will reach approximately 3.2 million new cases per year by 20501. The mortality rate from diagnosed BC is increasing rapidly.
The dramatic increase in incidence rates in Asian populations renders it conceivable that in the near future the majority of breast cancer patients worldwide will be of Asian ethnicity2. Women in developing countries tend to have lower mortality rates from BC possibly because they know more about BC, are involved in public screening programs and seek medical help early which leads to early detection. Better understanding of the biological reason of BC and the use of biomarkers is essential for the early diagnosis and for better disease stratification and management.
Several genetic variants that predispose humans to BC have been identified; BRCA1/BRCA2 is the best-known hereditary gene for BC involved in 15% of all BC. Mutations of the TP53, CDH1, LKB1, PTEN, CHEK2, ATM, RAD51C, RAD51D and PALB2 genes are linked to 7% and known SNPs to 15%. However, while these genes contribute to over 36% of BC, 14% unknown SNPs and 50% is not related to any of these genetic abnormalities3. It is probably caused by the role of epigenetic which has not known.
Epigenetic factors also appear to play a role in BC incidence. These are heritable and modifiable markers that regulate gene expression without changing the underlying DNA sequence4. Epigenetic factors include DNA methylation, miRNA and histone deacetylation. Epigenetic mechanisms are recognized as significant factors in the development of BCs through the effects in genomic stability and regulated gene expression5.
In this study, the focus was on microRNAs (miRNAs) which are small 22-25 nucleotides non-coding RNAs, that have been conserved during evolution and control gene expression primarily at post-transcriptional and transcriptional levels6. The miRNAs have essential role in all biological processes, derive from genome encoded stem-loop precursors and function through RNA-induced silencing complex (RISC) mediated binding to targeted miRNAs by base pairing, mostly at the 3'-untranslated region (3'-UTR)7. Recently, the roles of miRNAs as important regulators of metabolism8, cancer-related immune pathways and to mediate the secretion of immunosuppressive or immunostimulating factor have been discovered9. Many miRNAs have been identified along with their roles in maintaining a variety of important processes in cell growth, such as differentiation, apoptosis, adhesion and other cellular processes8. The miRNAs functionally regulate gene expression by repressing mRNA translation and thus reducing target protein levels10.
Regulated miRNA expression is essential for maintaining cellular differentiation; therefore alterations in miRNA expression patterns are associated with several diseases, including various of cancers11. The miRNAs function as regulatory molecules as oncogenes or tumor suppressors. Amplification or overexpression of miRNAs down-regulate tumor suppressors or other genes involved in cell differentiation and genes expression12.
Abnormal expression of microRNAs has been observed in various types of cancer including breast cancer. Great efforts have been made to identify an association between microRNA expression profiles and breast cancer and to understand the functional role and molecular mechanism of aberrant-expressed microRNAs. As research has progressed, ‘oncogenic microRNAs’ and ‘tumor suppressive microRNAs’ have become a focus of interest13.
There are two types of miRNA; miRNA oncogenic (oncomiR) which inhibits the tumor suppressor gene and tumor suppressor miRNA (tumor suppressor miR) that inhibits the gene expression of oncogenes14. Both types may hold promise for miRNA gene therapies and are used as biomarkers. Each miRNA gene has different targets at each phase of carcinogenesis15. Alteration of miRNA levels is used for early detection of disease and intervention of the progress of the disease. However, miRNA levels and effectiveness in treatment may produce different results in different patients.
Iorio and Croce16 performed down-regulated processes with miR-10b, miR-125b and miR-145 and up-regulated miRNA-21 and miR-155 in BC using a miRNA microarray. Another study reported that miR-10b has altered expression in BC and hepatocellular cancer17. Interference in miRNA expression causes disruption of gene expression which in turn plays a role in the initiation, progression and metastasis of the developmental phase of BC. Micro RNA-10b plays a role in BC in a variety of cellular process such as proliferation, metastasis, angiogenesis and invasion of BC cells18,19. Increased proliferation occurs because of elevated expression of HOXD-10 (Homeobox Transcription Factor) which leads to the epithelial cells undergoing Epithelial Mesenchymal Transition (EMT) causing the progression of BC20.
