
Raed S. Said
Department of Anatomy, Faculty of Medicine, Jordan University of Science and Technology, 22110 Irbid, Jordan
Ayman G. Mustafa
Department of Anatomy, Faculty of Medicine, Jordan University of Science and Technology, 22110 Irbid, Jordan
Hasan A. Asfour
Department of Anatomy, Faculty of Medicine, Jordan University of Science and Technology, 22110 Irbid, Jordan
Emad I. Shaqoura
Department of Anatomy, Faculty of Medicine, Jordan University of Science and Technology, 22110 Irbid, Jordan
Pakistan Journal of Biological Sciences
Year: 2017 | Volume: 20 | Issue: 1 | Page No.: 1-11







ABSTRACT
Adult skeletal muscle is a post-mitotic terminally differentiated tissue that possesses an immense potential for regeneration after injury. This regeneration can be achieved by adult stem cells named satellite cells that inhabit the muscular tissue. These cells were first identified in 1961 and were described as being wedged between the plasma membrane of the muscle fiber and the surrounding basement membrane. Since their discovery, many researchers investigated their embryological origin and the exact role they play in muscle regeneration and repair. Under normal conditions, satellite cells are retained in a quiescent state and when required, these cells are activated to proliferate and differentiate to repair pre-existing muscle fibers or to a lesser extent fuse with each other to form new myofibers. During skeletal muscle regeneration, satellite cell actions are regulated through a cascade of complex signaling pathways that are influenced by multiple extrinsic factors within the satellite cell micro-environment. Here, the basic concepts were studied about satellite cells, their development, function, distribution and the different cellular and molecular mechanisms that regulate these cells. The recent findings about some of their clinical applications and potential therapeutic use were also discussed.
PDF Abstract XML References Citation
Received: August 30, 2016;
Accepted: November 26, 2016;
Published: December 15, 2016
Copyright: © 2017. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Raed S. Said, Ayman G. Mustafa, Hasan A. Asfour and Emad I. Shaqoura, 2017. Myogenic Satellite Cells: Biological Milieu and Possible Clinical Applications. Pakistan Journal of Biological Sciences, 20: 1-11.
DOI: 10.3923/pjbs.2017.1.11
URL: https://scialert.net/abstract/?doi=pjbs.2017.1.11
DOI: 10.3923/pjbs.2017.1.11
URL: https://scialert.net/abstract/?doi=pjbs.2017.1.11
PREVIOUS STUDIES
Postnatal growth of skeletal muscle is mainly attributed to a group of adult stem cells known as satellite cells (SCs)1. The SCs are wedged between the basal lamina and the sarcolemma of the skeletal muscle fiber (Fig. 1). These spindle shaped, mononucleated cells were first observed by Mauro and Katz while studying the muscle of the frog2,3 in 1961. The satellite cell long axis is parallel to the long axis of the muscle fiber and their length may reach 25 μm but its width is less than 5 μm. Although some other non-satellite stem cells may also contribute in skeletal muscle growth and regeneration, the satellite cell group remains the major source of postnatal muscle growth, repair and maintenance4.
The purpose of this study is to report the most recent research findings regarding all biological aspects of SCs. It aims to cover the basic concepts about satellite cells, their development, function, distribution and the different cellular and molecular mechanisms that regulate their fate. This study also intends to discuss the recent findings regarding the clinical applications of SCs and their potential therapeutic use.
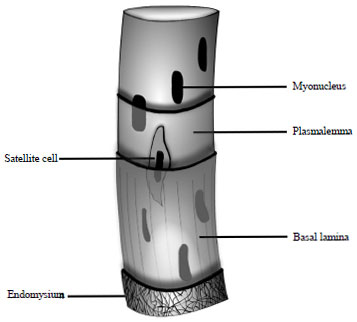 | |
| Fig. 1: | A satellite cell (SC) and myonuclei within a muscle fiber. It shows from deep to superficial, the plasmalemma, basal lamina and endomysium layers that cover the muscle fiber. Myonuclei are located within the fiber beneath the plasmalemma, while the SC is sandwiched between the plasmalemma and basal lamina. Source: Allouh7 |
DEVELOPMENTAL ORIGIN OF SATELLITE CELLS
During myogenesis, myogenic precursor cells, known as myoblasts, differentiate from mesodermal cells derived from the paraxial somites and migrate to their final destination in the developing muscles. After that, many myoblasts fuse together forming myotubes which mature into muscle fibers. However, some of these myogenic precursors do not differentiate into myotubes and reside beneath the basal lamina of the muscle fiber to become SCs postnatally5.
The concept that SCs originate from the somite as a distinct lineage of myogenic precursors, or as a preexisting lineage, such as embryonic or fetal myoblasts is still an area of investigation. A previous study had revealed that clones of myogenic stem cells that closely resemble satellite-cell-derived myogenic precursors are isolated from the dorsal aorta of mouse embryos and express myogenic and endothelial markers that are also expressed in adult SCs6. The development of the dorsal aorta from migratory angioblasts derived from the paraxial mesoderm of the somites suggests that SCs can develop from either a non-myogenic origin, such as endothelial cell precursors or a common precursor for both SCs and endothelial cells6.
SATELLITE CELL CYCLE
Satellite cells are present in a quiescent state within the mature skeletal muscle. However, if there is a need for muscle growth, repair or regeneration, as in weight-bearing exercises or muscle trauma, SCs will be activated and enter the cell cycle. The activated satellite cells then proliferate by undergoing multiple cycles of divisions. Eventually, they differentiate either by fusing to pr-existing muscle fibers, where their nuclei become new myonuclei, or (albeit less commonly) by fusing together to give rise to new myofibers (Fig. 2)7. Nevertheless, quiescent satellite cell numbers in adult muscle remain relatively constant over several cycles of proliferation and differentiation, revealing an inherent self-renewal capacity8.
Satellite cells permit skeletal muscle growth and repair via two mechanisms, either by increasing the size of a pre-existing muscle fiber (hypertrophy) or by the formation of new muscle fibers (hyperplasia). Hypertrophy is considered the main mechanism by which growth in the growing adult skeletal muscle occurs, while the hyperplasia mechanism is described to be very limited. However, It must be noted that hyperplasia is the more potent mechanism of muscle growth, as it involves the addition of new muscle fibers, rather enlargement of an already pre-existing fibers.
