
A.T. Djami-Tchatchou
Department of Agriculture and Animal Health, University of South Africa, Private Bag X6, 1710 Florida, South Africa
K. Ntushelo
Department of Agriculture and Animal Health, University of South Africa, Private Bag X6, 1710 Florida, South Africa
LiveDNA: 27.15205
Pakistan Journal of Biological Sciences
Year: 2017 | Volume: 20 | Issue: 3 | Page No.: 147-153







ABSTRACT
Background: Pectobacterium carotovorum subsp. carotovorum (Pcc) is a soft rot bacterium which upon entry into the plant macerates plant tissues by producing plant cell wall degrading enzymes. It has a wide host range which includes carrot, potato, tomato, leafy greens, squash and other cucurbits, onion, green peppers and cassava. During plant-microbe interactions, one of the ways of plant response to pathogen infection is through the small RNA silencing mechanism. Under pathogen attack the plant utilizes microRNAs to regulate gene expression by means of mediating gene silencing at transcriptional and post-transcriptional level. This study aims to assess for the first time, the expression profile of some stress-responsive miRNA and differential expression pattern of their target genes in Arabidopsis thaliana inoculated with Pcc. Materials and Methods: Leaves of five weeks old Arabidopsis thaliana plants were infected with Pcc and the quantitative real time-PCR, was used to investigate after 0, 24, 48 and 72 h post infection, the expression profiling of the stress-responsive miRNAs which include: miR156, miR159, miR169, miR393, miR396 miR398, miR399 and miR408 along with their target genes which include: Squamosa promoter-binding-like protein, myb domain protein 101, nuclear factor Y subunit A8, concanavalin A-like lectin protein kinase, growth regulating factor 4, copper superoxide dismutase, ubiquitin-protein ligase and plantacyanin respectively. Results: The findings showed that the overexpression of 6 miRNAs at 24, 48 and 72 h after infection resulted in the repression of their target genes and the expression of 2 miRNAs didn’t affect their target genes. Conclusion: These results provide the first indication of the miRNAs role in response to the infection of Pcc in A. thaliana and open new vistas for a better understanding of miRNA regulation of plant response to Pcc.
PDF Abstract XML References Citation
Received: December 23, 2016;
Accepted: January 24, 2017;
Published: February 15, 2017
Copyright: © 2017. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
A.T. Djami-Tchatchou and K. Ntushelo, 2017. Expression Profile of Stress-responsive Arabidopsis thaliana miRNAs and their Target Genes in Response to Inoculation with Pectobacterium carotovorum subsp. carotovorum. Pakistan Journal of Biological Sciences, 20: 147-153.
DOI: 10.3923/pjbs.2017.147.153
URL: https://scialert.net/abstract/?doi=pjbs.2017.147.153
DOI: 10.3923/pjbs.2017.147.153
URL: https://scialert.net/abstract/?doi=pjbs.2017.147.153
INTRODUCTION
Pectobacterium carotovorum subsp. carotovorum (Pcc) is a gram-negative enteric bacterium which, upon contact with the plant, produces enzymes which degrade the cell walls of plants to macerate plant tissue, causing the commonly known soft-rot symptoms1.
A transposon mutagenesis study on Pcc identified 14 genes as pathogenicity-related factors2 and this bacterium was also found to trigger defense responses and cell death in the model plant Arabidopsis thaliana3. During plant-microbe interactions, one of the ways of plant defense against pathogen infection is through the small RNA silencing mechanism4.
In order to further understand Pcc infection and extrapolate its pathogenicity-related factors with other infectious agents, the pathogenicity of Pcc was assessed in the model plant A. thaliana with a focus on the expression of A. thaliana stress-responsive microRNAs (miRNAs) and their target genes in response to inoculation with Pcc.
In this study, for the first time, we analysed expression profiles of stress-responsive microRNAs (miRNAs) and their target genes following the inoculation of A. thaliana with Pcc. The miRNAs are a class of functional, small non-coding RNAs ranging in length from 18 to 25 nucleotides that regulate gene expression either through post-transcriptional degradation or translational repression of their target mRNAs5. They are involved in the regulation of a multitude of biological processes in plants which include plant growth and development, maintenance of genome integrity, signal transduction, hormone signalling pathways, hormone homeostasis, plant immunity and responses to different environmental abiotic and biotic stresses4. The manipulation of stress-associated miRNAs and their target genes’ expression levels in plants is emerging as an effective strategy for improving the responses of plant crops to environmental stress and parasite attacks such as bacteria. In this regard, the aim of this study was to analyse, for the first time, the expression profiling of some stress-responsive miRNAs and their target genes in A. thaliana response to with Pcc infection.
