
Lagzouli Mohamed
Laboratory of Agro-physiology, Biotechnologies and Quality, Sciences College, University IBN Tofail, BP 133, 14000 Kenitra, Morocco
Youssfi El Kettani
C.I.R.O.S Laboratory, Department of Mathematics, Sciences College, University IBN Tofail, BP 133, 14000 Kenitra, Morocco
Aitounejjar Ali
National Institute of Agronomic Research, P.O. Box 589, Settat 26000, Morocco
Elyachioui Mohamed
Laboratory of Agro-physiology, Biotechnologies and Quality, Sciences College, University IBN Tofail, BP 133, 14000 Kenitra, Morocco
Jadal Mohamed
Laboratory of Agro-physiology, Biotechnologies and Quality, Sciences College, University IBN Tofail, BP 133, 14000 Kenitra, Morocco
Pakistan Journal of Biological Sciences
Year: 2017 | Volume: 20 | Issue: 2 | Page No.: 100-107







ABSTRACT
Objective: Glucoamylase is among the most important enzymes in biotechnology. The present study aims to determine better conditions for growth and glucoamylase production by Candida guilliermondii and to reduce the overall cost of the medium using Box-Behnken design with one central point and response surface methodology. Methodology: Box-Behnken factorial design based on three levels was carried out to obtain optimal medium combination of five independent variables such as initial pH, soluble starch, CH4N2O, yeast extract and MgSO4. Forty one randomized mediums were incubated in flask on a rotary shaker at 105 rpm for 72 h at 30°C. Results: The production of biomass was found to be pH and starch dependent, maximum production when the starch concentration was 8 g L–1 and the initial pH was 6, while maximum glucoamylase production was found at 6.5 of initial pH, 4 g L–1 yeast extract and 6 g L–1 starch, whereas yeast extract and urea were highly significant, but interacted negatively. Box-Behnken factorial design used for the analysis of treatment combinations gave a second-order polynomial regression model with R2 = 0.976 for Biomass and R2 = 0.981 for glucoamylase. Conclusion: The final biomass and glucoamylase activity obtained was very close to the calculated parameters according to the p-values (p<0.001), the predicted optimal parameters were confirmed and provides a basis for further studies in baking additives and in the valuation of starch waste products.
PDF Abstract XML References Citation
Received: October 30, 2016;
Accepted: December 13, 2016;
Published: January 15, 2017
Copyright: © 2017. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Lagzouli Mohamed, Youssfi El Kettani, Aitounejjar Ali, Elyachioui Mohamed and Jadal Mohamed, 2017. Application of Response Surface Methodology for Optimization of Extracellular Glucoamylase Production by Candida guilliermondii. Pakistan Journal of Biological Sciences, 20: 100-107.
DOI: 10.3923/pjbs.2017.100.107
URL: https://scialert.net/abstract/?doi=pjbs.2017.100.107
DOI: 10.3923/pjbs.2017.100.107
URL: https://scialert.net/abstract/?doi=pjbs.2017.100.107
INTRODUCTION
Amylases are among the most important enzymes used in biotechnology, particularly in processes involving starch hydrolysis. Though amylases originate from different sources (plants, animals and microorganisms) the microbial amylases are the most used in food, biofuels and detergents industries. Natural fermented media (foods, soils and wastes) offers sources for isolation of microorganism strains producing amylases, many strains used in the food industry are isolated from fermented food media1.
Generally, two main methods are used for the process of optimization, which are usually referred to as the classical and statistical methods. The classical method is based on the ‘One-factor-at-a-time’ method, in which one independent variable is observed, whereas the other factors are kept at a fixed level. However, this method cannot guarantee the determination of optimal conditions and is unable to study the interactions between the factors, thus probably leads to unreliable results and inaccurate conclusions2. The statistical optimization method (response surface methodology) uses the data from a few sets of experiments to determine equations, this method can overcome the limitations of the classical method, it has been proved to be a powerful tool for designing experiments, building models, evaluating the effects of factors and analyzing optimal conditions of factors for desirable responses3.
Response Surface Methodology (RSM) has already been effectively applied during optimization of different media and culture conditions for the production of microbial metabolites, such as, in the improvement of biomass production and glucoamylase activity by Candida famata4 and in the optimization of α-amylase production by Aspergillus niger5 and by Aspergillus oryzae6. These designs were also used for the optimization of the culture medium for the production of β-galactosidase from Kluyveromyces marxianus7.
