
Abdalla Dao
Institute of Environment and Agricultural Research (INERA), BP 910, Bobo, Dioulasso, Burkina Faso
Jacob Sanou
Institute of Environment and Agricultural Research (INERA), BP 910, Bobo, Dioulasso, Burkina Faso
Edgar V.S. Traore
Institute of Environment and Agricultural Research (INERA), BP 910, Bobo, Dioulasso, Burkina Faso
Vernon Gracen
Department of Plant Breeding and Genetics, 520 Bradfield Hall, Cornell University, NY 14850, Ithaca, USA
Eric Y. Danquah
West Africa Centre for Crop Improvement (WACCI), University of Ghana, BMP 30, Legon, Accra, Ghana
Pakistan Journal of Biological Sciences
Year: 2017 | Volume: 20 | Issue: 3 | Page No.: 132-139







ABSTRACT
Background: In drought-prone environments, direct selection for yield is not adequate because of the variable environment and genotype x environment interaction. Therefore, the use of secondary traits in addition to yield has been suggested. The relative usefulness of secondary traits as indirect selection criteria for maize grain yield is determined by the magnitudes of their genetic variance, heritability and genetic correlation with the grain yield. Materials and Methods: Forty eight testcross hybrids derived from lines with different genetic background and geographical origins plus 7 checks were evaluated in both well-watered and water-stressed conditions over two years for grain yield and secondary traits to determine the most appropriate secondary traits and select drought tolerant hybrids. Results: Study found that broad-sense heritability of grain yield and Ear Per Plant (EPP) increased under drought stress. Ear aspect (EASP) and ear height (EHT) had larger correlation coefficients and direct effect on grain yield but in opposite direction, negative and positive respectively. Traits like, EPP, Tassel Size (TS) and Plant Recovery (PR) contributed to increase yield via EASP by a large negative indirect effect. Under drought stress, EHT had positive and high direct effect and negative indirect effect via plant height on grain yield indicating that the ratio between ear and plant heights (R-EPH) was associated to grain yield. Conclusion: Path coefficient analysis showed that traits EPP, TS, PR, EASP, R-EPH were important secondary traits in the present experiment. These traits were used in a selection index to classify hybrids according to their performance under drought. The selection procedure included also a Relative Decrease in Yield (RDY) index. Some secondary traits reported as significant selection criteria for selection under drought stress were not finally established in the present study. This is because the relationship between grain and secondary traits can be affected by various factors including germplasm, environment and applied statistical analysis. Therefore, different traits and selection procedure should be applied in the selection process of drought tolerant genotypes for diverse genetic materials and growing conditions.
PDF Abstract XML References Citation
Received: November 10, 2016;
Accepted: January 16, 2017;
Published: February 15, 2017
Copyright: © 2017. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Abdalla Dao, Jacob Sanou, Edgar V.S. Traore, Vernon Gracen and Eric Y. Danquah, 2017. Selection of Drought Tolerant Maize Hybrids Using Path Coefficient Analysis and Selection Index. Pakistan Journal of Biological Sciences, 20: 132-139.
DOI: 10.3923/pjbs.2017.132.139
URL: https://scialert.net/abstract/?doi=pjbs.2017.132.139
DOI: 10.3923/pjbs.2017.132.139
URL: https://scialert.net/abstract/?doi=pjbs.2017.132.139
INTRODUCTION
Maize is an important staple food in Sub-Saharan Africa, occupying over 50% of land devoted to cereal production in over 50% of countries in this region1. However, maize productivity in this part of the world is limited by drought. There is need therefore to improve yield of maize under arid and semi-arid environments through breeding. Drought tolerance is a complex trait which is controlled by polygenes and their expressions are influenced by various environmental factors2,3. Breeding for drought tolerance is always an important challenge for plant breeders. In maize, yield is the primary trait but selection for improved genotypes under drought based on grain yield alone was revealed inefficient4 due to low heritability of grain yield and the complexity of genotype-environment interactions. Genetic variance and heritability of maize grain yield are reduced under drought whereas secondary traits have relatively high genetic variance and heritability5,6. Secondary traits correlated to grain yield are therefore used with grain yield to select drought tolerant maize genotypes7,8. However, the relative usefulness of secondary traits as indirect selection criteria for grain yield is often not conclusively established in all the experiments because of the nature of the genetic material and conditions of experiments. Thus, there is need to ascertain the value of a secondary trait before to include in a selection index for each experiment4. In addition, the correlation coefficient between grain yield and secondary traits is explained by the direct contribution of the trait on grain yield as well as the indirect contribution through other traits which should also be considered during selection9,10.