MATERIALS AND METHODS
In order to assess the role of miRNAs in A. thaliana after infection with Pcc, the Pcc strain BD163 (obtained from the Agricultural Research Council-Plant Protection Research Institute, Tshwane, South Africa) was selected for this study. The Pcc was grown in lysogeny broth and about 10 mL of culture with approximately 2.5×106 CFU mL–1 from a 12 h old culture was used to inoculate (pressure infiltration) leaves of five weeks old A. thaliana plants6. The A. thaliana plants, derived from ecotype Columbia (Col-0), had been grown in a controlled greenhouse environment in a 10/14 h light-dark photoperiod. Control plants (mock sample) were also pressure-infiltrated with an equal amount of lysogeny broth. After infection, plants were kept at 25°C, 100% relative humidity under a 10/14 h light-dark. Leaf tissue was sampled after 0 h (mock sample, C0), 24, 48 and 72 h post infection and kept at -80°C for further analysis.
Based on their involvement in plant stress response as we found in our previous investigation7, the following stress-related miRNAs: miR156, miR159, miR169, miR393, miR396, miR398, miR399 and miR408, along with their target genes which include: Squamosa promoter-binding-like protein (At1g27360; SPL), Myb domain protein 101 ( At2g32460; MYB101), nuclear factor Y subunit A8 (At1g17590; NFY), concanavalin A-like lectin protein kinase (At3g59740; LEC), growth-regulating factor 4 (At3g52910; GR), copper superoxide dismutase (At1g08830; CDS), ubiquitin-protein ligase (At2g33770; UBC) and plantacyanin (At2g02850; PCYA) were selected. All primer pairs of the selected miRNAs and target genes were obtained from our previous study7.
To assess their response when A. thaliana is infected with Pcc, real-time PCR (qPCR) was used. Using TRIzol® Reagent, (Invitrogen, Carlsbad, CA, USA), total RNA was extracted from 300 mg of leaf tissues obtained from the mock sample (C0) and the samples obtained after 24, 48 and 72 h post infection followed by DNase treatment using DNase I (Thermo Scientific, Waltham, MA, USA). The RNA yield was measured with a ND-1000 spectrophotometer (Nano Drop) and the quality indexes A260/280 and A260/230 absorption ratios were verified. The integrity and size distribution of the extracted RNA molecules were checked electrophoretically on a 1.5% agarose gel in 1X TBE stained with 0.5 μg mL–1 ethidium bromide. For the target genes, the DNase-treated RNA was reverse transcribed to cDNA using a RevertAid™ Premium First Strand cDNA synthesis kit (Fermentas, Thermo Scientific, Waltham, MA, USA). For miRNAs, the DNase-treated RNA samples were polyadenylated and reverse transcribed using the Mir-X miRNA first-strand synthesis Kit (Takara, Clontech, Mountain View, CA, USA). Then the qPCR was performed on the Rotor gene-3000A machine (Qiagen, Venlo, Netherlands) using the Mir-X miRNA qRT-PCR SYBR Kits (Takara, Clontech, Mountain View, CA, USA) and the FastStart Essential DNA Green Master Kit (Roche, Mannheim, Germany) as described previously by Djami-Tchatchou and Dubery7 (Table 1). The expression analysis was performed on three biological repeats with three technical replicates of each.