The present study aims to determine better conditions for growth and glucoamylase productivity by Candida guilliermondii. It used of response surface methodology and Box-Behnken8 design with one central point to optimize media composition and reduce the overall cost of the medium, providing a basis for further studies in the baking additives and in the valuation of starch waste products.
MATERIALS AND METHODS
Microorganism used: Candida guilliermendii was isolated from traditional moroccan sourdough using medium containing soluble starch (5 g L–1), KH2PO4 (3 g L–1), (NH4)2SO4 (1 g L–1), MgSO4 (0.5 g L–1) and yeast extract (4 g L–1). The initial pH was adjusted to 5.0 with HCl 0.1 M. The medium was solidified by the addition of 1.5% Agar and autoclaved at 121°C for 15 min. Liquid medium was incubated in flask on a rotary shaker at 105 rpm for 72 h at 30°C.
Cultivation and production of glucoamylase by Candida guilliermendii
Growth rate and enzyme assays: Growth rate was determined after 72 h of incubation by measuring the absorbance of the suspension at 600 nm, to estimate enzyme activity a free substrate supernatant by 7000 rpm for 10 min centrifugation was used. Glucoamylase activity was determined by measuring the reducing sugar formed by the enzymatic hydrolysis of starch using the method of Somogyi and Nelson9, 0.25 mL soluble starch (1%), 0.15 mL phosphate buffer ( 0.1M) and 0.1 mL enzyme solution were mixed then incubated at 40°C in water bath for 30 min. The reaction was stopped by 2 mL of Somogiy reactive and 1.5 mL of distilled water, followed by boiling for 15 min to develop blue color. The absorbance was measured at 540 nm with a spectrophotometer against the control in which no enzyme was added. A calibration curve of absorbance and concentration of glucose was established with known amount of glucose.
One unit (μmol L–1 min–1) of amylase was defined as the amount of μmol of reducing sugar per liter of enzymes per min, measured as glucose under the conditions of assay.
Box-Behnken design experiments: A 3-level 5-factor Box-Behnken design was adopted to evaluate the effects of initial pH (X1) starch (X2), CH4N2O(X3), yeast extract (X4) and MgSO4 (X5) on the biomass and glucoamylase production by Candida guilliermondii. In this study, the independent variables were studied at three different levels; namely; low (-1), medium (0) and high (+1), providing 41 trials (mediums). Levels of variables chosen for the Box-Behnken optimization experiment are presented in Table 1.
| Table 1: | Levels of variables chosen for the Box-Behnken optimization experiment |
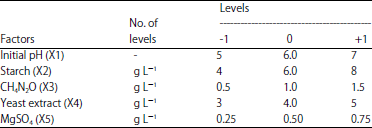 | |
The response variable was fitted by a second-order model in order to correlate the response variables to the independent variables, the second-order polynomial coefficients were calculated and analyzed using the adequate statistical software. The general form of the second-degree polynomial equation is:
where, Y is the predicted response, b0 the intercept, bi the linear coefficient, bij the quadratic coefficient, bii is the linear-by-linear interaction between Xi and Xj regression coefficients and Xi Xj are input variables that influence the response variable Y.
Statistical analysis: Statistical analysis of the model was performed to evaluate the analysis of variance (ANOVA), this analysis included Fisher’s F test (overall model significance), its associated probability p(F), correlation coefficient R, determination coefficient R2 which measure the goodness of fit of regression model. The quadratic models were represented as contour plots (2D) and response surface curves (3D) for each variable. The 41 randomized experiments with the coded and real values of the experimental variables are given in Table 2.
RESULTS
The experimental and calculated results of experiments carried out with the Box-Behnken design are given in Table 3. The analysis of variance (ANOVA) was calculated for each responses to determine significant parameters, then was carried out by Fisher’s statistical test for the analysis of variance.
The corresponding analysis of variance (ANOVA) employed to determine signification was presented in Table 4. The ANOVA of the quadratic regression model demonstrates that the model is highly significant, as it is evident in the two results from the Fisher’s test, with a very high probability value (p<0.001).