Western models for risk of BC include factors such as age (over 50 years of age), reproductive status (nulliparous, early menarche and late menopause) family history, history of benign tumors, alcohol consumption, obesity and lack of physical activity have been identified21. In West Sumatera, different risk factors need to be considered. Most women with BC have low economic status, age 30-45 years of age, they are thin, multiparous, may be nursing a child and married before 25 years old and do not consume alcohol22. These facts raise the question whether the risk factor in of BC in West Sumatra, Indonesia are different from the West. Because of this different profile of BC incidence, the role of epigenetic factors could also be different. Oncogenic miR-21 and miR-10b clearly have important roles in the development of BC tissue. Understanding the way these epigenetic factors operate within West Sumatra population could be important to determine disease diagnosis and management.
In this study, it was tried to determine whether the expression of both miR-21 and miR-10b is different in (1) BC tissue and (2) FATs in the West Sumatra population. Fibroadenomas (FATs) are benign (non-cancer) tumors made up of both glandular breast tissue and stromal (connective) tissue that are most common in young women in 15-20 years of age. The authors are aware of no previous studies comparing miR-10b in BC tissues and FATs but the ready availability of tissue samples of FATs make them a practical control without the ethical issues related to obtaining healthy normal tissue in the absence of a tissue bank that provides healthy tissues. Characteristics of miR-10b and miR-21 in BC tissue serve as the basis for further studies to determine the best methods for treating BC that is most appropriate for West Sumatra women.
MATERIALS AND METHODS
Material: The material was BC tissues and fibroadenoma ((FATs), the most common benign mass in the female breast.
Main reagents: The main reagents were miRNA isolation Mini Kit (GenAids), cDNA synthesis kit II (Exiqon, Denmark), hsa-miR-16-5p LNA PCR primer set, hsa-miR-10b-5p LNA™ PCR primer set and hsa-miR-21-5p LNA PCR primer set (Exiqon, Denmark), ExiLENT SYBR Green master (Exiqon, Denmark).
Apparatus: The apparatus was micropipette (Eppendorf), qPCR system (CFX96 Realtime, Biorad USA), Nanodrop 1000 (Thermo, USA).
Preparation of samples: In the current study, miRNA-21 and miR-10b expression were evaluated in human BC tissue and FATs in West Sumatera with 30-50 years of age in range. Thirty samples of BC tissues and 10 FATs (non-cancerous) were used. Tissues were collected from BioBank of Medical Faculty Andalas University, Padang. All samples came from patients who had not undergone radical or modified mastectomies and had not received any chemotherapy or radiation before surgery.
miRNA extraction from BC tissue: Samples were stored at -80°C. Extraction was performed by using miRNA isolation Mini Kit (GenAids), the extraction was done based on the manufacturer's protocols. The miRNAs were purified on a glass-fiber filter and quantified by using the Nanodrop 1000 (Thermo, USA).
cDNA synthesis: The miRNAs (10-30 ng) were converted to cDNA by using miRCURY LNA Universal RT miRNA PCR, polyadenylation and cDNA synthesis kit II (Exiqon, Denmark) protocol. The miRNAs were incubated for 60 min at 42°C, the reverse transcriptase heat-inactivated for 5 min at 95°C, immediately cooled to 4°C and stored at 4°C or frozen to at least -20°C.
