
Mehrnoosh Maalhagh
Student Research Committee, Jahrom University of Medical Sciences, Jahrom, Iran
Abdolreza Sotoodeh Jahromi
Research Center for non-Communicable Diseases, Jahrom University of Medical Sciences, Jahrom, Iran
Alireza Yusefi
Research Center for non-Communicable Diseases, Jahrom University of Medical Sciences, Jahrom, Iran
Ali Razeghi
Department of Pathology, Jahrom University of Medical Science, Jahrom, Iran
Hassan Zabetiyan
Research Center for non-Communicable Diseases, Jahrom University of Medical Sciences, Jahrom, Iran
Mohammad Yasin Karami
Department of Surgery, Shiraz University of Medical Science, Shiraz, Iran
Abdol Hossein Madani
Research Center for Social Determinants of Health Promotion, Hormozgan University of Medical Sciences, Bandarabbas, Iran
Pakistan Journal of Biological Sciences
Year: 2016 | Volume: 19 | Issue: 1 | Page No.: 43-48







ABSTRACT
Stress has inhibitory effect on HPG axis through increasing cortisol serum level. In this study, the effect of acute prepubertal stress on kisspeptin, which plays essential role in puberty achievement is assessed. To do this experimental study thirty immature healthy male wistar rats of 4 weeks old and without any symptoms of puberty were selected randomly. These rats were divided into three groups, randomly. Two groups were chosen as control and pretest and one as stress (test) group. Immobilization stress was applied for 10 days and serum level of cortisol, testosterone and kisspeptin were measured. Primary and secondary spermatocyte and sertoli cell evaluated and compared among groups. Mean serum level of kisspeptin in pretest group, control group and stress (test) group were 0.0381±0.0079, 91.0500±4.87430 and 15.2156±3.88135 pg mL–1 respectively. Serum level of kisspeptin had significant differences between three groups (p<0.001). Acute prepubertal immobilization stress led to decrease in serum level of kisspeptin and testosterone in stress (test) group compared to control groups. Also stress caused a significant decrease in the numbers of secondary spermatocytes of the test group.
PDF Abstract XML References Citation
Received: June 12, 2015;
Accepted: October 16, 2015;
Published: December 15, 2015
How to cite this article
Mehrnoosh Maalhagh, Abdolreza Sotoodeh Jahromi, Alireza Yusefi, Ali Razeghi, Hassan Zabetiyan, Mohammad Yasin Karami and Abdol Hossein Madani, 2016. Effects of Prepubertal Acute Immobilization Stress on Serum Kisspeptin Level and Testis Histology in Rats. Pakistan Journal of Biological Sciences, 19: 43-48.
DOI: 10.3923/pjbs.2016.43.48
URL: https://scialert.net/abstract/?doi=pjbs.2016.43.48
DOI: 10.3923/pjbs.2016.43.48
URL: https://scialert.net/abstract/?doi=pjbs.2016.43.48
INTRODUCTION
The kisspeptins (KP) encoded by the KISS1 gene are located on human chromosome 1q32 (Navarro et al., 2007; Mogford et al., 2014). Kisspeptins are a group of newly discovered peptides secreted by hypothalamic nuclei anteroventral periventricular nucleus (AVPV) a major product of KISS1 is a 54-amino acid peptide called kisspeptin-54, although there are other kinds of kisspeptins, which are the same in c-terminal area and act the same in stimulating their receptor called GPR5 (Navarro et al., 2007; Morelli et al., 2008).
After clarifying the role of kisspeptins in reproduction, other roles including essential regulatory function in gonadotropins secretion, the initiation of human reproduction and maintenance of sexual maturity has been mentioned (Morelli et al., 2008; Kanasaki et al., 2013). Further, it has been discovered that KP is secreted from other organs such as, placenta and other parts of the brain (Majzoub and Karalis, 1999).