The objective of this study was to determine the most appropriate secondary traits to be included in the selection index to select maize drought tolerant hybrids. Thus, broad-sense heritability of the traits and phenotypic correlations were computed. The correlation was further portioned into direct and indirect effects using path coefficient analysis.
MATERIEL AND METHODS
Plant materials: Forty eight maize testcross hybrids plus 7 checks were used in this study. The hybrids were generated using line by tester mating design from twenty four white inbred lines with different genetic background and geographical origin.
Field evaluation and experimental design: The trials were conducted during the off-season in two years at the experimental station of INERA at Valley Du Kou located at 11°22’ N latitude, 4°22’ W longitude and at 300 m above sea level, characterized by ferruginous and acid soils with silty texture. The 48 white hybrids plus 7 checks were randomized in incomplete block, 11×5 alpha lattice design with three replications. The experimental unit was a single row of 5 m spaced 80 cm. Two seeds were planted per hill spaced 25 cm and thinned to one plant per hill to give a final population density of 50,000 plants ha–1. The trials were established and managed according to procedures developed by CIMMYT4. Adequate irrigation was applied in both water regimes from planting and throughout the vegetative phase, using furrow irrigation system. Drought stress was achieved in water stressed regime by withholding water two weeks before the expected flowering time for 21 days.
Field measurements: Grain yield and 12 secondary traits were measured or calculated. In each plot, days to anthesis and days to silking were recorded as the number of days from planting to when 50% of the plants had shed pollen and emerged silks, respectively. Anthesis Silking Interval (ASI) was computed as the interval in days between dates of silking and anthesis. Plant and ear heights were measured in centimeter as the distance from the base of the plant to the height of the first tassel branch and the node bearing the upper ear, respectively. Ear aspect was scored on a 1-5 scale, where 1 = Clean, uniform, large and well-filled ears and 5 = Rotten, variable, small and partially filled ears. The total number of plants and ears were counted in each plot at the time of harvest. The number of ears per plant was then calculated as the proportion of the total number of ears at harvest divided by the total number of plants at harvest. Additional data obtained from drought stress plots were the leaf senescence recoded two times at weekly interval commencing two weeks after stress application on a scale of 1-10, where 1= Almost all leaves were green and 10 = Virtually all leaves were dead; Leaf rolling recorded on a scale of 1-5, where 1 = All leaves are unrolled and 5 = All leaves are rolled, leaf rolling was recorded three time at weekly interval commencing a week after stress application; leaf erectness was recorded based on visual score of 1 (erect leaves) to 5 (lax leaves) two times at weekly interval commencing a week after stress application; tassel size was recorded two times at weekly interval commencing 21 days after stress application based on scale of 1(small tassel size) to 5 (large tassel size); plant recovery was recorded at 7 and 14 days after stopping drought stress on a scale of 1-5, where 1 = All plants recovered from drought stress and 5 = All plants were dead.
Grain yield was calculated as follow: In well-watered evaluation, all ears harvested from each plot were weighed and representative samples of ears were shelled to determine per cent moisture. Grain yield adjusted to 15% moisture was, thus, computed from ear weight and grain moisture assuming a shelling percentage of 80%, based on the following formula:
Grain yield (kg ha–1) = Ear weight (kg)× 0.8×(100-moisture)/85)×(10/area m2 )×1000
In water stressed evaluation, all ears harvested from each plot were shelled to determine per cent moisture. Grain yield adjusted to 15% moisture was computed from the shelled grain based on the following formula:
Grain yield (kg ha–1) = Grain weight (kg)×(100-moisture)/85)×(10/area m2)×1000 Grain yield
Statistical analysis
Analysis of variance: Individual analysis of variance of each measured trait in each year and water regime were conducted with the PROC MIXED procedure from SAS with genotypes considered as fixed effects and replication and blocks within replication as random effects. Because the alpha lattice did not provide significant efficiency over Randomized Complete Block Design (RCBD), data were analyzed as from RCBD. Combined analysis of variance were conducted with PROC GLM in SAS11 using RANDOM statement with Test option. For combined analysis in Well Watered (WW) and Water Stressed (WS) conditions, mean square for genotypes, years and genotypes x years (G x Y) were tested against the mean squares for G x Y, replications within years and the pooled error, respectively.