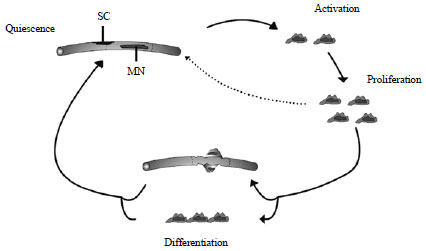 | |
| Fig. 2: | Life cycle of satellite cells (SCs). The SCs are usually quiescent and located beneath the fiber’s basal lamina. Under an appropriate stimulus, they become active and start to express the different myogenic factors. Their cytoplasmic volume increases significantly with well-developed organelles. After that, SCs start to proliferate and then differentiate by either fusing to a pre-existing fiber or by fusing together to form a new fiber. Eventually, SC nuclei become new myonuclei. However, a small proportion of proliferated SCs will not differentiate. Instead they return to quiescence to reestablish the pool of Scs. Source: Allouh7 |
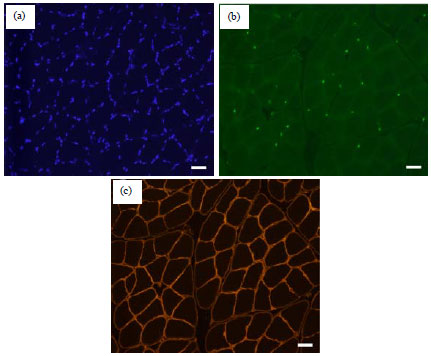 | |
| Fig. 3(a-c): | Immunofluorescence identification of satellite cell nuclei and myonuclei in cross sections from avian skeletal muscle, (a) All nuclei in blue stained with DAPI, (b) Satellite cell nuclei in green labeled by anti-Pax7 and (c) The muscle fiber basal laminae in red labeled by anti-laminin. Scale bar: 30 μm. Source: Allouh7 |
MARKERS OF SATELLITE CELLS
Many markers can be used to identify satellite cells, some of which are characteristically upregulated during different developmental stages of SC lifetime. One of the main markers of SCs is the paired box transcription factor 7 (Pax7). The Pax7 is a stable protein that belongs to the helix-turn-helix class of transcription factors. It was found to be expressed specifically in cells residing beneath the basal lamina in positions characteristic for SCs within the muscle (Fig. 3)9,10. It was shown that skeletal muscle from Pax7 knockout mice lacks SCs. In addition, a higher expression of Pax7 was evident during SC proliferation and this expression decreased as these cells differentiate8. The Pax7 is also expressed in the few cells that exit the cell cycle and revert back to quiescence representing a self-renewing pool of SCs8. The Pax7 is favorable to other SC markers since it is expressed by SCs in their different stages of development. As it is expressed during quiescence, activation and proliferation phase, but not by myonuclei7. In addition, effective antibodies against Pax7 are readily available7,8.
Another, well studied helix-turn-helix transcription factor is Pax3. This transcription factor has been implicated in the regulation of other SC markers, such as the receptor tyrosine kinase C-met. The Pax3 plays an important role in the initial stages of myogenesis. However, its expression is eventually down regulated once the cell is committed to myodifferentiation11.
Other markers include the myogenic regulatory factors, Myf5, MyoD and MRF4. The Myf5 which is co-expressed along with Pax7 in the activation and proliferation phases of the cells. The Myf5 is exclusively expressed in SCs that are committed to myogenic differentiation while the reservoir pool of stem cells are Myf5-ve. Another important marker for SC is the transcription factor MyoD, which is highly expressed during early activation of SCs. However, it is also expressed by differentiating myonuclei. The MRF4 expression, on the other hand, is augmented mainly during the differentiation stage of the cell12.
In addition, SCs express other markers that include, Barx2 transcription factor which is expressed with Pax7 and aid in the regulation of muscle growth and maintance13, cluster of differentiation protein (CD34) that is expressed in the quiescent cells; transmembrane heparan sulfate proteoglycanssyndecan-3 and syndecan-414, chemokine receptor CXCR415, caveolae-forming protein caveolin-116, calcitonin receptor17, vascular cell adhesion protein 1 VCAM-118, neural cell adhesion molecule 1 NCAM-119 and nuclear envelop proteins lamin A/C and emerin20. However, it must be noted that a lot of these markers are not exclusively expressed in SCs.
SATELLITE CELLS HETEROGENEITY
Due to the fact that most SCs express Pax7, they were used to be perceived as a homogenous group of cells. However, more recent studies suggest the presence of variable differentiation potential among SCs. It has been proposed that SCs are composed of two different populations: One population consists of SCs that possess a high myogenic differentiation potential, while the other population consist of a poorly differentiated subgroup of cells that are believed to be responsible for maintaining the stem cell pool21. Additionally, most of the SCs possess a high proliferative capacity, whereas the remaining few cells are characterized by their slow mitotic activity. These findings suggest that the minor slow dividing subpopulation is responsible for maintaining the quiescent stem cell pool, whereas the major highly proliferative cell population constitutes the cells that undergo myogenic differentiation into mature muscle fibers21.
Satellite cells also exhibit heterogeneity in their gene expression. Several studies reported that only a small subset expresses Pax3 within the adult muscular tissue11. These Pax3+ cells tend to be more concentrated in the diaphragm and the axial body musculature1. In addition, a subpopulation of SCs does not express the CD34, Myf5 and M-cadherin receptors22. A recent study reported the presence of a minor subpopulation of SCs that express the surface markers Sca1/ABCG2, which are not normally expressed by the majority of adult SCs23. This Sca1/ABCG2+ subpopulation is referred to as SC side population23. Doyle et al.24 demonstrated the regenerative capacity of these cells when transplanted into damaged muscle tissue. However, their contribution to muscle regeneration seems to be limited.
Although SCs are considered to be the main contributor in muscle regeneration, other studies have revealed that non-satellite cells can also play a role in muscle repair and growth. These cells include; bone marrow derived stem cells, mesoangioblasts, pericytes and CD133+ cells. During muscle regeneration, these cells are able to contribute in myotube formation as well as SCs reconstitution. These cells do not initially express Pax7. However, once they enter in the SC niche, they can fuse with other myogenic cells to form muscle fibers. In addition, these cells are able to differentiate into Pax7-expressing cells, indicating that non-SCs may also form muscle SCs. This phenomenon may also attribute to satellite cell heterogeneity25.