| Table 1: | Nucleotide sequence of primers used for stress responsive miRNAs and target genes for qPCR from Djami-Tchatchou and Dubery7 |
 | |
| Table 2: | The qPCR of miRNAs expression analysis in A. thaliana leaf infected with Pcc |
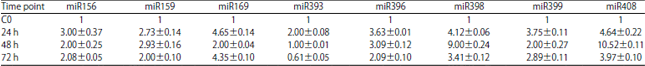 | |
Values are expressed as Mean±SEM of three biological repeats with three technical replicates of each (one-way ANOVA), The data was normalised to give relative gene expression using U6 small nuclear RNA. The expression ratio of the control sample (C0) was set as 1 at each time as it was used as the calibrator. Expression analysis was performed on three biological repeats with three technical replicates of each | |
The miRNA data was normalised to give relative gene expression using U6 small nuclear RNA and elongation factor 1-alpha and actin 8 were used to normalised the expression of the target genes. Data sets were statistically compared between non-treated samples and treated samples at each time point using one-way analysis of variation (ANOVA) with the statistical analysis software GraphPad in Stat 3 (GraphPad software, San Diego, CA, USA). The confidence level of all analyses was set at 95% and values with p<0.05 were considered significant.
RESULTS
At 24 h following Pcc infection the qPCR results (Fig. 1 a-h) showed that the expression profiles of all miRNAs were significantly up regulated with a maximum expression observed for miR156, miR159, miR169, miR393, miR396 and miR399 with the fold expression varying from relatively low (>2 fold) to high (>4 fold) compared to the basal levels of non-treated leaf. The expression profiles of SPL, CDS, UBC, PCYA were significantly down regulated with a minimum expression varying from relatively high (<1 fold) to Low (<0.1 fold) and the expression of GR, MYB101, NFY were up regulated.
At 48 h post bacterial infection the qPCR results showed that the expression profiles of all miRNAs except miR393 were significantly increased with the highest expression observed for miR159, miR396, miR398 and miR408 with the fold expression varying from relatively low (>3 fold) to high (>8 fold) compared to the basal levels of non-treated leafs (Fig. 1a-h).
At 72 h post infection with Pcc the qPCR results showed that the expression profiles of all miRNAs except miR393 were significantly increased with the highest expression observed for miR169 with the fold (>4fold) compared to the basal levels of non-treated leafs (Fig. 1a-h). A down regulation was observed for miR393 (Fig. 1d). At 48 and 72 h the expression profiles of MYB101 and GR were up regulated while the expression of SPL, NFY, LEC, CDS and UBC were significantly down regulated. All the statistical values of the gene expression of miRNA and their target genes are summarized in Table 2 and 3.
DISCUSSION
In this study, the miRNAs exhibited altered expression in A. thaliana in response to infection with Pcc. The miR156 known to be involved in plant defense response was found to be up-regulated at all time points (Fig. 1a), which resulted in the repression of its target SPL (Fig. 1a) a transcription factor of a family of plant-specific zinc finger protein that are involved in responses to abiotic and biotic stresses8.
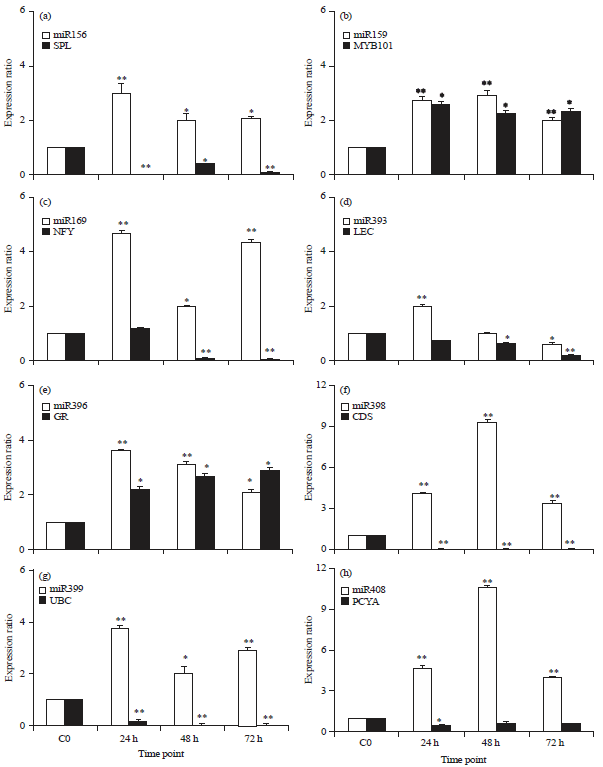 | |
| Fig. 1(a-h): | The qPCR of miRNAs and their target genes expression analysis in A. thaliana leaf inoculated with Pcc. The data was normalised to give relative gene expression using U6 small nuclear RNA for miRNA; elongation factor 1-alpha and actin 8 for target genes. The expression ratio of the control sample (C0) was set as 1 at each time as it was used as the calibrator. Expression analysis was performed on three biological repeats with three technical replicates of each wherein error bars represent the standard error of mean. *There was a significant difference with p<0.05, **There was a highly significant difference with p<0.01. Squamosa promoter-binding-like protein (At1g27360; SPL), Myb domain protein 101 (At2g32460; MYB101), nuclear factor Y subunit A8 (At1g17590; NFY), concanavalin A-like lectin protein kinase (At3g59740; LEC), growth-regulating factor 4 (At3g52910; GR), copper superoxide dismutase (At1g08830; CDS), ubiquitin-protein ligase (At2g33770; UBC) and plantacyanin (At2g02850; PCYA) |