Puberty is the period of life that is led to adulthood through dramatic physiologic and psychological changes (Bianco, 2012). Puberty achievement depends on several processes to activate hormonal axis which are necessary for starting puberty and maturation of reproductive function (Rhie, 2013). Factors such as genetic, nutritional state or environmental conditions can affect the onset of puberty and its subsequent events (Blum et al., 1997; Skorupskaite et al., 2014).
The KP gene mutations led to the loss of functions or decrease in the level of kisspeptin or its receptor have been shown to cause pubertal failure, abnormal gonadal function and impaired reproduction such as that found in patients with idiopathic hypogonadotropic hypogonadism (Zhang et al., 2013).
The KP regulates the secretion of gonadotropins by stimulation of upper pathway in Hypophysis Pituitary Gonadal (HPG) axis (Jameel et al., 2014). It is obvious that secreting hypothalamic gonadotropins realizing hormone (GnRH) is essential and required for the maturation of HPG axis and puberty switching. It has been mentioned that KP plays a primordial role in the onset of puberty by the stimulation of GnRHsecretion (Barnea and Tal, 1991; Kinsey Jones et al., 2009; Rhie, 2013; Mogford et al., 2014).
The KISS-1 is an extraordinary potent elicitor of LH and FSH release which acts on GnRH neurons (Rhie, 2013). Also increasing the cortisol serum level, it provides inhibitory effect on HPG axis (Rao et al., 2011). Chronic or acute stress decrease serum level of testosterone in male rats and estradiol in female rats (Iwasa et al., 2009).
There are several studies done in this field to prove the essential role of KP in puberty process. In previous study KP has been introduced as putative conduit for the metabolic gating of reproduction (Iwasa et al., 2009).
In another study, Pineda et al. (2010) mentioned the importance of KP in reproduction axis and the therapeutic role of agonist and antagonist of these peptides. Kinsey Jones et al. (2009) study revealed that pivotal role of KP in releasing hormones in HPG axis. Also reduction in expression of KISS-1 gene can decrease the secretion of Luteinizing Hormone (LH).
Prepubertal stress affects the puberty process and maturation of reproductive organs. Therefore in adults with delayed puberty, one of the differential diagnoses may be prepubertal stressor (Barnea and Tal, 1991; Kinsey Jones et al., 2009).
Considering all the studies mentioned above, there is a promising role for KP in puberty process. Therefore, in present study we decided to evaluate the effect of acute prepubertal immobilization stress on serum level of KP secretion and puberty conduction compared with control group.
MATERIALS AND METHODS
Experimental study: Thirty Immature and healthy Wistar male rats (100-150 g) of 4 weeks of age (prepubertal state) were used to apply in this research work. The animals were housed in standard environmental conditions of temperature (22±20°C), humidity (55±15%) and a 12 h light/dark cycle for at least 7 days for adaptation with research environment (Jahrom University of Medical Sciences Animal Laboratory, Iran). During the time of experiment, rats received the standard pellet diet and they had easy access to food and water. Protocol of the study was approved by the Medical Research Ethics Committee of Jahrom University of Medical Sciences.
According to the study design, sample size was determined based on Kohen chart. Thirty Rats were selected randomly and were divided into three equal groups (ten rats in each group). Being assured that all rats in the study were not mature, those of the first group were killed and their both testis were evicted as a pretest group and sent for histopathological studies. Blood samples were taken from the tail vein to measure cortisol, testosterone and kisspeptin1 (KP1).
The first group’s (pretest or immature rats) blood samples and histopathological data were used to be compared with those of mature rats (stress and control groups) histopathological and blood samples data at the end of study.
The second group or stress (test) group lived with immobilization stress. The third group or control (stress free) group living in the same condition were compared to those of the test group but without stress.