MOLECULAR AND CELLULAR PATHWAYS OF SATELLITE CELLS
The molecular mechanisms and signaling pathways required for SC activation are still not very well understood. Previous studies proposed that activation of SC starts by nitric oxide synthesis from L-arginine within the injured muscle fibers. Nitric oxide reacts with its cytoplasmic receptor, guanylylcyclase, leading to the formation of cyclic guanylyl monophosphate. Consequently, this leads to Hepatocyte Growth Factor (HGF) release from the extracellular domains of myofibers. Thereafter, HGF binds to its tyrosine kinase c-Met receptors found on the cell membrane of SCs26,27. The HGF binding to its receptor stimulates multiple mitogen-activated protein kinases (MAPK) and phosphatidyl inositol 3 kinase pathways (PI3k-Akt- mTORC1)26,27. These activated pathways induce the transcription and translation of myogenic regulatory factors as well as other transcriptional factors required for SC activation and proliferation. In addition to HGF, many other factors may contribute to the process of SC activation, such as fibroblast growth factor-2, insulin-like growth factor-1 (IGF-1) and endothelial growth factor28.
A major signaling pathway activated after injury to muscle fibers is the p38α/MAPK, which leads to increased expression of the basic helix-loop-helix (bHLH) transcription factor MyoD29. The MyoD and other myogenic bHLH proteins (Myf5 and MRF4) are considered master regulatory genes of skeletal muscle differentiation, as they directly regulate gene expression throughout the differentiation program. The MyoD action in SCs activation and differentiation is antagonized by another bHLH transcription factors called the Twist genes30,31.
Without injury, SCs maintain their quiescent state through activation of the notch signaling pathway that reversibly arrests their mitosis and decrease their metabolic activity32. The notch signaling pathway is extremely complex and is controlled by multiple regulators33. In humans, this pathway involves 4 receptors and five ligands33. Prior to SCs activation, this pathway must be inactivated in order for SCs to exit their quiescent state and achieve a state of alertness. Lin et al.34 reported that continuous notch signaling blockade will lead to muscular dystrophic phenotypes and impaired muscle regeneration due to exhaustion of SCs.
Once activated, SCs undergo asymmetric division, with one daughter cell committed to myogenis is while the other cell reverts back to quiescence to maintain a constant pool. The transcription factors MyoD, Myf5 and Myogenin have asymmetric expression within the daughter cells, as committed cells will upregulate the expression of MyoD and Myogenin while MyoD and myogenin negative cells will remain as reservoir35. It was also found that Myf5-ve cells contain an abundance of activated Notch receptors, suggesting a quiescent state while Myf5+ve cells receive the notch ligand Delta 1, which inhibits the notch signaling pathway and promote myogenesis12.
After an injury, the degree of muscle regeneration would not only depend on the potency of SCs but also on the surrounding composition of the extracellular matrix which harbors specific growth factors and signaling molecules7. The actions of these growth factors would be further regulated by the availability of appropriate primary and secondary SC receptors, including heparan sulfate proteoglycans (HSPGs). Many growth factors require HSPGs as co-receptors in addition to their specific primary receptors36. The HSPGs interact with growth factors and their primary receptors via their heparan sulfate chains. Due to that, the sulfation levels of HSPGs play a role in the regulation of some growth factors activity. A previous study revealed that HGF and fine-tuned sulfation levels are considered two major factors controlling SCs through regulating their relative activities of proliferation and differentiation37.
The different cellular components of the innate immune system are believed to play a role in muscle regeneration after injury. Their role is mediated by the secretion of a variety of cytokines and chemokines which regulate SC function38. The cytokine IL-6, secreted by neutrophils, acts as a chemotactic factor that attracts monocytes and macrophages allowing them to infiltrate the injured muscle tissue. It also induces the macrophages to produce granulocyte colony-stimulating factor which plays a role in myoblast proliferation and differentiation38.
Moreover, neutrophils produce tumor necrosis factor alpha (TNFα) which promotes the classical differentiation of monocyte into M1 macrophage (pro-inflammatory macrophage) which can be observed 24 h after injury. The M1 macrophages start to accumulate in the injured tissue reaching their peak concentration 2-3 days later39. Another form of macrophages, the M2 macrophages, peak 4-6 days after injury due to the release of IL-4 and IL-13 from the cellular components of the adaptive immune system40. These M2 macrophages secrete anti-inflammatory cytokines that are essential for tissue repair and regeneration41, such as IL-10. It is speculated that TNFα, secreted by neutrophils and M1 macrophages, stimulates SCs to enter the cell cycle and promotes their proliferation while suppressing their final stages of differentiation42. It also has been shown that TNFα acts as a chemotactic factor that attracts SC to the site of injury43. As SCs switch from proliferation to differentiation, the number of M1 macrophage decreases while the M2 macrophage population increases within the damaged tissue44. This shift increases the production of the anti-inflammatory cytokines IL-10 which enhances the proliferation and differentiation of SCs45. Furthermore, M2 macrophages produce the growth factor IGF-1 which plays a major role in muscle regeneration46,47.
SATELLITE CELL FREQUENCY AND DISTRIBUTION
The frequency of satellite cells is calculated by dividing the number of satellite cells nuclei by the sum of SC nuclei and myonuclei in the skeletal muscles. The SC frequency depends on several factors such as age, species, type of muscle fiber and health state of the muscle48. The number of SCs decreases with advancing age in rodents7. In addition, Allouh et al.49 showed that the frequency of SCs in female chickens decreased from about 20% at 9 days post-hatch to <5% at 115 days. However, the frequency was retained to approximalty 16% at the tapered ends of muscle fibers49. The progressive decline in SC number is presumably attributed to fusion of SCs into new or preexisting muscle fibers during postnatal muscle growth.