miRNA expression by real-time PCR: The expression of miRNA precursors was determined by using real-time quantitative PCR. The hsa-miR-16-5p LNA PCR primer set was used as an endogenous control. The miRNA experimental targets were hsa-miR-10b-5p LNA™ PCR primer set and hsa-miR-21-5p LNA PCR primer set (Exiqon, Denmark). The realtime mix was miRCURY LNA microRNA PCR, ExiLENT SYBR Green master (Exiqon, Denmark). The method is based on universal Reverse Transcription (RT) followed by real-time PCR amplification with LNA™ enhanced primers. The master mix contained 0.5 μL of 10X PCR buffer, 0.7 μL of 25 mM MgCl2, 0.1 μL of 12.5 mM dNTPs, 0.01 μL UNG, 0.5 μL of DNA Taq polymerase, SYBR Green, 0.5 μL of diluted cDNA (1:50) with sufficient water to make 3 μL. Three micro liters of the master mix containing all of the reaction components except the primers were dispensed into a 96-well real-time PCR plate (qPCR system, CFX96 Realtime, Biorad USA). Each primer solution (2 μM) was stored in 12-well PCR strip tubes and 2 μL was dispensed into duplicate wells of the 96-well plate. All reactions were performed in duplicate, including controls for each miRNA. The reactions were amplified for 15 sec at 95°C and 1 min at 60°C for 40 cycles. The thermal denaturation protocol (dissociation curve) was run at the end of the PCR to determine the number of products. Data were normalized to a universal endogenous control miRNA, which ideally was stably-expressed across the test sample set to correct for systematic variables such as amount of starting template, RNA quality and enzymatic efficiencies. The expression of each miRNA relative to miR-16 RNA was determined using 2‾ΔΔCT, ΔCT = (CtmiRNA-CtmiR-16).
The expression levels were normalized to the corresponding mean value of the reference miR-16. At first the expression levels of the analyzed microRNAs between breast cancer tissues and FATs tissues were compared. The relative expression ratio R which is presented as the n-fold change in miRNA expression.
Statistical analysis: A real-time PCR was used to determine the fluorescence intensity curve as it reaches the threshold cycle, crossing point (Cp), for each sample and a mean PCR efficiency corresponding to a primer pair in the exponential phase of each reaction. The expression of miRNA-21 and miRNA-10b of human BC and FATs was analyzed with t-test and correlation test by using the SPSS version 15.0 software programs, a p-value was 0.05 required to determine statistically.
qPCR experimental conditions: Amplification and data collection used CFX-96 Real-Time PCR (Biorad). Reactions were carried out in duplicate with 1 μM Eva Green (Biorad) of each primer. Condition profiles were 95°C for 5 min, 35 cycles; at 95°C for 30 sec; at 60°C for 10 sec and at 72°C for 30 sec.
RESULTS
To identify miRNAs associated with BC, the current study explored the expression of two human oncogenic miRNAs (miR-10b and miR-21) from 30 BC and 10 FATs samples with LNA-based on qPCR. Fibro adenoma was used as a control. The miR-16 was used as the control endogen for miR-21 and miR-10b. The melting curves of miR-21, miR-10b and miR-16 (as internal endogenous) were sharply defined with narrow peaks, indicating that pure and homogenous PCR products were produced. Highest peak mean lowest expression miR-21 has lower peaks (red) than miR-16 (endogenous control), it means that the expression of miR-21 is more higher (Fig. 1).
The result showed that miR-21 in BC tissue was higher than in FATs. The ΔCt value of miR-21 in BC tissue was lower than miR-21 in FATs, indicating that the expression of miR-21 in breast tumors tissue was higher than in the control. The median relative expression of miR-21 (2‾ΔΔCt) was 4.24 fold higher in BC tissue compared with FATs (Fig. 2).
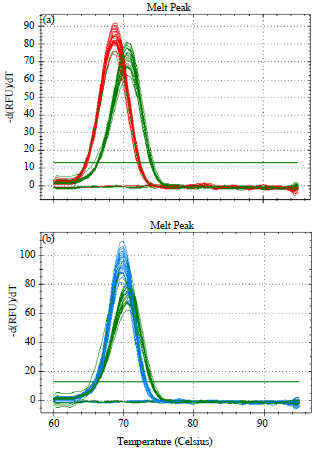 | |
| Fig. 1(a-b): | Melting curve of miRNA expression (a) Melting curve of miR-21 (red) relative to miR-16 (green) (b) Melting curve of miR-10b (blue) relative to miR-16 (green) |
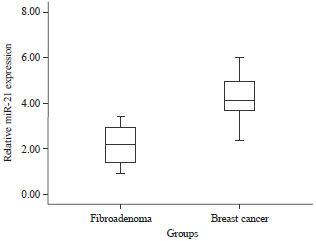 | |
| Fig. 2: | Difference in expression of miR-21 level between breast cancer tissue and fibroadenoma tissues (box-plot diagram with median) |
The level of expression of miR-21 between BC and FATs was statistically significantly different (p = 0.000) (Table 1).