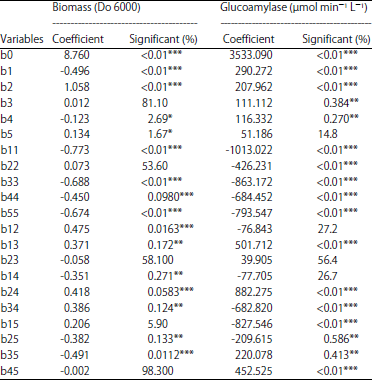
The model presents a high determination coefficient (R2 = 0.976) for (Biomass) explaining 97% of the variability in the response, linear regression (R2 = 0.981) of glucoamylase production explains 98% of the variability in the response. Effects of factors upon the biomass and glucoamylase production are reported in Table 5.
| Table 2: | Coded levels (in parentheses) and real values of experimental variables |
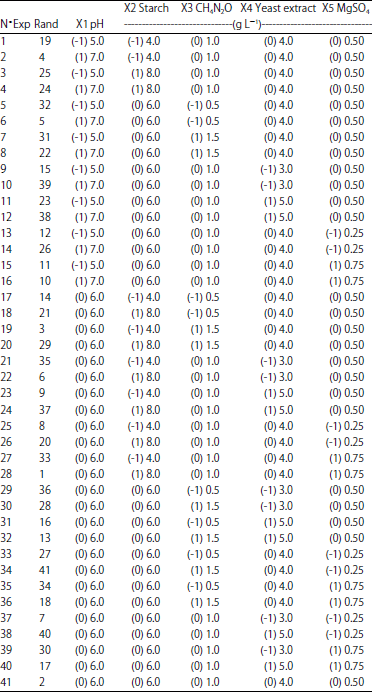 | |
Among the five variables tested, starch (b2) and MgSO4 (b5) were very significant; those variables increased the biomass production as they have a large positive coefficient of linear effect on the contrary, urea had no effect on the biomass production (p = 81.1), while yeast extract showed a negative effect (-0.123) (p = 2.69).
The probability values of coefficients of linear effects of pH, starch, urea and yeast extract on glucoamylase production were very significant. Initial pH, soluble starch were highly significant, according to their p values (p<0.001) indicating that the model was extremely affected by these variables; followed by yeast extract (p = 0.270) and urea (p = 0.384). However, despite the positive effect of urea and yeast extract on glucoamylase production, they exhibited a negative interaction (-682.820).
The p-value of the linear effect of MgSO4 (b5) was very high (p = 14.8) explaining that MgSO4 has no effect on the glucoamylase production, but interact positively with urea and yeast extract.
| Table 3: | Box-Behnken design for the production of calculated and experimental yields of biomass and glucoamylase |
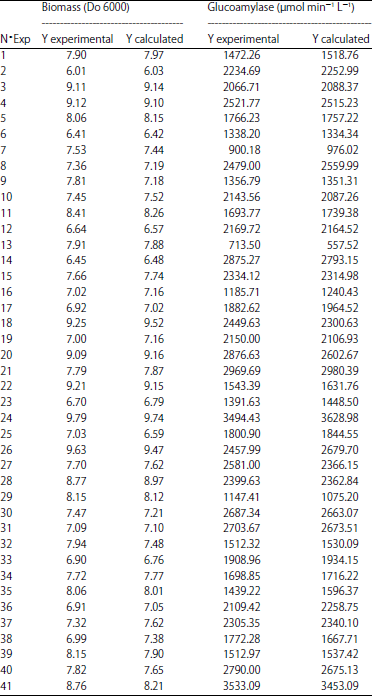 | |
Furthermore, interaction between soluble starch and yeast extract (b24) and between initial pH and urea (b13) were positively significant.
Contour plots of initial medium pH (b1) and starch (b2) effect on the biomass production are not elliptical in Fig. 1. Maximum biomass production was reached at the higher concentration of soluble starch (8 g L–1), it seems that maximum biomass would be reached beyond (8 g L–1) of starch; medium level of initial pH (pH = 6) provided maximum biomass and further increase in the initial pH value resulted in a decrease of the biomass production.
Surface plots of the interaction effect between initial pH and yeast extract indicated an elliptical shape with positive effect (Fig. 2), glucoamylase production increased gradually with an increment of the two variables to reach to a maximum value at pH = 6.5 and at 4 g L–1 of yeast extract, beyond these two values glucoamylase production decreased.
The response surface in Fig. 3 shows a net elliptical peak at 6.5 g L–1 of starch concentration and 6.5 of initial pH value. Beyond those two values, glucoamylase production decreased. The elliptical contour plots make out that the interaction between the initial pH and starch was significant on glucoamylase activity.
Perfect elliptical plot in Fig. 4 indicates a high interaction between urea and magnesium sulfate, the contour plot is almost helicoids, maximum glucoamylase activity shown with 1 g L–1 of urea and 0.5 g L–1 of magnesium sulfate, beyond the central point of both ingredient production of glucoamylase decreased.