At the 8th day of the study, immobilization stress applied for the test group using restrainer for 10 days and each time for 2 h in the morning from 8-10 AM. After applying immobilization stress for 10 days, both groups (test and control) were kept for 7 days in the same conditions as already mentioned above. When all rats were almost 7 weeks old (51 days old) that was equivalent to puberty, the secretion of sexual hormones and maturation of HPG axis, both groups were sampled for the measurement of testosterone, cortisol and KP1 serum level (Castellano et al., 2010), the rats were then killed and their both testis were evicted and sent for pathological studies.
Laboratory and histopathological study: All of the blood samplings were done between 8-10 AM (Almeida et al., 1998). Samples taken from the three groups were centrifuged at 3000 rpm for 15 min and isolated sera were stored at -20°C until test.
Enzyme Linked Immune Sorbent Assay (ELISA) method was used by commercial kits for determining serum levels of testosterone (Diagnostics Biochem Canada Inc., Ontario, Canada), cortisol (My Bio Source, San Diego, CA 92195-3308 USA ) and KP1 (CUSABIO, Code No: CSB-E13434r).
Testis evicted and fixed with formaldehyde 10%, 5 micron sections were stained by hematoxylin and eosin staining method for evaluation and comparison of the effect of prepubertal stress on testis (primary and secondary spermatocyte and sertoli cell in the pretest, test and the control groups). The study was blinded and the histologist began to perform the histopathological review.
Statistical analysis: The results were expressed as mean±standard deviation (SD). Differences were considered statistically significant in case of p value less than 0.05. Data collected were analyzed by statistical software SPSS 16.0 for windows.
RESULTS
The reliability of acute immobilization stress that applied for test group confirmed with serum level of cortisol that measured in all three groups and showed significant rising in test group compared with pretest and control groups that lived without stress (p<0.001) (Fig. 1).
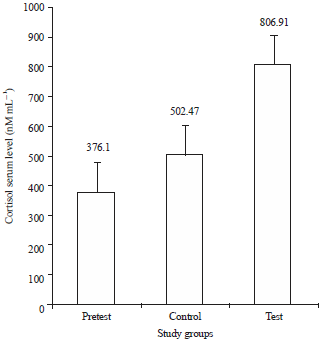 | |
| Fig. 1: | Serum level of cortisol in the three study groups |
Cortisol serum level of Pretest, Control and Test groups were 376.09±9.56, 502.46±35.18 and 806.91±24.59 nM mL–1, respectively. Serum level of testosterone and KP1 had significant differences in three groups as showed in Table 1 (p<0.001). Serum level of KP1, testosterone and cortisol had significant different among three groups compared together (pretest/test, control/test, control/pretest).
Serum level of testosterone in control group was more than test group (222.59±10.21 vs. 27.66±9.83 pg mL–1) also serum level of KP1 in test group showed the same results (90.60±4.882 vs. 15.50±3.774 pg mL–1). A negative effect of cortisol on serum level of testosterone was seen by comparing the serum level of cortisol in test and control groups (806.91±24.59 vs. 502.46±35.18 nM mL–1) with the serum level of testosterone in these two groups (27.66±9.83 vs. 222.59±10.21 pg mL–1).
Histological study: Testis cross sections in each three groups compared based on the number of primary and secondary spermatocytes. sertoli cell count in each section only reported to confirm the same size in different sections (p = 0.280).
The mean of primary spermatocyte count in pretest group was less than that in control and test groups and had significant difference (p<0.001). The mean of primary spermatocyte count in control and test groups had significant difference (p<0.001).
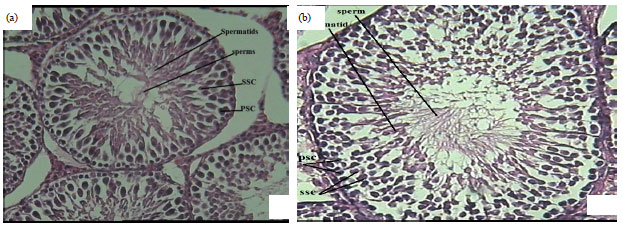 | |
| Fig. 2(a-b): | Cross sections of rat testes stained with H and E, magnification: (40×), (a) Test group and (b) Control group |
| Table 1: | Serum level of testosterone, KP1 and cortisol among three study groups |
 | |
| Table 2: | Primary and secondary spermatocytes and sertoli cell count among three study groups |
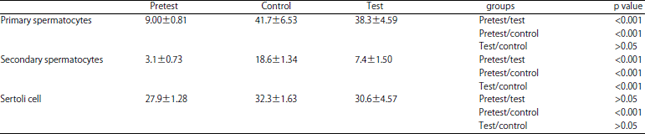 | |
The mean of secondary spermatocyte count was reduced in test group compared with control group and had significant difference (p<0.001) (Table 2 and Fig. 2).