The ΔCt was applied to mark the expression of miR-10b. If ΔCt was higher, this indicated that miR-10b expression quantity was lower. The expression of miR-10 and the Cp value of miR-10b in BC tissue were lower compared with FATs.
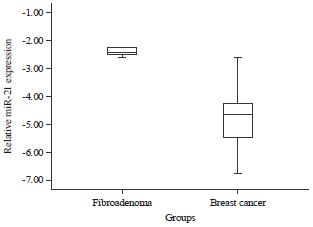 | |
| Fig. 3: | Difference in expression level of miR-10b between breast cancer tissue and fibroadenoma tissues which was used as control (box-plot diagram with median) |
| Table 1: | Difference in level of expression of miR-21 between BC and fibroadenoma |
| BC: Breast cancer tissue, FATs: Fibroadenoma tissues | |
| Table 2: | Difference in level of expression of miR-10b between BC and fibroadenoma |
| BC: Breast cancer tissue, FATs: Fibroadenoma tissues | |
The median relative expression of miR-10b (2‾ΔΔCt) was 3.34 fold lower in BC tissue compared to in FATs (Fig. 3).
The analysis of t-test statistically showed that the expression of miRNA-10b between BC tissue and FATs was significantly different (p = 0.001) (Table 2).
DISCUSSION
The miRNAs are endogenous non-encoding small RNAs, which regulate gene expression by binding with target mRNAs. The miRNAs bind to the 3' untranslated region (3' UTR) of mRNAs of the target and inhibit the translation of target proteins23. Each miRNA has multiple target genes and different miRNAs can regulate the same target gene. The miRNAs may act as an oncogene or tumor-suppressor, playing a role in tumorigenesis and development and regulating tumor proliferation, invasion, apoptosis and drug resistance13.
The miR-21 is a miRNA located in the FRA17B fragile area of chromosome 17q23.2 and has an independent transcription unit. The miR-21 expression is remarkably abnormal in many types of tumor cells, especially in breast cancer24. Abnormalities of miRNA expression in malignant cells compared with normal cells are often attributed to changes in genomic miRNA copy numbers and gene locations25. Alteration of copy numbers is indicated by decreased/increased miRNA expression. Aberrant miRNA expression may affect a multitude of transcripts.
The miR-21 is an oncogenic microRNA with increased expression in all human cancers, especially BC. Previous studies have found that miR-21 increased significantly in BC both in vivo and in vitro when detected by various methods26. In this study, it was found that expression miR-21 was higher in BC tissue than that in FATs. Huang et al.27 found that a high level of miR-21 is evident in breast cancer with highly uncontrolled cell proliferation and potential to be more aggressive. Hence it can be assumed, the fact that a higher level of miR-21 was found in these BC than in FATs also indicates the high level of uncontrolled proliferating cells which enable replicative immortality.
The miR-21 utilizes its oncogenic function mainly through cellular inhibition apoptosis28. Previous research also indicated that miR-21 is closely related to BCL-2. The BCL-2 is an anti-apoptosis protein which regulates caspase activity by helping to sequester cytochrome C in the mitochondria through inhibition of the mitochondrial permeabilizing protein Bax29. The miR-21 is involved in the apoptosis process. The miR-21 upregulation results in an increase of protein BCL-2. The increase of BCL-2 expression is induced by directly targeting miR-21, which is able to unconventionally upregulate the expression of its direct target by binding to its 3’ UTR30. Shi et al.31 discovered that miR-21 may downregulate Bax and upregulate BC-l2, inhibiting apoptosis. The outcome of these experiments confirmed that BCL-2 is a target for miR-21 and found that miR-21 is connected with intrinsic and/or extrinsic cellular apoptosis32. Thus, miR-21 has an important role in progression, invasion, dissemination and apoptosis of human breast cancer in vitro and in vivo.