The frequency of satellite cells differs among different species. For example, the frequency of SCs in procrine sartorius muscle was found to be 0.01, while in frog sartorius muscle28 it was 0.10. The frequency of SCs also depends on the type of skeletal muscle fiber. There are more SCs in red slow twitch (oxidative) fibers than in white fast twitch (glycolytic) fibers. This is probably due to the higher demand for myonuclei in red than white fibers, since SC population in a muscle always reflects the requirement for myonuclei production. It is well established that red fibers possess more nuclei than white fibers of the same size7. This could be attributed to the higher metabolic activity and higher protein turnover in red than white fibers. In addition, SCs are more concentrated near specific anatomic structures, such as motor end plates, blood capillaries and at the ends of growing fibers48-50.
SATELLITE CELLS AND MUSCLE HYPERTROPHY
The exact role of SCs in muscle hypertrophy is still a matter of debate. Some studies proposed that SCs activation is essential in hypertrophy due to the fact that they represent the sole source for new myonuclei51. This necessity of SCs was further demonstrated in a recent study by Egner et al.52, who showed that hypertrophy was not possible in the SC depleted plantaris mouse muscle. However, other studies demonstrated that skeletal muscle can undergo hypertrophy without SCs by enhancing the transcription of muscle genes within the preexisting myonuclei. McCarthy et al.53 demonstrated that the overloaded mouse plantaris muscle was able to undergo normal hypertrophy even after SC depletion. In a similar experiment, Fry et al.54 also reported that overloaded muscles were able to undergo normal hypertrophy after SC depletion, but this hypertrophic growth was not sustainable and started to wane after 2 weeks of SCs ablation.
Other studies proposed that muscle hypertrophy occurs coordinately by both SC differentiation and increased transcriptional activity of preexisting myonuclei55. However, one of these mechanisms usually predominates the other depending on several factors such as, age, species and the nature of the growth stimulus55. More specifically, it is suggested that when the size of myofiber increases during muscle hypertrophy, the sarcoplasmic volume per myonucleus, known as the myonuclear domain (MND) will increase gradually before any increase in the number of myonuclei56,57. The increase in MND size reaches a maximum level designating that each myonucleus may also have reached the maximum transcriptional ability. Thereafter, new myonuclei are added from proliferating SCs to the hypertrophied fiber58.
Petrella et al.59 performed a cohort study of 66 human subjects to support the evidence that extreme muscle hypertrophy is facilitated by the continuous proliferation of SCs and their fusion to the preexisting muscle fibers. The findings suggested that the availability of SCs in the untrained muscles appear to be a very important determinant of hypertrophic potential. It is also showed that extreme hypertrophy was coupled with two-fold increase in the pool of SCs. It is proposed that once the initial myonuclear protein synthesis reaches its maximum threshold, it induces new SCs recruitment and proliferation for continuous growth59.
ANABOLIC ANDROGENIC STEROIDS AND SATELLITE CELLS
Anabolic androgenic steroids (AASs) are synthetic drugs that resemble the male endogenous steroid hormone, testosterone. Skeletal muscle is a primary target for these drugs where they exert their anabolic effects60-62. Several studies had demonstrated that administration of AASs leads to muscle hypertrophy associated with significant increases in all SC parameters, including SC frequency, concentration and number per mm of muscle fiber63,64. It is still unclear whether AASs increase SC numbers through enhancing their proliferation, reducing their apoptosis or both. In addition, administration of AASs leads to significant increases in the levels of Pax7 expression, number of myonuclei and DNA content in the skeletal muscle tissue55.
The exact mechanisms by which AASs activate SCs is still not very well understood. It is believed that the anabolic effect of AASs is mediated by their binding to the nuclear androgen receptors that are expressed by both myonuclei and SCs. Several studies showed that administration of AASs leads to upregulation in androgen receptors65. A second mechanism for AASs action includes raising the level of serum growth hormone which in turn increases serum IGF-1. The IGF-1 acts through 2 mechanisms to induce skeletal muscle growth. The first one, by enhancing translational ability within preexisting muscle nuclei, which increases protein synthesis within the muscle. The second one, by diminishing the inhibitory effect of p27 cyclin-dependent kinase protein on cyclin-dependent kinase 2, which is a necessary enzyme for activating SCs66. A third speculated mechanism implies that some AASs may act on the muscle indirectly by increasing the level of free endogenous testosterone. It is possible that certain AASs can compete with endogenous testosterone for binding to certain testosterone-binding proteins in the circulation. This will lead to an increase in serum concentration of free endogenous testosterone55.
SATELLITE CELLS IN MUSCULAR DYSTROPHY
One of the most widely accepted concepts regarding the role of SCs in Duchenne Muscular Dystrophy (DMD) is that they become exhausted by continuous cycles of muscle regeneration. Eventually, SCs lose their regenerative capacity and become unable to comply with the high regenerative demand by the muscle fibers. However, some studies reported that DMD patients have a significant increase in SC frequency. For example, Kottlors and Kirschner67 showed a significant expansion in SC population in DMD patients. In addition, Bankole et al.68 revealed a significant raise in SC numbers in type I oxidative fibers of patients with advanced stages of DMD.
Due to these inconsistent findings, several studies in the literature proposed an alternative mechanism explaining the loss of SC regenerative capacity in DMD patients. It is suggested that due to the dystrophin absence, SC will lose their polarity. This apolar state will abolish the SCs of their capability to undergo asymmetric mitosis and proper differentiation, resulting in a higher frequency of dividing but not appropriately differentiated SCs in DMD patients69-71.
THERAPEUTIC USE OF SATELLITE CELLS
The ability to isolate and transplant SCs could serve as the basis for translational efforts to regenerate human skeletal muscle. However, multiple obstacles stand in the way of effectively harnessing human endogenous SCs, such as the inability to expand the limited population of SCs ex vivo without losing their stem cell properties and the difficulty of developing proper models to test the harnessed cells in vivo. In the last five decades, several studies and clinical trials have been conducted to transplant myogenic stem cells into hosts suffering from muscle wasting diseases. Partridge et al.72 demonstrated the ability of transplanted myoblasts to fuse with the host myofibers. A decade later, it is demonstrated that injection of normal myoblasts into mdx mice led to formation of dystrophin expressing myofibers73. This opened the door for multiple subsequent clinical trials on human and animal subjects74,75. Meng et al.76 demonstrated that human CD133+ myoblast were able to generate functional SCs when xeno transplanted into immuno-deficient mice.