Heritability estimates: Variance components were estimated from plot raw data using VARCOMP in SAS with years, replications, genotypes and the genotype x year interactions as random factors.
Broad-sense heritability of traits (![]() ) was also calculated on an plot basis as the ratio between the genetic variance (
) was also calculated on an plot basis as the ratio between the genetic variance (![]() ) and the sum of the genetic variance, the variance of the genotype x year interaction (
) and the sum of the genetic variance, the variance of the genotype x year interaction (![]() ) divided by the number of years (y) and the variance of the residuals (
) divided by the number of years (y) and the variance of the residuals (![]() ) divided by the number of replications (r) and years:
) divided by the number of replications (r) and years:
![]()
Path coefficient analysis: Path coefficient analysis were performed to evaluate the relative importance of direct and indirect effects of secondary traits on yield. Pearson correlation (phenotypic correlation) was first calculated and then the correlation coefficients were partitioned into direct and indirect effects through path coefficient analysis using PATHSAS program developed by Cramer et al.12.
Selection tolerance index and estimation of relative decrease in yield: Selection Tolerance Index (STI) was proposed by Hao et al.8. The algorithm is as follows for STI:
| (1) |
| (2) |
| (3) |
In the above three equations, j represents secondary trait. Sij in the Eq. 1, indicates the standardized phenotypic value of trait j to genotype i under the WW regime (w) and under the WS regime (d), χij indicates the real phenotypic value of genotype i on trait j, μj indicates the mean value of trait j for all materials and σj indicates the standard deviation of trait j. The MPij in Eq. 2, indicates the mean productivity of trait j on genotype i; bj in Eq. 3, indicates the given weight value of trait j.
Relative Decrease in Yield (RDY) was defined as:
RDY = 100-(Ys/Yp×100)13
where, Yp and Ys are yield potential under well-watered and drought stress respectively.
RESULTS AND DISCUSSION
Phenotypic variation: Table 1 shows the average phenotypic data per water regime and the mean squares of genotype and genotype by year interactions of the 13 measured traits. Results showed significant variability for many traits. Significant genotype x year interaction was also observed for different traits including Grain Yield (GY) and Anthesis Silking Interval (ASI) in both water regimes. Drought stress significantly reduced the GY (34%) and increased the ASI (37%).
Path coefficient analysis and identification of suitable secondary traits
Grain Yield (GY): Maize grain yield is affected by abiotic stresses such as drought and low N. Genetic variance and heritability for GY decline under stress14,15.