It is well established that miR-21 regulates the expression of many anti-oncogenes, including TPM1, maspin and BCL-2 and programs cell death33. The overexpression of miR-21 in breast cancer suggests that miR-21 functions as an oncogene and its target genes are tumor suppressors. The miR-21 targets phosphatase angiotensin homolog (PTEN), one of the most commonly affected tumor suppressor genes. Moreover, miR-21 targets the (PTEN) gene by inhibiting gene expression in breast cancer cell34. In breast cancer cells, PTEN controls apoptosis and cell cycle arrest. Furthermore, the increase of miR-21 expression decreases the expression of PTEN and increases the activity of PI3K/Akt, as a result, NF-kB and Cyclin D1 increase. The PTEN is also involved in regulating cell migration and invasion by regulating matrix metalloproteinase 2 (MMP2) and matrix metalloproteinase 9 (MMP9)32. While the MMPs family genes are a key start the occurrence of the migration and invasion. These processes trigger uncontrolled proliferation. In addition, an increasing of miR-21 inhibits the metalloprotein tissue inhibitor-1 (TIMP1) gene accelerating the process of epithelial-mesenchymal transition (EMT) which is the main process in the EMT of cancer metastases35.
Another oncogenic miRNA that was researched in this study was mir-10b. Micro RNA-10b is located in chromosome 2q31. The miR-10b plays a role as an oncogene in many human malignancies, such BC. Studies have shown that miR-10b is related to cancer cells undergoing metastasis. The miR-10b is highly expressed in positive metastatic cancer cells and overexpressed in non-metastatic cancer cells which have begun a robust invasion and on the point of metastasis36.
In mammary epithelial cells and breast carcinoma cells, miR-10b directly suppresses the translation of HOXD10, an mRNA encoding a transcriptional repressor. The HOXD-10 represses RhoC, α3-integrin and MMP. All of three genes are involved in cell migration and extracellular matrix remodeling and stimulate the EMT process. This process is a key for regulation of cancer metastases36. The relation between miR-10b and metastasis remains controversial. Increased expression of miR-10b appears to be associated with breast cancer brain metastasis37. Thus, the over expression of miRNA-10b is closely related to the metastasis of breast cancer.
However, interestingly in this study, it was found that expression of miR-10b was lower in BC tissue than in FATs. This result was in contrast to previous studies that found miR-10b was over-expressed in hepatocellular carcinomas with positive metastasis38. Similarly, Ma et al.39 found that miR-10b was lower in non-metastatic breast tumors experiencing initial strong invasion and at the point of metastasis. Gee et al.40 also discovered that a lower level of expression of miR-10b occurred in early-stage of breast tumors. A similar result was found by Iorio and Croce16 who compared miR-10b expression in primary breast tumor tissues with non-cancer tissues. Those studies used normal tissues (NATs) as a comparison and the result showed that miR-10b expression was lower in breast tumors.
Furthermore, research meta-analysis by Lu et al.17 showed that expression of miR-10b varies and experiments show inconsistent results; sometimes miR-10b is up-regulated and sometimes down-regulated. These differences may be explained by type of cells, stage of a tumor or control used and source type of tissue. Low levels of miR-10b in BC tissue in this study could indicate that, although samples were selected without regard to the stage of cancer, most tissues used probably came from the primary breast tumor and suggests these cancers were highly proliferated but may not yet have metastasized and were still in an initial stage. The metastasis process is different from the initiation and progression of cancer in that not all of the transformed cells become metastatic41. The breast cancer tissue used was sourced from the compact solid tumor and not from cells that had escaped through the stromal cells in metastasis. So even though women in West Sumatra are often reluctant to seek early treatment for medical conditions and present with advanced cancer the tissue samples studied are likely to still only contain cancer cells that have not undergone metastasis. This seems unlikely as women in West Sumatera are often reluctant to seek early treatment for medical conditions. The result may be related to the control used. The miR-10b expression of tissue breast cancer was compared with FATs but not with Normal Adjuvant Tissue (NATs). Further exploration is required to determine the significance of the lower expression of miR-10b in this study.