DISCUSSION
The neuroendocrine response to stress played an important role in puberty; the author showed for the first time that acute immobilization stress played a role in the decrease of serum KP1 and testosterone concentrations and also reduce secondary spermatocyte. Acute immobilization stress during prepubertal phase in immature male rats decreased the serum level of kisspeptins. Rhie (2013) introduced KP as essential hormone in releasing GnRH hormone In the present study, the serum level of KP1 was correlated to that of testosterone in control group. Other studies showed mRNA expression of testosterone had negative feedback on KP mRNA expression which revealed the stimulatory effect of KP on HPG axis (Maric et al., 1996; Sawyer et al., 2011). These findings corresponded with the results of our study, while in the present study, only the correlation of serum level of testosterone and KP were determined. The negative and positive feedbacks in HPG axis couldn’t be proved due to the lack of measurements of other hormones of axis. In the present study, reliability of applying immobilization stress confirmed with measuring serum level of cortisol. According to previous studies, increasing in the cortisol serum level determined the valid immobilization stress, i.e., parallel to the results of previous studies (Huhman et al., 1991; Almeida et al., 1998; Majzoub and Karalis, 1999).
The present study showed that subsequent to increase in the cortisol serum level, sexual hormones of HPG axis (testosterone) decreased due to inhibitory effect of cortisol. These results were similar to those of previous studies (Huhman et al., 1991; Sawyer et al., 2011; Mogford et al., 2014). In the present study, the serum level of testosterone decreased in stress group compared to the control group (which had no stressor and had normal cortisol level).
Applying immobilization stress decreased testosterone and KP1 serum level and also there was no significant correlation between serum levels of testosterone and KP1, although this correlation was obvious in control group without any intervention. But reduction in the serum level of KP1 had not significant correlation with the decreased serum level of testosterone in test group compared to the control group. This result indicated that reducing affect of acute stress on serum level of testosterone was not related to the serum level of KP1.
Rao et al. (2011) study showed the inhibitory effect of inflammation as an environmental stress that was inducted by the injection of lipopolysaccharide on the expression of KP mRNA (Robb et al., 1978; Shibata et al., 2007) and another study showed that inflammation would decrease the sensitivity of tissues to the effects of KP (Smith et al., 2006).
Maturation of reproductive organs was dependent to the activity of HPG axis and adequate secretion of sexual hormones such as KP (first hormone of HPG axis) and testosterone (last hormone of HPG axis). In present study, the effect of KP and testosterone on maturation of gonadal tissue was interrupted by applying immobilization stress. This effect has been proved in previous studies that the chronic intermittent stress-induced alterations in the spermatogenesis (Nirupama et al., 2013; Skorupskaite et al., 2014), agreed the results of other studies. Metabolic state, stress and other neuropeptides such as, neurokinin B (NKB) had effects on KP and its stimulatory function (Aziz et al., 2013). In present study, the serum level of KP1 decreased by applying immobilization stress in prepubertal phase. Rats in test group had significant decreased in secondary spermatocyte. The difference between our study and previous studies was that in the present study, we applied prepubertal acute immobilization stress despite other studies which applied chronic immobilization stress or other kinds of environmental stress.