| Table 1: | Phenotypic variation for maize hybrids under well watered and water stress conditions across two years |
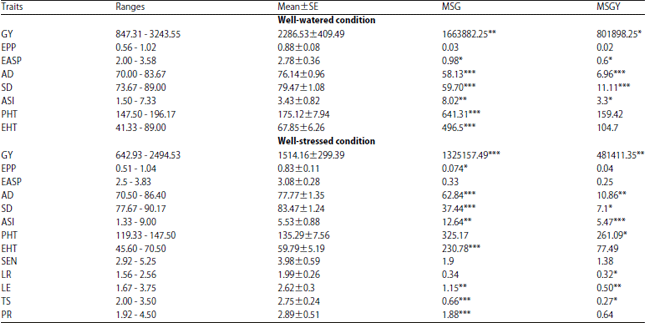 | |
SE: Standard error, *’ **’***Significant at p = 0.05, 0.01 and 0.001 respectively, GY: Grain yield, EPP: Ear per plant, EASP: Ear aspect, AD: Anthesis days, ASI: Anthesis silking interval, PHT: Plant height, EHT: Ear height, SEN: Leaf senescene, LR: Leaf rolling, LE: Leaf erectness, TS: Tassel size, PR: Plant recovery | |
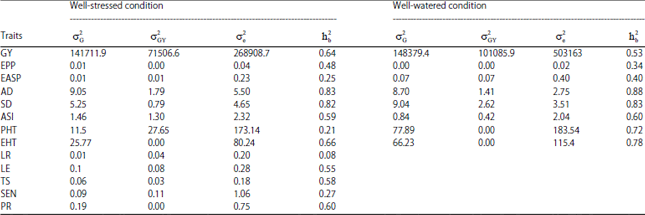
In the present study, the genetic variance for grain yield was lower under drought conditions compared to well-watered conditions (Table 2). Reduced genetic variance for grain yield under moisture stress conditions was also reported by Messmer et al.16 and Almeida et al.6. However the heritability estimates for grain yield of 0.53 under well-watered conditions was less than the value of 0.64 estimated under moisture stressed (WS) conditions, indicating high yield stability of hybrids. According to Almeida et al.6, high heritability estimates of grain yield under WS environments implies stability of drought tolerant genotypes across diverse environments.
Ear Per Plant (EPP) and grain yield: The phenotypic correlation coefficient between EPP and grain yield was significant and positive (r = 0.63***) and the direct effect of EPP on grain yield was positive and high (0.27). The heritability value (0.48) of EPP for hybrids was similar to the 0.54 reported by Bolanos and Edmeades5. The high positive direct effect of EPP on grain yield is explained by the high indirect contribution of EASP (0.23) and EHT (0.17) on grain yield.
Ear aspect (EASP) and grain yield: Among the traits, ear aspect had the highest total direct effect (-0.48) on grain yield. This result is in agreement with Badu-Apraku et al.17 who identified ear aspect with highest direct effect on grain yield in the evaluation of extra early maize inbred lines under drought stress. The phenotypic correlation between the traits in the present study was also negative and significant (r = - 0.70***). Therefore, the correlation represented the true relationship between the two traits. Ear aspect scores captured the different parameter of yield components such as ear diameter, ear length, kernels rows and kernel per row. High positive direct effect of ear diameter, ear length and number of kernels per row on maize grain yield was reported18. Ahmad and Saleem19 showed that the direct effect of kernel rows on grain yield per plant was also positive and greater in magnitude than that of genotypic correlation. Heritability of EASP in the present study was low (0.25) under drought. Because of the small variance, selection for this trait may not be effective even though the correlation of EASP with grain yield under drought was high.
Anthesis DAYS (AD) and grain yield: Direct effect of AD on grain yield was negative (-0.28) but the positive and high indirect effect through EHT and PR may have caused the positive correlation observed under drought stress (0.16) (Table 3). Heritability of AD was high (Table 2) which is consistent with earlier reports5,20. Grain yield potential of hybrids tested was less affected by their maturity since the correlation between AD and grain yield was low and non significant. However a negative significant phenotypic and genotypic correlation were found elsewhere17,21.
| Table 2: | Estimates of genetic variance components and broad sense heritabilities of grain yield and other traits of hybrids evaluated under two water regimes over 2 years |
 | |
| Table 3: | Partition of correlation coefficients into direct and indirect effects on grain yield under drought stress |
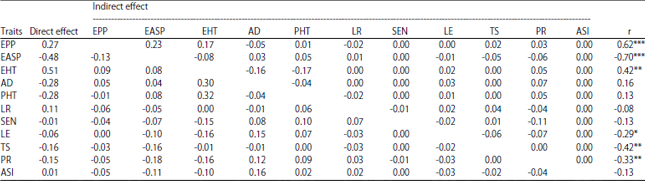 | |
R2: 0.74, N = 51, r: Coefficient of phenotypic correlation with grain yield, EPP: Ear per plant, EASP: Ear aspect, EHT: Ear height, AD: Anthesis days, PHT: Plant height, LR: Leaf rolling, SEN: Leaf senescence, LE: Leaf erectness, TS: Tassel size; PR: Plant recovery, ASI: Anthesis silking | |
Anthesis days was therefore indicated as a reliable trait for indirect selection for improved grain yield under drought stress.