The findings of this study are relevant since miR-10b could serve as a prognostic for early detection and/or a therapeutic target for anti-metastatic therapy. The measurement of miR-21 and miR-10b expression could also provide the rationale and selection criteria for treating early stage or pre-metastatic breast tumors with the miR-10b antagomiR or mimic miR as a part of a neoadjuvant regimen. The current study also suggested further analyzing the correlation between of miR-21 and miR-10b with stage of BC. However, further studies using a larger number of BC samples and using normal tissue as a control should be performed to confirm our findings and also to analyze the association between clinical parameters such as tumor subtypes, type of cell tumor, hormone status and clinical pathology profile.
CONCLUSION
This study showed miR-21 in BC in West Sumatran women was higher than in FATs, while miR-10b in BC was lower than in FATs. The results of this study can estimate that the tissue samples of subjects are generally still in early stages of tumorigenesis. So it can be used as a guide for oncologist later. It also be suggested further to analyze miRNA from blood or circulations, so it may likely be a non-invasive detection tool to investigate cancer development of patient whether in metastasis or has not metastasis. Finally it was concluded that miR-21 and miR-10b are potential biomarkers for early detection.
ACKNOWLEDGMENTS
This study was supported by a grant from the Research and Technology Ministry of Indonesian Government No. Sp DIPA-042.01.2.400928/2016. Special thanks to Dr. Daan Khambri, MD. for his contribution in sample preparation of BioBank tissue in Biomedical Laboratory.
REFERENCES
- Bhoo-Pathy, N., C.H. Yip, M. Hartman, C.S.P.M. Uiterwaal and B.C.R. Devi et al., 2013. Breast cancer research in Asia: Adopt or adapt Western knowledge? Eur. J. Cancer, 49: 703-709.
CrossRefDirect Link - Couch, F.J., K.L. Nathanson and K. Offit, 2014. Two decades after BRCA: Setting paradigms in personalized cancer care and prevention. Science, 343: 1466-1470.
CrossRefDirect Link - Terry, M.B., J.A. McDonald, H.C. Wu, S. Eng and R.M. Santella, 2016. Epigenetic Biomarkers of Breast Cancer Risk: Across the Breast Cancer Prevention Continuum. In: Novel Biomarkers in the Continuum of Breast Cancer, Stearns, V. (Ed.). Springer International Publishing, Switzerland, ISBN: 978-3-319-22908-9, pp: 33-68.
- Verma, M., S. Rogers, R.L. Divi, S.D. Schully and S. Nelson et al., 2014. Epigenetic research in cancer epidemiology: Trends, opportunities and challenges. Cancer Epidemiol. Prev. Biomarkers, 23: 223-233.
CrossRefDirect Link - Ahmad, J., S.E. Hasnain, M.A. Siddiqui, M. Ahamed, J. Musarrat and A.A. Al-Khedhairy, 2013. MicroRNA in carcinogenesis and cancer diagnostics: A new paradigm. Indian J. Med. Res., 137: 680-694.
PubMedDirect Link - Lin, S. and R.I. Gregory, 2015. MicroRNA biogenesis pathways in cancer. Nat. Rev. Cancer, 15: 321-333.
CrossRefDirect Link - Slack, F.J. and J.B. Weidhaas, 2008. MicroRNA in cancer prognosis. N. Engl. J. Med., 359: 2720-2722.
CrossRefDirect Link - Paladini, L., L. Fabris, G. Bottai, C. Raschioni, G.A. Calin and L. Santarpia, 2016. Targeting microRNAs as key modulators of tumor immune response. J. Exp. Clin. Cancer Res., Vol. 35.
CrossRefDirect Link - Meister, G., M. Landthaler, Y. Dorsett and T. Tuschl, 2004. Sequence-specific inhibition of microRNA- and siRNA-induced RNA silencing. RNA, 10: 544-550.