| Table 3: | The qPCR of miRNA target genes expression analysis in A. thaliana leaf infected with Pcc |
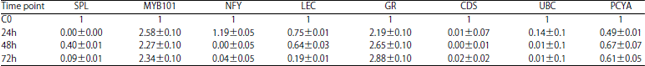 | |
Values are expressed as Mean±SEM of three biological repeats with three technical replicates of each (one-way ANOVA), SPL: Squamosa promoter-binding-like protein (SPL), MYB101: Myb domain protein 101, NFY: Nuclear factor Y subunit A8, LEC: Concanavalin A-like lectin protein kinase, GR: growth-regulating factor 4, CDS: Copper superoxide dismutase, UBC: Ubiquitin-protein ligase, PCYA: Plantacyanin, The data was normalised to give relative gene expression using elongation factor 1-alpha and actin 8. The expression ratio of the control sample (C0) was set as 1 at each time as it was used as the calibrator. Expression analysis was performed on three biological repeats with three technical replicates of each | |
This suggests that miR156 is involved in the response of A. thaliana to Pcc infection.
The miR159 was reported to be a bacteria-responsive miRNA induced in tomato plants during response to the infection by the bacterial pathogen Pseudomonas syringae pv. tomato9. We found that it was induced following inoculation of A. thaliana with Pcc (Fig. 1b). In contrast, the expression of its target a transcription factor Myb 101 showed a significant up-regulation at all time points (Fig. 1b), indicating that miR159 did not mediate their cleavage but. As it is known that the transcriptional termination of negative regulators7 and the transcriptional activation of positive regulators10 of gene expression are important steps for plant defense response; our result could imply that the induction of the transcription factor Myb101 indicated their involvement in the regulation of the response of A. thaliana to Pcc infection.
The expression patterns of miR169 showed significant high expression at all time points following the infection with the bacterium (Fig. 1c). Then the miR169 repressed the expression of its target gene NFY (Fig. 1c) a group of transcription factors that have three distinct subunits (NF-YA, NF-YB and NF-YC) that bind to the CCAAT box11. In a previous study the miR169 was shown to be induced in response to the bacterium Erwinia amylovora in apples12. In this regard, our finding revealed that miR169 is a bacteria-responsive miRNA to Pcc infection.
The miR393 investigated in this study was also reported to be a bacteria-responsive miRNA involved in the resistance to infection by Pseudomonas syringae of A. thaliana13. It targets LEC an inducible receptor for recognition of extracellular stimuli involved in plant stress response14. It was found that the increased expression of miR393 after 24 h (Fig. 1d) negatively affects the expression of its target LEC which showed a slight down-regulation (Fig. 1d). Therefore the result suggests that miR393 contributes to the response of A. thaliana to the infection of Pcc.
The expression level of miR396 was induced at all time points following the infection with the highest level at 24 h with a 3.63 fold change (Fig. 1e). It was shown that miR396 was involved in the regulation of defense in response to a citrus greening disease caused by Candidatus Liberibacter15. It targets GR which was reported to play a role in coordinating the interaction between developmental processes and defense dynamics16. It was found that the expression of its target GR was also induced and not controlled by miR396 at all time points (Fig. 1e), meaning that miR396 and GR are both involved in high expression during A. thaliana response to Pcc infection.