Anthesis-Silking Interval (ASI) and grain yield: The phenotypic correlation coefficient was negative and low (-0.13) between the two traits. The magnitude of direct effect of ASI on grain yield was low (0.01). This result contrasts with results from several drought tolerance studies in maize6,16. This unexpected result was explained by the relative high number of missing values recorded for silking days, since some plots under drought stress failed to reach 50% silking approximately 20-30 days after 50% anthesis and were declared missing values. Usefulness of ASI as an indirect selection trait for grain yield under drought stress was suggested but not conclusively established in this study though high heritability (0.59) was found for this trait under drought.
Plant height and grain yield: The phenotypic correlation coefficient was positive and low (0.13) between the two traits but the direct effect of plant height was negative and high which was also found under severe drought conditions by Khalili et al.22, indicating that hybrids with high plant height were more sensitive to drought stress than medium plant height. The indirect positive and high effect of ear height and ear aspect are the possible cause of positive and low correlation between plant height and grain yield. Therefore, these traits must be considered if selection is made through plant height. Similar result was reported by Ahmad and Saleem19 in non-drought condition.
Ear height (EHT) and grain yield: The phenotypic correlation between EHT and grain yield and direct effect of ear height on grain yield were both positive and almost equal in magnitude (Table 3). Shi et al.23 reported also positive direct effect of EHT on single ear yield of maize inbred lines under medium drought conditions. Under drought stress, EHT had positive and high direct effect and negative indirect effect via plant height on grain yield indicating that the ratio between ear and plant height (R-EPH) was associated to grain yield which was calculated and; significant and positive phenotypic correlation was found between R-EPH and grain yield (r = 0.44**). Moreover high heritability was found for EHT in the present study under drought as well as under well-watered conditions, indicating that characteristic of ear position should be considered in selection of high yielding drought tolerant genotypes.
Leaf Rolling (LR) and senescence (SEN) and grain yield: The phenotypic correlation coefficient and the heritability of LR and SEN were all low (Table 2). This result showed that any progress can be made by including these traits in selection index for selecting high yielding drought tolerant hybrids. Bolanos and Edmeades5 reported also a low genetic correlation of grain yield with leaf rolling and leaf senescence.
Leaf Erectness (LE) and grain yield: The direct effect of leaf erectness was negative (-0.06). The phenotypic correlation between LE and grain yield with grain yield was significant and negative (r = -0.29*), however the heritability under drought was higher (0.55). Similar result was reported by Bolanos and Edmeades5 for the heritability.
Tassel Size (TS) and grain yield: A negative direct effect of TS on grain yield as well as negative and significant phenotypic correlation between TS and grain yield were found (Table 3). Moreover this trait showed a high heritability (0.58), indicating that TS can be included in the selection index as an indirect selection trait for grain yield under drought conditions. However the high negative contribution to grain yield was found through ear aspect, therefore this trait should also be included in the selection index.
Plant Recovery (PR) and grain yield: Results showed a significant and negative phenotypic correlation between PR score and grain yield (r = -0.33**). The magnitude of the direct effect on grain yield was negatively high (-015). The same trend was observed for the heritability (0.60). The use of plant recovery as secondary trait would be effective for selection of drought tolerant genotypes. The PR had important indirect effect on grain yield through EASP (- 0.18), EHT (- 0.16) and AD (0.12). Thus, those traits should also be considered in the selection index.