CrossRefDirect Link - Bajan, S. and G. Hutvagner, 2014. Regulation of miRNA processing and miRNA mediated gene repression in cancer. MicroRNA, 3: 10-17.
Direct Link - Shenouda, S.K. and S.K. Alahari, 2009. MicroRNA function in cancer: Oncogene or a tumor suppressor? Cancer Metastasis Rev., Vol. 28.
CrossRefDirect Link - Wang, W. and Y.P. Luo, 2015. MicroRNAs in breast cancer: Oncogene and tumor suppressors with clinical potential. J. Zhejiang Univ. Sci. B, 16: 18-31.
CrossRefDirect Link - Fu, S.W., L. Chen and Y.G. Man, 2011. miRNA biomarkers in breast cancer detection and management. J. Cancer, 2: 116-122.
CrossRefDirect Link - George, G.P. and R.D. Mittal, 2010. MicroRNAs: Potential biomarkers in cancer. Indian J. Clin. Biochem., 25: 4-14.
CrossRefDirect Link - Iorio, M.V. and C.M. Croce, 2012. Causes and consequences of microRNA dysregulation. Cancer J., 18: 215-222.
CrossRefDirect Link - Lu, Y., J. Yao, J. Yu, Q. Wei and X. Cao, 2014. The association between abnormal microRNA-10b expression and cancer risk: A meta-analysis. Scient. Rep., Vol. 4.
CrossRefDirect Link - Xiao, H., H. Li, G. Yu, W. Xiao and J. Hu et al., 2014. MicroRNA-10b promotes migration and invasion through KLF4 and HOXD10 in human bladder cancer. Oncol. Rep., 31: 1832-1838.
CrossRefPubMedDirect Link - Schickel, R., B. Boyerinas, S.M. Park and M.E. Peter, 2008. MicroRNAs: Key players in the immune system, differentiation, tumorigenesis and cell death. Oncogene, 27: 5959-5974.
CrossRefDirect Link - Zhang, Z.J. and S.L. Ma, 2012. miRNAs in breast cancer tumorigenesis (review). Oncol. Rep., 27: 903-910.
CrossRefDirect Link - Hunt, K.K., L.A. Newman, E.M. Copeland III and K.I. Bland, 2010. The Breast. In: Schwartz's Principles of Surgery, Brunicardi, F.C., D.K. Andersen, T.R. Billiar, D.L. Dunn, J.G. Hunter, J.B. Matthews and R.E. Pollock (Eds.). 9th Edn., Chapter 17, McGraw Hill, New York, USA., ISBN-13: 9780071547697.
Direct Link - Harahap, W.A., D. Arisanty, D. Khambri, Yanwirasti and S. Mubarika, 2015. [Methylated promoter gene BRCA1 and its influence on characteristics of premenopausal sporadic breast cancer among Minang ethnic patients]. Cermin Dunia Kedokteran, 42: 186-190, (In Indonesian).
Direct Link - Saito, T. and P. Sætrom, 2010. MicroRNAs-targeting and target prediction. New Biotechnol., 27: 243-249.
CrossRefDirect Link - Fujita, S., T. Ito, T. Mizutani, S. Minoguchi, N. Yamamichi, K. Sakurai and H. Iba, 2008. miR-21 gene expression triggered by AP-1 is sustained through a double-negative feedback mechanism. J. Mol. Biol., 378: 492-504.
CrossRefDirect Link - Lu, J., G. Getz, E.A. Miska, E. Alvarez-Saavedra and J. Lamb et al., 2005. MicroRNA expression profiles classify human cancers. Nature, 435: 834-838.
CrossRefPubMedDirect Link - Si, M.L., S. Zhu, H. Wu, Z. Lu, F. Wu and Y.Y. Mo, 2007. miR-21-mediated tumor growth. Oncogene, 26: 2799-2803.
CrossRefDirect Link - Huang, G.L., X.H. Zhang, G.L. Guo, K.T. Huang and K.Y. Yang et al., 2009. Clinical significance of miR-21 expression in breast cancer: SYBR-Green I-based real-time RT-PCR study of invasive ductal carcinoma. Oncol. Rep., 21: 673-679.