An alternative approach of transplantation is to implant entire donor myofibers with their residing SCs77. This method has the advantage of transplanting SCs with their surrounding niche. However, this method is still difficult to apply in clinical practice. In addition, recent experiments demonstrated the ability to isolate SCs using different techniques such as fluorescence-activated cell sorting (FACS). The isolated cells can then be cultured and transplanted into the host tissue. Nonetheless, the culturing procedure appears to cause reduction in the regenerative potential of the isolated SCs78. In a recent study by Xu et al.79, CD29+/56+ human SCs have been successfully identified and isolated from a diverse group of muscles in the head, trunk and limbs. These cells were then xenotransplanted into mice either by complete myofiber engraftment or as FACS isolated cells without their niche. In both techniques, the transplanted cells were able to produce a reasonably considerable amount of new human muscle fibers within the muscular tissue of the injured mice79.
The therapeutic potential of SCs transplantation was further demonstrated in a recent study by Abou-Khalil et al.80, who showed that SCs activation after injury may play an important role in bone healing after fracture. As activated SCs produce multiple bone morphogenic proteins and growth factors that are crucial for bone regeneration like IGF-1 and fibroblast growth factor 2. In addition, the activated SCs surrounding the fracture site can be integrated into the callus where they will differentiate into new chondrocytes80.
Despite the promising potential of SCs to be cultivated in cell therapy, many obstacles prevent their use from becoming a reality. These include the difficulty in providing the ideal culturing conditions, collection of the most fitting subpopulation of SCs and proper manipulation of the signaling pathways that control the SCs lifecycle. Moreover, clinicians must consider multiple factors when they decide whether to deliver the cells by local or systemic injections. These factors include the number of the affected muscles, the extent of damage and the degree of disease severity12. Due to the fact that each mode of delivery has its own advantages and disadvantages, combined internal and external injections of SCs may produce the most promising outcome12. However, more investigations are still needed in the future to explore the full therapeutic potential for SCs.
SUMMARY OF FINDINGS
| • | The SCs contribute in postnatal muscle growth and repair by proliferation and fusion to pre-existing muscle fibers where their nuclei differentiate into new muscle nuclei (myonuclei) and their cytoplasm contribute to the fiber sarcoplasm |
| • | The transcription factor Pax7 is considered to be the most reliable marker for SCs, as it is expressed by SCs through their different stages (quiescence, activation and proliferation) |
| • | The two most noticeable signaling pathways that control SC activity are the notch signaling pathway that maintains SC quiescence and the HGF activated P13k-Akt signaling pathway that leads to activation of SCs |
| • | The SC is a key player in skeletal muscle hypertrophy. However, whether hypertrophy can occur without the need of SCs is still a matter of debate which requires further investigations |
| • | Administration of AASs increases the number of SCs in skeletal muscles. This may represent one of the underlying mechanisms by which these drugs exert their anabolic effects on skeletal muscles |
| • | Cultivation and transplantation of SCs provides a potential therapeutic option for treating different muscle wasting disorders |
CONCLUSION
In conclusion, SCs are considered the key players in skeletal muscle growth, maintenance and repair. Since their discovery five decades ago, many of SC regulatory pathways have been reported. Despite these findings, many concepts pertaining to these cells are still not completely understood. More research is still needed to elucidate the main cellular signaling pathways that control SC fate and the influence of anabolic androgenic steroids on these cells. The better understanding of SCs at both cellular and subcellular levels will aid in developing beneficial cell therapy protocols that can be translated into the clinical field.
SIGNIFICANCE STATEMENT
In this study we thoroughly studied the most recent findings in literature regarding the role of SCs as adult myogenic stem cells necessary for skeletal muscle growth, maintenance and repair. We revised that what has hitherto been reported regarding SC biological markers and molecular pathways that regulate their activity and life cycle. In addition, we have reviewed, for the first time, what has been reported about the effects of anabolic androgenic steroids on the numbers and distribution of SCs. This study is original in providing a detailed discussion and summary on the most recent reports in literature regarding the clinical applications of satellite cells. It especially highlighted the immense therapeutic potential of SCs in muscle wasting diseases (e.g., duchene muscular dystrophy). The findings of this study are considered of great importance to researchers in the field of skeletal muscle regeneration, since it provides a comprehensive summary for all the biological aspects of SCs which are considered to be the main contributor in postnatal muscle regeneration.
ACKNOWLEDGMENT
The authors sincerely thank Dr. Mohammed Z. Allouh for his courtesy in providing the figures used in this study and for his valuable comments on a previous draft of this manuscript.
REFERENCES
- Yin, H., F. Price and M.A. Rudnicki, 2013. Satellite cells and the muscle stem cell niche. Physiol. Rev., 93: 23-67.
CrossRefDirect Link - Mauro, A., 1961. Satellite cell of skeletal muscle fibers. J. Biophys. Biochem. Cytol., 9: 493-495.
PubMedDirect Link - Katz, B., 1961. The terminations of the afferent nerve fibre in the muscle spindle of the frog. Philos. Trans. R. Soc. London B: Biol. Sci., 243: 221-240.
CrossRefDirect Link - Scharner, J. and P.S. Zammit, 2011. The muscle satellite cell at 50: The formative years. Skeletal Muscle, Vol. 1.
CrossRefDirect Link - Zammit, P.S., T.A. Partridge and Z. Yablonka-Reuveni, 2006. The skeletal muscle satellite cell: The stem cell that came in from the cold. J. Histochem. Cytochem., 54: 1177-1191.
CrossRefDirect Link - De Angelis, L., L. Berghella, M. Coletta, L. Lattanzi and M. Zanchi et al., 1999. Skeletal myogenic progenitors originating from embryonic dorsal aorta coexpress endothelial and myogenic markers and contribute to postnatal muscle growth and regeneration. J. Cell Biol., 147: 869-878.
CrossRefDirect Link - Collins, C.A., 2006. Satellite cell self-renewal. Curr. Opin. Pharmacol., 6: 301-306.
CrossRefDirect Link - Halevy, O., Y. Piestun, M.Z. Allouh, B.W.C. Rosser and Y. Rinkevich et al., 2004. Pattern of Pax7 expression during myogenesis in the posthatch chicken establishes a model for satellite cell differentiation and renewal. Dev. Dynam., 231: 489-502.