Selection for drought tolerance: Results showed that important genetic variability for grain yield and secondary traits existed among the hybrids.
| Table 4: | Procedure for classifying hybrids into four categories of degree of drought resistance |
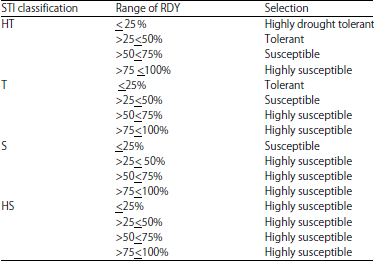 | |
HT: Highly drought tolerant, R: Drought tolerant, S: Drought susceptible, HS: Highly drought susceptible | |
This variability will facilitate selection for drought tolerant hybrids with high yield potential, either by drought escape mechanisms or by using secondary traits in a selection index. The relative value of secondary traits for indirect selection for a grain yield is determined based on variance and heritability and on genetic correlation with yield24. Then Ear Per Plant (EPP), Tassel Size (TS) and Plant Recovery (PR) were identified as important indirect selection criteria which were suggested with the Grain Yield (GY) to use in the selection for drought tolerant hybrids. In addition EASP and R-EPH should also be added. They are not stress-adaptive traits by themselves but EPP, TS and PR had important indirect effect on grain yield via EASP. Moreover ear aspect had a high correlation coefficient with grain yield and a high direct effect on grain yield. The ratio, R-EPH would avoid selection of genotypes with undesirable ear position and plant height characteristics.
The function of the selection index will be as follow:
Selection Tolerance Index (STI) = 5 f (GY)+3 f (EPP)-2 f (TS)-2 f (PR)-2 f (EASP)+1 f (R-EPH)
In maize, weights typically allocated to secondary traits are +3, -2, -2, -2 and -1 for ears per plant, ASI, leaf senescence, tassel size and leaf rolling, respectively24, however based on the results of path analysis in the present study different traits and weights were chosen in the applied selection index. The weight of GY is higher because the heritability for grain was higher in WS than the corresponding in WW conditions, indicating stability of grain yield6.
Base on the Selection Tolerance Index (STI) values, the hybrids were classified into four groups based on the degree of drought tolerance: Highly drought tolerant “HT”, drought tolerant “T”, drought susceptible “S” and highly drought-susceptible “HS”. The Selection Tolerance Index (STI) does not consider the relative yield reduction under drought stress, thus a genotype classified as highly tolerant based on STI value may have high percent of yield reduction under drought stress which is not desirable. Therefore, the classification of the hybrids into four categories of drought tolerance based on STI value was improved by incorporating the relative decrease in yield under drought (RDY) index. The selection procedure involving the two selection criteria, STI and RDY is presented in Table 4. Based on this selection procedure, of the 48 hybrids evaluated 6 were highly drought tolerant, 18 drought tolerant, 10 drought susceptible and 14 highly drought susceptible (data not shown).
CONCLUSION
The study identified ear per plant, tassel size and plant recovery as important indirect selection criteria for grain yield under drought conditions. The usefulness of ASI, LR, LS and LE as indirect selection traits for grain yield under drought stress were not demonstrated in this study. The path coefficient analysis showed that the traits identified as useful secondary traits were significantly correlated to grain yield but, their direct effect on grain yield were low and/or had important contribution to grain yield through other traits. Therefore, these traits should also be considered in selection for drought tolerance.
ACKNOWLEDGMENT
We thank our field staff of INERA research station at Valley Du Kou for plot management and assistance with data collection. We also thank D. Elola, master student, for supervision of the trial and assistance in data collection. This work was founded by West Africa Centre for Crop Improvement (WACCI).
REFERENCES
- Ashraf, M., 2010. Inducing drought tolerance in plants: Recent advances. Biotechnol. Adv., 28: 169-183.
CrossRefPubMedDirect Link - Bruce, W.B., G.O. Edmeades and T.C. Barker, 2002. Molecular and physiological approaches to maize improvement for drought tolerance. J. Exp. Bot., 53: 13-25.
CrossRefPubMedDirect Link - Ali, F., N. Kanwal, M. Ahsan, Q. Ali, I. Bibi and N.K. Niazi, 2015. Multivariate analysis of grain yield and its attributing traits in different maize hybrids grown under heat and drought stress. Scientifica.
CrossRefDirect Link - Bolanos, J. and G.O. Edmeades, 1996. The importance of the anthesis-silking interval in breeding for drought tolerance in tropical maize. Field Crops Res., 48: 65-80.
CrossRefDirect Link - Almeida, G.D., D. Makumbi, C. Magorokosho, S. Nair and A. Borem et al., 2013. QTL mapping in three tropical maize populations reveals a set of constitutive and adaptive genomic regions for drought tolerance. Theoret. Applied Genet., 126: 583-600.