CrossRefPubMedDirect Link - Medina, P.P., M. Nolde and F.J. Slack, 2010. OncomiR addiction in an in vivo model of microRNA-21-induced pre-B-cell lymphoma. Nature, 467: 86-90.
CrossRefDirect Link - Wickramasinghe, N.S., T.T. Manavalan, S.M. Dougherty, K.A. Riggs, Y. Li and C.M. Klinge, 2009. Estradiol downregulates miR-21 expression and increases miR-21 target gene expression in MCF-7 breast cancer cells. Nucleic Acids Res., 37: 2584-2595.
CrossRefDirect Link - Dong, J., Y.P. Zhao, L. Zhou, T.P. Zhang and G. Chen, 2011. Bcl-2 upregulation induced by miR-21 via a direct interaction is associated with apoptosis and chemoresistance in MIA PaCa-2 pancreatic cancer cells. Arch. Med. Res., 42: 8-14.
CrossRefDirect Link - Shi, L., J. Chen, J. Yang, T. Pan, S. Zhang and Z. Wang, 2010. MiR-21 protected human glioblastoma U87MG cells from chemotherapeutic drug temozolomide induced apoptosis by decreasing Bax/Bcl-2 ratio and caspase-3 activity. Brain Res., 1352: 255-264.
CrossRefDirect Link - Buscaglia, L.E.B. and Y. Li, 2011. Apoptosis and the target genes of miR-21. Chin. J. Cancer, 30: 371-380.
CrossRefDirect Link - Ozgun, A., B. Karagoz, O. Bilgi, T. Tuncel, H. Baloglu and E.G. Kandemir, 2013. MicroRNA-21 as an indicator of aggressive phenotype in breast cancer. Oncol. Res. Treat., 36: 115-118 36: 115-118.
CrossRefDirect Link - Kuang, Y. and Y.J. Nie, 2016. Exploration of the regulatory effect of miR-21 on breast cancer cell line proliferation and invasion as well as the downstream target genes. Asian Pac. J. Trop. Med., 9: 470-473.
CrossRefDirect Link - Samatov, T.R., A.G. Tonevitsky and U. Schumacher, 2013. Epithelial-mesenchymal transition: Focus on metastatic cascade, alternative splicing, non-coding RNAs and modulating compounds. Mol. Cancer, Vol. 12.
CrossRefDirect Link - Ma, L., J. Teruya-Feldstein and R.A. Weinberg, 2007. Tumour invasion and metastasis initiated by microRNA-10b in breast cancer. Nature, 449: 682-688.
CrossRefDirect Link - Ahmad, A., S. Sethi, W. Chen, R. Ali-Fehmi, S. Mittal and F.H. Sarkar, 2014. Up-regulation of microRNA-10b is associated with the development of breast cancer brain metastasis. Am. J. Transl. Res., 6: 384-390.
Direct Link - Liao, C.G., L.M. Kong, P. Zhou, X.L. Yang, J.G. Huang, H.L. Zhang and N. Lu, 2014. miR-10b is overexpressed in hepatocellular carcinoma and promotes cell proliferation, migration and invasion through RhoC, uPAR and MMPs. J. Transl. Med., Vol. 12.
CrossRefDirect Link - Ma, L., F. Reinhardt, E. Pan, J. Soutschek and B. Bhat et al., 2010. Therapeutic silencing of miR-10b inhibits metastasis in a mouse mammary tumor model. Nature Biotechnol., 28: 341-347.
CrossRefDirect Link - Gee, H.E., C. Camps, F.M. Buff, S. Colella and H. Sheldon et al., 2008. MicroRNA-10b and breast cancer metastasis. Nature, 455: E8-E9.
CrossRefDirect Link - Heerboth, S., G. Housman, M. Leary, M. Longacre and S. Byler et al., 2015. EMT and tumor metastasis. Clin. Transl. Med., Vol. 4.
CrossRefDirect Link