CONCLUSION
In conclusion, the present study demonstrated that the KP1 had a role in immobilization stress-induced suppression of the HPG axis. This was demonstrated by reversing the decrease in the serum testosterone level and KP1 serum level of test group. These data were the first to identify KP1 level, testosterone and secondary spermatocyte suppression by acute immobilization stress.
ACKNOWLEDGMENT
We would like to express our sincere gratitude to Jahrom University of Medical Sciences for financing this study.
REFERENCES
- Almeida, S.A., J.A. Anselmo-Franci, A.A. Rosa-e-Silva and T.L.L. Carvalho, 1998. Chronic intermittent immobilization of male rats throughout sexual development: A stress protocol. Exp. Physiol., 83: 701-704.
Direct Link - Aziz, N.M., M.M. Ragy and M.F. Gayyed, 2013. Effect of acute immobilization stress with or without a heme oxygenase inducer on testicular structure and function in male albino rats. J. Basic Clin. Physiol. Pharmacol., 24: 255-262.
CrossRefDirect Link - Barnea, E.R. and J. Tal, 1991. Stress-related reproductive failure. J. Assisted Reprod. Genet., 8: 15-23.
CrossRefDirect Link - Bianco, S.D.C., 2012. A potential mechanism for the sexual dimorphism in the onset of puberty and incidence of idiopathic central precocious puberty in children: Sex-specific kisspeptin as an integrator of puberty signals. Frontiers Endocrinol., Vol. 3.
CrossRef - Blum, W.F., P. Englaro, S. Hanitsch, A. Juul and N.T. Hertel et al., 1997. Plasma leptin levels in healthy children and adolescents: Dependence on body mass index, body fat mass, gender, pubertal stage and testosterone. J. Clin. Endocrin. Metab., 82: 2904-29010.
CrossRefDirect Link - Castellano, J.M., A.H. Bentsen, M. Romero, R. Pineda and F. Ruiz-Pino et al., 2010. Acute inflammation reduces kisspeptin immunoreactivity at the arcuate nucleus and decreases responsiveness to kisspeptin independently of its anorectic effects. Am. J. Physiol. Endocrinol. Metab., 299: E54-E61.
CrossRefDirect Link - Huhman, K.L., T.O. Moore, C.F. Ferris, E.H. Mougey and J.L. Meyerhoff, 1991. Acute and repeated exposure to social conflict in male golden hamsters: increases in plasma POMC-peptides and cortisol and decreases in plasma testosterone. Hormones Behav., 25: 206-216.
CrossRefDirect Link - Iwasa, T., T. Matsuzaki, M. Murakami, R. Kinouchi and R. Ogata et al., 2009. Neonatal lipopolysaccharide exposure attenuates the homotypic stress-induced suppression of LH secretion in adulthood in male rat. Int. J. Dev. Neurosci., 27: 345-349.
CrossRefDirect Link - Jameel, M.K., A.R. Joshi, J. Dawane, M. Padwal, A.R. Joshi, V.A. Pandit and R.R. Melinkeri, 2014. Effect of various physical stress models on serum cortisol level in Wistar rats. J. Clin. Diagnostic Res., 8: 181-183.
CrossRefDirect Link - Kanasaki, H., I.N. Purwana, A. Oride, T. Mijiddorj, U. Sukhbaatar and K. Miyazaki, 2013. Circulating kisspeptin and Pituitary Adenylate Cyclase-Activating Polypeptide (PACAP) do not correlate with gonadotropin serum levels. Gynecol. Endocrinol., 29: 583-587.
CrossRefDirect Link - Kinsey‐Jones, J.S., X.F. Li, A.M.I. Knox, E.S. Wilkinson and X.L. Zhu et al., 2009. Down-regulation of hypothalamic Kisspeptin and its receptor, Kiss1r, mRNA expression is associated with stress-induced suppression of luteinising hormone secretion in the female rat. J. Neuroendocrinol., 21: 20-29.
CrossRefDirect Link - Majzoub, J.A. and K.P. Karalis, 1999. Placental corticotropin-releasing hormone: Function and regulation. Am. J. Obstetrics Gynecol., 180: S242-S246.