The mathematical model relating the production of glucoamylase with the independent process variables, b1, b2, b3, b4 and b5 using results displayed in Table 5 is given in the following second order polynomial:
| Y | = | 3533.090b0+290.272b1+207.962b2+111.112b3+116.332b4+51.186b5-013.022b11-426.231b22-863.172b33- 684.452b44-793.547b55-76.843b12+ 501.712b13-39.905b23-77.705b14+882.275b24-682.820b34- 827.546b15-209.615b25+220.078b35+452.525b45 |
where, Y is the response variable, glucoamylase activity (μmol min–1 L–1) and b1, b2, b3, b4 and b5 the values of independent variables, initial pH, soluble starch, urea, yeast extract and MgSO4.
| Table 4: | ANOVA for Box-Behnken model results |
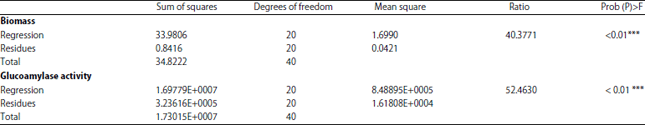 | |
| ***Very high signification degree | |
Candida guilliermondii cultivated under optimum conditions in the bioreactor for 3 days produced 5055.56 μmol min–1 L–1 of glucoamylase activity. This finding showed that the production of glucoamylase activity process under optimum condition determined was well reproducible.
| Table 5: | Regression coefficients of biomass and extracellular glucoamylase production model |
 | |
| *Signification degree, **High signification degree, ***Very high signification degree | |
DISCUSSION
The main goal of response surface is hunt efficiently for the optimum values of variables; each contour curve represents an infinitive number of combinations of two test variables. The maximum predicted value was indicated by the surface confined in the smallest ellipse in the contour diagram, Elliptical contours are obtained whenever there is a perfect interaction between the independent variables10.
Optimal pH is very important for growth of the microorganism and its metabolic activities, glucoamylase and biomass produced by Candida guilliermondii were affected by varying the pH of the medium. Biomass production was highly occurred at pH = 6.0, which are comparable to previously reported results, in the same way, the effect of initial medium pH on the production of biomass was reported by Keera et al.11 and with Aspergillus niger and Aspergillus flavus12.
The production of glucoamylase by Candida guilliermondii was found maximum at pH 6.5, beyond this pH, amylase production decreased gradually. Abeer found that gluamylase production by Aspergillus oryzae FK-923 was at pH 5.511, while Esra Uçkun Kiran found that high glucoamylase production at an initial pH of 7.913.
The study shows the prominent role played by starch as a carbon source for biomass and glucoamylase production14,15. Maximum biomass production was observed at 8 g L–1 of soluble starch; 3D indicated that maximum biomass production would be reached beyond 8 g L–1, whereas the maximum of glucoamylase production was obtained by 4.1 g L–1 of starch with Pinicillium notatum16, whereas other studies showed maximum glucoamylase activity by thermophilic fungus Humicola grisea MTCC 352 at 28.41 g L–1 14.
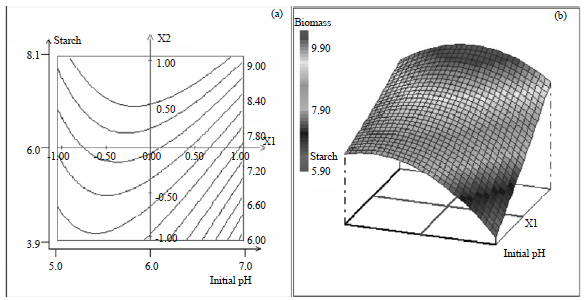 | |
| Fig. 1(a-b): | Contour and 3D plot of effects of (a) Initial pH and (b) soluble starch on biomass production |
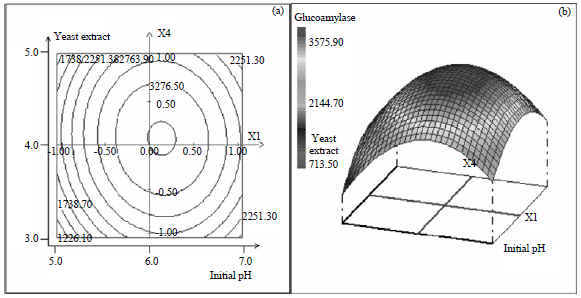 | |
| Fig. 2(a-b): | Contour and 3D plot of effects of (a) Initial pH and (b) Yeast extract on glucoamylase production |
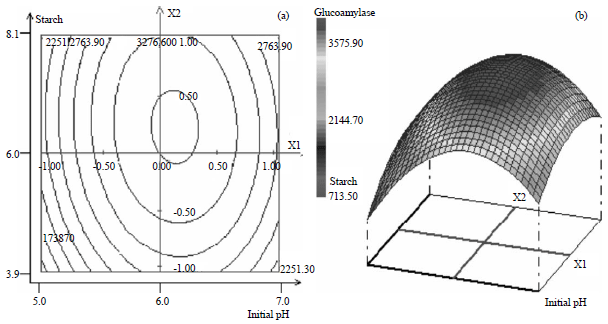 | |
| Fig. 3(a-b): | Contour and 3D plot of effects of (a) Initial pH and (b) Soluble starch on glucoamylase production |
Starch seems to have an "Inductive effect" and portrays a significant role in the production of starch-saccharifying amyloglucosidase by Aspergillus species17.