In A. thaliana, the miR398 was shown to be linked directly to the plant stress regulatory network and was confirmed to play a crucial role in responses to environmental and parasitic stresses17. We found that after Pcc infection, its expression significantly increased (Fig. 1f) and resulted in the significant down-regulation of its target CDS (Fig. 1f). The CDS act as antioxidant and protecting cellular components from being oxidized by Reactive Oxygen Species (ROS)18. Since pathogen invasion, can cause an enhanced production of ROS which result in the activation of the signal transduction our finding suggests that the miR398 repression of CDS prevented them to limit the formation of ROS and their removal from the plant cell in order to enhance the response of A. thaliana to the infection of Pcc. It was also found that miR399 was significantly overexpressed at all time points with a higher down-regulation of its target UBC (Fig. 1g). The UBC is involved in the degradation of proteins which act as negative regulators of defense during plant-pathogen interactions through ubiquitination process19. Accordingly we hypothesised that during the response of A. thaliana to Pcc inoculation, the ubiquitination process was not activated due to the absence of negative regulators of defense which resulted in the repression of UBC by overexpression of miR399. It was reported that the overexpression of miR399 played a crucial role in tomato viral defense response after a tomato was infected with the leaf curl virus20. Based on this observation, the present results suggest that miR399 is also involved in the response of A. thaliana to Pcc infection.
Finally, the qPCR highlighted that the expression of the stress-responsive miR408 was significantly up-regulated with the repression of their target PCYA at all time points following the infection of Arabidopsis with Pcc (Fig. 1h). The PCYA are blue copper proteins which are known to be involved in cell-to-cell signaling, redox reactions and stress responses that occur during primary defense responses21. Previous studies reported that miR408 is involved in plant stress response7. Therefore the present results suggest that the repression of the PCYA by the overexpression of miR408 might be required to boost the response of A. thaliana to infection with Pcc.
CONCLUSION
In this study for the first time, the expression profiling of some stress-responsive miRNAs and their target genes involved in the response of A. thaliana to inoculation with was analysed. The results initiate new insight to understand the function of the miRNA regulation of defense response against the bacterium Pcc and therefore open a way for future studies to gain a global perspective of the expression profiles of more miRNAs with their regulated target genes expressed in response to Pcc infection.
SIGNIFICANCE STATEMENT
To the best of our knowledge, this is the first attempt to explore Arabidopsis thaliana stress associated miRNAs regulation of the response to Pectobacterium carotovorum infection. The results thus provide valuable information and a new insight to understand the function of the miRNAs regulation of defense response against the bacterium Pcc attack and therefore may significantly contribute to the design strategy for crops prevention and protection against soft rot disease. However, for a better understanding of this miRNA regulation, present finding open a way for future strategy such as the high throughput deep sequencing technology to gain a global perspective of the expression profiles of miRNAs with their regulated target genes expressed in response to Pcc infection.
ACKNOWLEDGMENTS
The University of South Africa for the fellowship support to Djami-Tchatchou and support to Khayalethu Ntushelo. We thank Prof Ian Dubery from the University of Johannesburg for providing instruments. Prof Khajamohiddin Syed from the Central University of Technology, Bloemfontein, provided valuable suggestions for improving this article.
REFERENCES
- Barras, F., F. Van Gijsegem and A.K. Chatterjee, 1994. Extracellular enzymes and pathogenesis of soft-rot Erwinia. Ann. Rev. Phytopathol., 32: 201-234.
CrossRefDirect Link - Lee, D.H., J.A. Lim, J. Lee, E. Roh and K. Jung et al., 2013. Characterization of genes required for the pathogenicity of Pectobacterium carotovorum subsp. carotovorum Pcc21 in Chinese cabbage. Microbiology, 159: 1487-1496.
CrossRefDirect Link - Kariola, T., T.A. Palomaki, G. Brader and E.T. Palva, 2003. Erwinia carotovora subsp. carotovora and Erwinia-derived elicitors HrpN and PehA trigger distinct but interacting defense responses and cell death in Arabidopsis. Mol. Plant-Microbe Interact., 16: 179-187.
CrossRefDirect Link - Sun, G., 2012. MicroRNAs and their diverse functions in plants. Plant Mol. Biol., 80: 17-36.
CrossRefDirect Link - Sunkar, R. and J.K. Zhu, 2004. Novel and stress-regulated micrornas and other small RNAs from Arabidopsis. Plant Cell., 16: 2001-2019.