CrossRefDirect Link - Kirkpatrick, L.J., M.Z. Allouh, C.N. Nightingale, H.G. Devon, Z. Yablonka-Reuveni and B.W.C. Rosser, 2008. Pax7 shows higher satellite cell frequencies and concentrations within intrafusal fibers of muscle spindles. J. Histochem. Cytochem., 56: 831-840.
CrossRefDirect Link - Relaix, F., D. Montarras, S. Zaffran, B. Gayraud-Morel and D. Rocancourt et al., 2006. Pax3 and Pax7 have distinct and overlapping functions in adult muscle progenitor cells. J. Cell Biol., 172: 91-102.
CrossRefDirect Link - Almeida, C.F., S.A. Fernandes, A.F. Ribeiro Jr., O.K. Okamoto and M. Vainzof, 2016. Muscle satellite cells: Exploring the basic biology to rule them. Stem Cells Int.
CrossRefDirect Link - Meech, R., M. Gomez, C. Woolley, M. Barro and J.A. Hulin et al., 2010. The homeobox transcription factor barx2 regulates plasticity of young primary myofibers. PLoS ONE, Vol. 5.
CrossRefDirect Link - Cornelison, D.D.W., M.S. Filla, H.M. Stanley, A.C. Rapraeger and B.B. Olwin, 2001. Syndecan-3 and syndecan-4 specifically mark skeletal muscle satellite cells and are implicated in satellite cell maintenance and muscle regeneration. Dev. Biol., 239: 79-94.
CrossRefDirect Link - Ratajczak, M.Z., M. Majka, M. Kucia, J. Drukala, Z. Pietrzkowski, S. Peiper and A. Janowska-Wieczorek, 2003. Expression of functional CXCR4 by muscle satellite cells and secretion of SDF-1 by muscle-derived fibroblasts is associated with the presence of both muscle progenitors in bone marrow and hematopoietic stem/progenitor cells in muscles. Stem Cells, 21: 363-371.
CrossRefPubMedDirect Link - Volonte, D., Y. Liu and F. Galbiati, 2005. The modulation of caveolin-1 expression controls satellite cell activation during muscle repair. FASEB J., 19: 237-239.
CrossRefDirect Link - Fukada, S.I., A. Uezumi, M. Ikemoto, S. Masuda and M. Segawa et al., 2007. Molecular signature of quiescent satellite cells in adult skeletal muscle. Stem Cells, 25: 2448-2459.
CrossRefDirect Link - Rosen, G.D., J.R. Sanes, R. LaChance, J.M. Cunningham, J. Roman and D.C. Dean, 1992. Roles for the integrin VLA-4 and its counter receptor VCAM-1 in myogenesis. Cell, 69: 1107-1119.
CrossRefDirect Link - Dumont, N.A., Y.X. Wang and M.A. Rudnicki, 2015. Intrinsic and extrinsic mechanisms regulating satellite cell function. Development, 142: 1572-1581.
CrossRefDirect Link - Gnocchi, V.F., R.B. White, Y. Ono, J.A. Ellis and P.S. Zammit, 2009. Further characterisation of the molecular signature of quiescent and activated mouse muscle satellite cells. PLoS One, Vol. 4.
CrossRefDirect Link - Tierney, M.T. and A. Sacco, 2016. Satellite cell heterogeneity in skeletal muscle homeostasis. Trends Cell Biol., 26: 434-444.
CrossRefDirect Link - Beauchamp, J.R., L. Heslop, D.S.W. Yu, S. Tajbakhsh and R.G. Kelly et al., 2000. Expression of CD34 and Myf5 defines the majority of quiescent adult skeletal muscle satellite cells. J. Cell Biol., 151: 1221-1234.
CrossRefDirect Link - Tanaka, K.K., J.K. Hall, A.A. Troy, D.D.W. Cornelison, S.M. Majka and B.B. Olwin, 2009. Syndecan-4-expressing muscle progenitor cells in the sp engraft as satellite cells during muscle regeneration. Cell Stem Cell, 4: 217-225.
CrossRefDirect Link - Doyle, M.J., S. Zhou, K.K. Tanaka, A. Pisconti, N.H. Farina, B.P. Sorrentino and B.B. Olwin, 2011. Abcg2 labels multiple cell types in skeletal muscle and participates in muscle regeneration. J. Cell Biol., 195: 147-163.
CrossRefDirect Link - Motohashi, N. and A. Asakura, 2014. Muscle satellite cell heterogeneity and self-renewal. Front. Cell Dev. Biol., Vol. 2.
CrossRefDirect Link - Wozniak, A.C., J. Kong, E. Bock, O. Pilipowicz and J.E. Anderson, 2005. Signaling satellite-cell activation in skeletal muscle: Markers, models, stretch and potential alternate pathways. Muscle Nerve, 31: 283-300.
CrossRefPubMedDirect Link - Rodgers, J.T., K.Y. King, J.O. Brett, M.J. Cromie and G.W. Charville et al., 2014. mTORC1 controls the adaptive transition of quiescent stem cells from G0 to Galert. Nature, 510: 393-396.
CrossRefDirect Link - Hawke, T.J. and D.J. Garry, 2001. Myogenic satellite cells: Physiology to molecular biology. J. Applied Physiol., 91: 534-551.
Direct Link - Jones, N.C., K.J. Tyner, L. Nibarger, H.M. Stanley, D.D.W. Cornelison, Y.V. Fedorov and B.B. Olwin, 2005. The p38α/β mapk functions as a molecular switch to activate the quiescent satellite cell. J. Cell Biol., 169: 105-116.
CrossRefDirect Link - Franco, H.L., J. Casasnovas, J.R. Rodriguez-Medina and C.L. Cadilla, 2011. Redundant or separate entities?-Roles of Twist1 and Twist2 as molecular switches during gene transcription. Nucl. Acids Res., 39: 1177-1186.
CrossRefDirect Link - Coffman, J.A., S. Rieger, A.N. Rogers, D.L. Updike and V.P. Yin, 2016. Comparative biology of tissue repair, regeneration and aging. Regenerative Med., Vol. 1.