It found that maximum glucoamylase activity attained at 4 g L–1 of yeast extract concentration and slightly decreased beyond 4 g L–1, similar kind of results was obtained for glucoamylase production, which prove the positive effect played by yeast extract14,6, yeast extract helped in the development of mycelial structures with a corresponding higher yield of enzymes15, However, some studies showed that the peptone was the best nitrogen source with Bacillus licheniformis and Penicillium fellutanum18.
The similar inhibitory effect of the higher levels of yeast extract on α-amylase produced by Aspergillus niger WLB42 was reported by Wang et al.19. These observations indicate that Candida guilliermondii is sensitive to the variation of yeast extract level in the medium. It denoted a negative interaction between yeast extract and urea (-682.820), complex nitrogen sources at higher concentrations might have a toxic effect on enzyme production19.
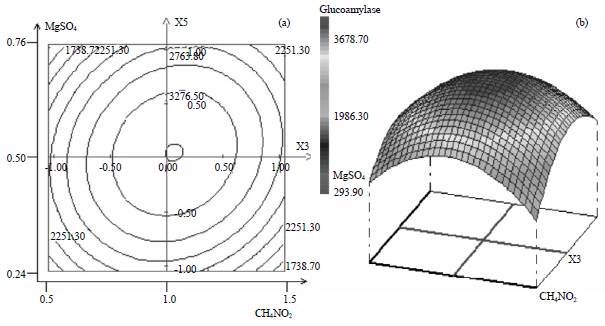 | |
| Fig. 4(a-b): | Contour and 3D plot of effects of (a) CH4N2O and (b) MgSO4 on glucoamylase production |
Glucoamylase production was affected by urea as nitrogen source, similar results of the beneficial effects of urea on amylase production was confirmed by Aspergillus oryzea11 and by Aspergillus niger 20.
CONCLUSION
In this study, Box-Behnken Design and response surface methodology were performed to optimize biomass and glucoamylase production by Candida guilliermondii. The significant achievement of the present study lies in the fact that maximum production of Biomass occurred when the starch concentration is 8 g L–1 and the initial pH is 6.0; While maximum glucoamylase production was found at 6.5 of initial pH, 4 and 6 g L–1 of starch, whereas yeast extract and urea were highly significant, but interacted negatively. The predicted values were verified experimentally and gave a second-order polynomial regression model. In the present study, the value of R2 was R2 = 0.976 for Biomass and R2 = 0.981 for glucoamylase production which indicates good correlation between observed and experimental values of both responses. The high value (close to 1) of R2 indicates good correlation between predicted and observed values. These predicted optimal parameters were confirmed in the laboratory and the final biomass and glucoamylase activity obtained was very close to the calculated parameters.
The optimized media composition found out in the present investigation might reduce the overall cost of the medium and provides a basis for further studies as a potential candidate for application in the baking additive, in the detergent industry and in the valuation of starch waste products.
ACKNOWLEDGMENTS
Authors express their acknowledgments to the Laboratory of Microbial Biotechnology (LMB), Sciences College, University IBN TOFAIL, for its contribution to the present work.
REFERENCES
- Hu, Y., H. Qin, Z. Zhan, Y. Dun and Y. Zhou et al., 2016. Optimization of Saccharomyces boulardii production in solid-state fermentation with response surface methodology. Biotechnol. Biotechnol. Equip., 30: 173-179.
CrossRefDirect Link - Wu, W., S. Zhao, C. Chen, F. Ge, D. Liu and X. He, 2014. Optimization of production conditions for antioxidant peptides from walnut protein meal using solid-state fermentation. Food Sci. Biotechnol., 23: 1941-1949.