CrossRefDirect Link - Maric, D., T. Kostic and R. Kovacevic, 1996. Effects of acute and chronic immobilization stress on rat Leydig cell steroidogenesis. J. Steroid Biochem. Mol. Biol., 58: 351-355.
CrossRefDirect Link - Mogford, J.T., C.N. Jayasena and W.S. Dhillo, 2014. Evaluating the potential utility of kisspeptin to treat reproductive disorders. Expert Rev. Endocrinol. Metab., 9: 251-261.
CrossRefDirect Link - Morelli, A., M. Marini, R. Mancina, M. Luconi and L. Vignozzi et al., 2008. Sex steroids and leptin regulate the "First Kiss" (KiSS 1/G-protein-coupled receptor 54 system) in human gonadotropin-releasing-hormone-secreting neuroblasts. J. Sexual Med., 5: 1097-1113.
CrossRefDirect Link - Navarro, V.M., J.M. Castellano, D. Garcia-Galiano and M. Tena-Sempere, 2007. Neuroendocrine factors in the initiation of puberty: The emergent role of kisspeptin. Rev. Endocrine Metab. Disorders, 8: 11-20.
CrossRefDirect Link - Nirupama, M., M. Devaki, R. Nirupama and H.N. Yajurvedi, 2013. Chronic intermittent stress-induced alterations in the spermatogenesis and antioxidant status of the testis are irreversible in albino rat. J. Physiol. Biochem., 69: 59-68.
CrossRefDirect Link - Pineda, R., D. Garcia-Galiano, A. Roseweir, M. Romero and M.A. Sanchez-Garrido et al., 2010. Critical roles of kisspeptins in female puberty and preovulatory gonadotropin surges as revealed by a novel antagonist. Endocrinology, 151: 722-730.
CrossRefPubMedDirect Link - Rao, Y.S., N.N. Mott and T.R. Pak, 2011. Effects of kisspeptin on parameters of the HPA axis. Endocrine, 39: 220-228.
CrossRefPubMedDirect Link - Rhie, Y.J., 2013. Kisspeptin/G protein-coupled receptor-54 system as an essential gatekeeper of pubertal development. Ann. Pediatr. Endocrinol. Metab., 18: 55-59.
CrossRefPubMedDirect Link - Robb, G.W., R.P. Amann and G.J. Killian, 1978. Daily sperm production and epididymal sperm reserves of pubertal and adult rats. J. Reprod. Fertil., 54: 103-107.
CrossRefDirect Link - Sawyer, I., S.J. Smillie, J.V. Bodkin, E. Fernandes, K.T. O'Byrne and S.D. Brain, 2011. The vasoactive potential of kisspeptin-10 in the peripheral vasculature. PLoS One.
CrossRefDirect Link - Shibata, M., R.L. Friedman, S. Ramaswamy and T.M. Plant, 2007. Evidence that down regulation of hypothalamic KiSS-1 expression is involved in the negative feedback action of testosterone to regulate luteinising hormone secretion in the adult male rhesus monkey (Macaca mulatta). J. Neuroendocrinol., 19: 432-438.
CrossRefPubMedDirect Link - Skorupskaite, K., J.T. George and R.A. Anderson, 2014. The kisspeptin-GnRH pathway in human reproductive health and disease. Hum. Reprod. Update, 20: 485-500.
CrossRefPubMedDirect Link - Smith, J.T., D.K. Clifton and R.A. Steiner, 2006. Regulation of the neuroendocrine reproductive axis by kisspeptin-GPR54 signaling. Reproduction, 131: 623-630.
CrossRefPubMedDirect Link - Zhang, M.J., Z.L. Gong, D. Han, X. Gao, Q. Tan and Y. Xu, 2013. Gene polymorphism and precocious puberty: A meta-analysis of case-control studies. Adv. Mater. Res., 647: 466-470.
CrossRefDirect Link