Direct Link - Kondoh, K., K. Sunadome and E. Nishida, 2007. Notch signaling suppresses p38 MAPK activity via induction of MKP-1 in myogenesis. J. Biol. Chem., 282: 3058-3065.
CrossRefDirect Link - Kaseb, H.O. and S.M. Gollin, 2015. Concerning consequences of blocking notch signaling in satellite muscle stem cells. Front. Cell Dev. Biol., Vol. 3.
CrossRefDirect Link - Lin, S., H. Shen, B. Jin, Y. Gu and Z. Chen et al., 2013. Brief report: Blockade of notch signaling in muscle stem cells causes muscular dystrophic phenotype and impaired muscle regeneration. Stem Cells, 31: 823-828.
CrossRefDirect Link - Yennek, S., M. Burute, M. Thery and S. Tajbakhsh, 2014. Cell adhesion geometry regulates non-random DNA segregation and asymmetric cell fates in mouse skeletal muscle stem cells. Cell Rep., 7: 961-970.
CrossRefDirect Link - Casar, J.C., C. Cabello-Verrugio, H. Olguin, R. Aldunate, N.C. Inestrosa and E. Brandan, 2004. Heparan sulfate proteoglycans are increased during skeletal muscle regeneration: Requirement of syndecan-3 for successful fiber formation. J. Cell Sci., 117: 73-84.
CrossRefDirect Link - Gill, R., L. Hitchins, F. Fletcher and G.K. Dhoot, 2010. Sulf1A and HGF regulate satellite-cell growth. J. Cell Sci., 123: 1873-1883.
CrossRefDirect Link - Zhang, C., Y. Li, Y. Wu, L. Wang, X. Wang and J. Du, 2013. Interleukin-6/Signal Transducer and Activator of Transcription 3 (STAT3) pathway is essential for macrophage infiltration and myoblast proliferation during muscle regeneration. J. Biol. Chem., 288: 1489-1499.
CrossRefDirect Link - Saclier, M., S. Cuvellier, M. Magnan, R. Mounier and B. Chazaud, 2013. Monocyte/macrophage interactions with myogenic precursor cells during skeletal muscle regeneration. FEBS J., 280: 4118-4130.
CrossRefDirect Link - Mantovani, A., A. Sica, S. Sozzani, P. Allavena, A. Vecchi and M. Locati, 2004. The chemokine system in diverse forms of macrophage activation and polarization. Trends Immunol., 25: 677-686.
CrossRefPubMedDirect Link - Arnold, L., A. Henry, F. Poron, Y. Baba-Amer and N. van Rooijen et al., 2007. Inflammatory monocytes recruited after skeletal muscle injury switch into antiinflammatory macrophages to support myogenesis. J. Exp. Med., 204: 1057-1069.
CrossRefDirect Link - Langen, R.C.J., J.L. van der Velden, A.M.W.J. Schols, M.C.J.M. Kelders, E.F.M. Wouters and Y.M.W. Janssen-Heininger, 2004. Tumor necrosis factor-alpha inhibits myogenic differentiation through myod protein destabilization. FASEB J., 18: 227-237.
CrossRefDirect Link - Torrente, Y., E. El Fahime, N.J. Caron, R. Del Bo and M. Belicchi et al., 2003. Tumor Necrosis Factor-α (TNF-α) stimulates chemotactic response in mouse myogenic cells. Cell Transplant., 12: 91-100.
CrossRefDirect Link - Saini, J., J.S. McPhee, S. Al-Dabbagh, C.E. Stewart and N. Al-Shanti, 2016. Regenerative function of immune system: Modulation of muscle stem cells. Ageing Res. Rev., 27: 67-76.
CrossRefDirect Link - Deng, B., M. Wehling-Henricks, S.A. Villalta, Y. Wang and J.G. Tidball, 2012. IL-10 triggers changes in macrophage phenotype that promote muscle growth and regeneration. J. Immunol., 189: 3669-3680.
CrossRefPubMedDirect Link - Tonkin, J., L. Temmerman, R.D. Sampson, E. Gallego-Colon and L. Barberi et al., 2015. Monocyte/macrophage-derived IGF-1 orchestrates murine skeletal muscle regeneration and modulates autocrine polarization. Mol. Therapy, 23: 1189-1200.
CrossRefDirect Link - Tidball, J.G. and S.S. Welc, 2015. Macrophage-derived IGF-1 is a potent coordinator of myogenesis and inflammation in regenerating muscle. Mol. Therapy, 23: 1134-1135.
CrossRefPubMedDirect Link - Allouh, M.Z. and B.W.C. Rosser, 2006. Greater frequency and concentration of satellite cells at the ends of developing skeletal muscle fibers. FASEB J., 20: A29-A29.
Direct Link - Allouh, M.Z., Z. Yablonka-Reuveni and B.W. Rosser, 2008. Pax7 reveals a greater frequency and concentration of satellite cells at the ends of growing skeletal muscle fibers. J. Histoch. Cytochem., 56: 77-87.
CrossRefPubMedDirect Link - Schultz, E. and K.M. McCormick, 1994. Skeletal muscle satellite cells. Rev. Physiol. Biochem. Pharmacol., 123: 213-257.
CrossRefDirect Link - O'Connor, R.S. and G.K. Pavlath, 2007. Point:Counterpoint: Satellite cell addition is/is not obligatory for skeletal muscle hypertrophy. J. Applied Physiol., 103: 1099-1100.
CrossRefDirect Link - Egner, I.M., J.C. Bruusgaard and K. Gundersen, 2016. Satellite cell depletion prevents fiber hypertrophy in skeletal muscle. Development, 143: 2898-2906.
CrossRefDirect Link - McCarthy, J.J., J. Mula, M. Miyazaki, R. Erfani and K. Garrison et al., 2011. Effective fiber hypertrophy in satellite cell-depleted skeletal muscle. Development, 138: 3657-3666.
CrossRefPubMedDirect Link - Fry, C.S., J.D. Lee, J.R. Jackson, T.J. Kirby and S.A. Stasko et al., 2014. Regulation of the muscle fiber microenvironment by activated satellite cells during hypertrophy. FASEB J., 28: 1654-1665.
CrossRefDirect Link - Allouh, M.Z. and M.H. Aldirawi, 2012. Influence of Mesterolone on satellite cell distribution and fiber morphology within maturing chicken pectoralis muscle. Anatom. Rec., 295: 792-799.