CrossRefDirect Link - Mosbah, H., I. Aissa, N. Hassad, D. Farh, A. Bakhrouf and S. Achour, 2015. Improvement of biomass production and glucoamylase activity by Candida famata using factorial design. Biotechnol. Applied Biochem., 63: 572-580.
CrossRefDirect Link - Oberoi, H.S., R. Rawat and B.S. Chadha, 2014. Response surface optimization for enhanced production of cellulases with improved functional characteristics by newly isolated Aspergillus niger HN-2. Antonie Leeuwenhoek, 105: 119-134.
CrossRefDirect Link - Naili, B., M. Sahnoun, S. Bejar and R. Kammoun, 2016. Optimization of submerged Aspergillus oryzae S2 α-amylase production. Food Sci. Biotechnol., 25: 185-192.
CrossRefDirect Link - Nachaiwieng, W., S. Lumyong, K. Yoshioka, T. Watanabe and C. Khanongnuch, 2015. Bioethanol production from rice husk under elevated temperature simultaneous saccharification and fermentation using Kluyveromyces marxianus CK8. Biocatal. Agric. Biotechnol., 4: 543-549.
CrossRefDirect Link - Feng, Y., X. Zhao, F. Lv, J. Zhang and B. Deng et al., 2015. Optimization on preparation conditions of salidroside liposome and its immunological activity on PCV-2 in mice. Evidence-Based Complemen. Altern. Med.
CrossRefDirect Link - Keera, A.A., F.E. Mouafi, T. Kahil, M. Fadel and A.A. Abedo, 2014. Hyper production of glucoamylase by Aspergillus oryzae FK-923 under solid state fermentation. World Applied Sci. J., 30: 1447-1461.
Direct Link - Colla, L.M., A.L. Primaz, S. Benedetti, R.A. Loss and M. de Lima et al., 2016. Surface response methodology for the optimization of lipase production under submerged fermentation by filamentous fungi. Braz. J. Microbiol., 47: 461-467.
CrossRefDirect Link - Kiran, E.U., A.P. Trzcinski and Y. Liu, 2014. Glucoamylase production from food waste by solid state fermentation and its evaluation in the hydrolysis of domestic food waste. Biofuel Res. J., 1: 98-105.
CrossRefDirect Link - Ramesh, V. and V.R. Murty, 2014. Sequential statistical optimization of media components for the production of glucoamylase by thermophilic fungus Humicola grisea MTCC 352. Enzyme Res.
CrossRefDirect Link - Arnthong, J., B. Wanitchaploy, K. Sakai, J.J. Sanglier and V. Kitpreechavanich, 2015. Statistical screening of factors affecting glucoamylase production by a thermotolerant rhizopus microsporus TISTR 3518 using plackett-burman design. Afr. J. Biotechnol., 9: 7312-7316.
Direct Link - Ghosh, P., A. Das, S. Gayen, K.C. Mondal and U. Ghosh, 2015. Statistical optimization of α-amylase production from Penicillium notatum NCIM 923 and kinetics study of the purified enzyme. Acta Biol. Szegedien., 59: 179-188.
Direct Link - Pervez, S., N.N. Siddiqui, A. Ansari, A. Aman and S.A.U. Qader, 2015. Phenotypic and molecular characterization of Aspergillus species for the production of starch-saccharifying amyloglucosidase. Ann. Microbiol., 65: 2287-2291.
CrossRefDirect Link - Lakshmi, M.V.V.C. and P. Jyothi, 2014. Production and optimization of glucoamylase from aspergillus oryzae NCIM 1212 using wheat bran, varying chemical parameters under solid state fermentation. Int. J. Curr. Microbiol. Applied Sci., 3: 70-76.
Direct Link - Wang, S., C. Lin, Y. Liu, Z. Shen, J. Jeyaseelan and W. Qin, 2016. Characterization of a starch-hydrolyzing α-amylase produced by Aspergillus niger WLB42 mutated by ethyl methanesulfonate treatment. Int. J. Biochem. Mol. Biol., 7: 1-10.
Direct Link - Lima-Orozco, R., I. van Daele, U. Alvarez-Hernandez and V. Fievez, 2014. Combined conservation of jack bean and velvet bean with sorghum: Evaluation of lab-scale silages and in vitro assessment of their nutritive value. J. Agric. Sci., 152: 967-980.
CrossRefDirect Link