CrossRefPubMedDirect Link - Norman-Setterblad, C., S. Vidal and E.T. Palva, 2000. Interacting signal pathways control defense gene expression in Arabidopsis in response to cell wall-degrading enzymes from Erwinia carotovora. Mol. Plant-Microbe Interact., 13: 430-438.
CrossRefDirect Link - Djami-Tchatchou, A.T. and I.A. Dubery, 2015. Lipopolysaccharide perception leads to dynamic alterations in the microtranscriptome of Arabidopsis thaliana cells and leaf tissues. BMC Plant Biol., Vol. 15.
CrossRefDirect Link - Usami, T., G. Horiguchi, S. Yano and H. Tsukaya, 2009. The more and smaller cells mutants of Arabidopsis thaliana identify novel roles for squamosa promoter binding protein-like genes in the control of heteroblasty. Development, 136: 955-964.
CrossRefDirect Link - Zhang, W., S. Gao, X. Zhou, P. Chellappan and Z. Chen et al., 2011. Bacteria-responsive microRNAs regulate plant innate immunity by modulating plant hormone networks. Plant Mol. Biol., 75: 93-105.
CrossRefDirect Link - Djami-Tchatchou, A.T., M.P. Maake, L. Piater and I.A. Dubery, 2015. Isonitrosoacetophenone drives transcriptional reprogramming in Nicotiana tabacum cells in support of innate immunity and defense. PLoS ONE, Vol. 10.
CrossRefDirect Link - Gusmaroli, G., C.Tonelli and R. Mantovani, 2002. Regulation of novel members of the Arabidopsis thaliana CCAAT-binding nuclear factor Y subunits. Gene, 283: 41-48.
CrossRefDirect Link - Kaja, E., M.W. Szczesniak, P.J. Jensen, M.J. Axtell, T. McNellis and I. Makalowska, 2015. Identification of apple miRNAs and their potential role in fire blight resistance. Tree Genet. Genomes, Vol. 11.
CrossRefDirect Link - Navarro, L., P. Dunoyer, F. Jay, B. Arnold and J.D.G. Jones et al., 2006. A plant miRNA contributes to antibacterial resistance by repressing auxin signaling. Science, 312: 436-439.
CrossRefDirect Link - Sanabria, N.M., H. van Heerden and I.A. Dubery, 2012. Molecular characterisation and regulation of a Nicotiana tabacum S-domain receptor-like kinase gene induced during an early rapid response to lipopolysaccharides. Gene, 501: 39-48.
CrossRefDirect Link - Rawat, N., S.P. Kiran, D. Du, F.G. Gmitter and Z. Deng, 2015. Comprehensive meta-analysis, co-expression, and miRNA nested network analysis identifies gene candidates in citrus against Huanglongbing disease. BMC Plant Biol., Vol. 15.
CrossRefDirect Link - Liu, J., J.H. Rice, N. Chen, T.J. Baum and T. Hewezi, 2014. Synchronization of developmental processes and defense signaling by growth regulating transcription factors. PLoS ONE, Vol. 9.
CrossRefDirect Link - Bouche, N., 2010. New insights into miR398 functions in Arabidopsis. Plant Signal. Behav., 5: 684-686.
CrossRefDirect Link - Alscher, R.G., N. Erturk and L.S. Heath, 2002. Role of superoxide dismutases (SODs) in controlling oxidative stress in plants. Exp. Bot., 53: 1331-1341.
CrossRefPubMedDirect Link - Zeng, L.R., M.E. Vega-Sanchez, T. Zhu and G.L. Wang, 2006. Ubiquitination-mediated protein degradation and modification: An emerging theme in plant-microbe interactions. Cell Res., 16: 413-426.
CrossRefDirect Link - Naqvi, A.R., Q.M.R. Haq and S.K. Mukherjee, 2010. MicroRNA profiling of tomato leaf curl New Delhi virus (ToLCNDV) infected tomato leaves indicates that deregulation of mir159/319 and mir172 might be linked with leaf curl disease. Virol. J., Vol. 7.
CrossRefDirect Link - Romo, S., E. Labrador and B. Dopico, 2001. Water stress-regulated gene expression in Cicer arietinum seedlings and plants. Plant Physiol. Biochem., 39: 1017-1026.
CrossRefDirect Link