CrossRefDirect Link - Allouh, M.Z., 2011. Effect of Ferula hermonis root extract on rat skeletal muscle adaptation to exercise. Exp. Biol. Med., 236: 1373-1378.
CrossRefPubMedDirect Link - Rehfeldt, C., 2007. Satellite cell addition is/is not obligatory for skeletal muscle hypertrophy. J. Applied Physiol., 103: 1104-1106.
CrossRefDirect Link - Kadi, F., 2008. Cellular and molecular mechanisms responsible for the action of testosterone on human skeletal muscle. A basis for illegal performance enhancement. Br. J. Pharmacol., 154: 522-528.
CrossRefPubMedDirect Link - Petrella, J.K., J.S. Kim, D.L. Mayhew, J.M. Cross and M.M. Bamman, 2008. Potent myofiber hypertrophy during resistance training in humans is associated with satellite cell-mediated myonuclear addition: A cluster analysis. J. Applied Physiol., 104: 1736-1742.
CrossRefDirect Link - Allouh, M.Z. and B.W.C. Rosser, 2007. The effect of nandrolone decanoate on the frequency and concentration of satellite cells in skeletal muscle. FASEB J., 21: A203-A203.
Direct Link - Allouh, M.Z. and M.H. Aldirawi, 2012. Effects of mesterolone on satellite cell distribution and myonuclear number in maturing skeletal muscle fibers. FASEB J., 26: 914.5-914.5.
Direct Link - Allouh, M.Z. and M.H. Aldirawi, 2013. Sustanon administration induces satellite cell proliferation and giant fiber formation in growing skeletal muscle without exercise. FASEB J., 27: 524.2-524.2.
Direct Link - Allouh, M.Z. and M.H. Aldirawi, 2012. Effects of sustanon on the distribution of satellite cells and the morphology of skeletal muscle fibers during maturation. Pak. J. Biol. Sci., 15: 215-223.
CrossRefDirect Link - Allouh, M.Z. and B.W. Rosser, 2010. Nandrolone decanoate increases satellite cell numbers in the chicken pectoralis muscle. Histol. Histopathol., 25: 133-140.
PubMedDirect Link - Carson, J.A., W.J. Lee, J. McClung and G.A. Hand, 2002. Steroid receptor concentration in aged rat hindlimb muscle: Effect of anabolic steroid administration. J. Applied Physiol., 93: 242-250.
CrossRefDirect Link - Alen, M., P. Rahkila, M. Reinila and R. Vihko, 1987. Androgenic-anabolic steroid effects on serum thyroid, pituitary and steroid hormones in athletes. Am. J. Sports Med., 15: 357-361.
CrossRefDirect Link - Kottlors, M. and J. Kirschner, 2010. Elevated satellite cell number in duchenne muscular dystrophy. Cell Tissue Res., 340: 541-548.
CrossRefDirect Link - Bankole, L.C., L. Feasson, E. Ponsot and F. Kadi, 2013. Fibre type-specific satellite cell content in two models of muscle disease. Histopathology, 63: 826-832.
CrossRefDirect Link - Chang, N.C., F.P. Chevalier and M.A. Rudnicki, 2016. Satellite cells in muscular dystrophy-lost in polarity. Trends Mol. Med., 22: 479-496.
CrossRefDirect Link - Kharraz, Y., J. Guerra, P. Pessina, A.L. Serrano and P. Munoz-Canoves, 2014. Understanding the process of fibrosis in duchenne muscular dystrophy. Biomed. Res. Int.
CrossRefDirect Link - Dumont, N.A., Y.X. Wang, J. von Maltzahn, A. Pasut, C.F. Bentzinger, C.E. Brun and M.A. Rudnicki, 2015. Dystrophin expression in muscle stem cells regulates their polarity and asymmetric division. Nat. Med., 21: 1455-1463.
CrossRefDirect Link - Partridge, T.A., M. Grounds and J.C. Sloper, 1978. Evidence of fusion between host and donor myoblasts in skeletal muscle grafts. Nature, 273: 306-308.
CrossRefDirect Link - Partridge, T.A., J.E. Morgan, G.R. Coulton, E.P. Hoffman and L.M. Kunkel, 1989. Conversion of mdx myofibres from dystrophin-negative to -positive by injection of normal myoblasts. Nature, 337: 176-179.
CrossRefDirect Link - Mendell, J.R., J.T. Kissel, A.A. Amato, W. King and L. Signore et al., 1995. Myoblast transfer in the treatment of duchenne's muscular dystrophy. New Engl. J. Med., 333: 832-838.
CrossRefDirect Link - Morandi, L., P. Bernasconi, M. Gebbia, M. Mora, F. Crosti, R. Mantegazza and F. Cornelio, 1995. Lack of mRNA and dystrophin expression in DMD patients three months after myoblast transfer. Neuromusc. Disord., 5: 291-295.
CrossRefDirect Link - Meng, J., S. Chun, R. Asfahani, H. Lochmuller, F. Muntoni and J. Morgan, 2014. Human skeletal muscle-derived CD133+ cells form functional satellite cells after intramuscular transplantation in immunodeficient host mice. Mol. Therapy, 22: 1008-1017.
CrossRefPubMedDirect Link - Bentzinger, C.F., Y.X. Wang, J. von Maltzahn and M.A. Rudnicki, 2013. The emerging biology of muscle stem cells: Implications for cell-based therapies. BioEssays, 35: 231-241.
CrossRefDirect Link - Montarras, D., J. Morgan, C. Collins, F. Relaix and S. Zaffran et al., 2005. Direct isolation of satellite cells for skeletal muscle regeneration. Science, 309: 2064-2067.
CrossRefDirect Link - Xu, X., K.J. Wilschut, G. Kouklis, H. Tian and R. Hesse et al., 2015. Human satellite cell transplantation and regeneration from diverse skeletal muscles. Stem Cell Rep., 5: 419-434.
CrossRefDirect Link - Abou-Khalil, R., F. Yang, S. Lieu, A. Julien and J. Perry et al., 2015. Role of muscle stem cells during skeletal regeneration. Stem Cells, 33: 1501-1511.
CrossRefDirect Link