CrossRefDirect Link - Banziger, M., G.O. Edmeades, D.L. Beck and M.R. Bellon, 2000. Breeding for Drought and Nitrogen Stress Tolerance in Maize: From Theory to Practice. CIMMYT, Mexico, ISBN: 970-648-46-3, Pages: 68.
Direct Link - Hao, Z.F., X.H. Li, Z.J. Su, C.X. Xie and M.S. Li et al., 2011. A proposed selection criterion for drought resistance across multiple environments in maize. Breed. Sci., 61: 101-108.
CrossRefDirect Link - Khalily, M., M. Moghaddam, H. Kanouni and E. Asheri, 2010. Dissection of drought stress as a grain production constraint of maize in Iran. Asian J. Crop Sci., 2: 60-69.
CrossRefDirect Link - Pham, H.T.T., K.T. Do, M.N. Truong, X.D. Tran, L.T. Nguyen and B.C. Bui, 2016. Path analysis for yield traits in F2 generation and molecular approaches for breeding rice tolerant to drought and submergence. Afr. J. Agric. Res., 11: 2329-2336.
CrossRefDirect Link - Cramer, C.S., T.C. Wehner and S.B. Donaghy, 1999. PATHSAS: A SAS computer program for path coefficient analysis of quantitative data. J. Heredity, 90: 260-262.
CrossRefDirect Link - Farshadfar, E., M.M. Poursiahbidi and S.M. Safavi, 2013. Assessment of drought tolerance in land races of bread wheat based on resistance/tolerance indices. Int. J. Adv. Biol. Biomed. Res., 1: 143-158.
Direct Link - Dixit, S., B.E. Huang, M.T.S. Cruz, P.T. Maturan, J.C.E. Ontoy and A. Kumar, 2014. QTLs for tolerance of drought and breeding for tolerance of abiotic and biotic stress: An integrated approach. PLoS ONE, Vol. 9.
CrossRefDirect Link - Ramya, P., G.P. Singh, N. Jain, P.K. Singh and M.K. Pandey et al., 2016. Effect of recurrent selection on drought tolerance and related morpho-physiological traits in bread wheat. PLoS ONE, Vol. 11.
CrossRefDirect Link - Messmer, R., Y. Fracheboud, M. Banziger, M. Vargas, P. Stamp and J.M. Ribaut, 2009. Drought stress and tropical maize: QTL-by-environment interactions and stability of QTLs across environments for yield components and secondary traits. Theoret. Applied Genet., 119: 913-930.
CrossRefDirect Link - Badu-Apraku, B., R.O. Akinwale, J. Franco and M. Oyekunle, 2012. Assessment of reliability of secondary traits in selecting for improved grain yield in drought and low-nitrogen environments. Crop Sci., 52: 2050-2062.
CrossRefDirect Link - Filipovic, M., M. Babic, N. Delic, G. Bekavac and V. Babic, 2014. Determination relevant breeding criteria by the path and factor analysis in maize. Genetika, 46: 49-58.
CrossRefDirect Link - Ahmad, A. and M. Saleem, 2003. Path coefficient analysis in Zea mays L. Int. J. Agric. Biol., 5: 245-248.
Direct Link - Magorokosho, C., K.V. Pixley and P. Tongoona, 2003. Selection for drought tolerance in two tropical maize populations. Afr. Crop Sci. J., 11: 151-161.
Direct Link - Chapman, S.C. and G.O. Edmeades, 1999. Selection improves drought tolerance in tropical maize populations: II. Direct and correlated responses among secondary traits. Crop Sci., 39: 1315-1324.
CrossRefDirect Link - Shi, G.Z., Z. Zhao, W.X. Zhang and Y.Q. Yuan, 2008. Genetic correlation and path analysis of agronomic traits for maize inbred lines under drought stress. Southwest China J. Agric. Sci., 21: 570-574.
Direct Link - Araus, J.L., M.D. Serret and G.O. Edmeades, 2012. Phenotyping maize for adaptation to drought. Front. Physiol., Vol. 3.
CrossRefDirect Link