
S. M. Nasr
Department of Animal Husbandry and Animal Wealth Development, Faculty of Veterinary Medicine, Damanhour University, Egypt
LiveDNA: 20.12509
A. I. Ateya
Department of Animal Husbandry and Animal Wealth Development, Faculty of Veterinary Medicine, Mansoura University, Egypt
LiveDNA: 20.16146
K. M. Sadek
Department of Biochemistry, Faculty of Veterinary Medicine, Damanhour University, Egypt
LiveDNA: 20.15058
H. A. Radwan
Department of Animal Husbandry and Animal Wealth Development, Faculty of Veterinary Medicine, Mansoura University, Egypt
Pakistan Journal of Biological Sciences
Year: 2016 | Volume: 19 | Issue: 8-9 | Page No.: 338-344







ABSTRACT
Objective: The GH and MSTN gene polymorphisms and their association with body weight were declared in a population of 100 Friesian bull calves. Materials and Methods: For DNA extraction, collection of blood samples was carried out from the studied animals. The PCR for GH and MSTN genes yielded fragments of 329 and 1346 bp, respectively. Results: The PCR-HpaII digestion of 329 bp of GH gene revealed three genotypes: AA genotype possess undigested fragment (329 bp), AB genotype has three fragments (329, 224 and 105 bp) and BB genotype has two fragments (224 and 105 bp). The GH genotypes incidence and alleles frequency were calculated. For the 100 Friesian bull calves, genotypic frequencies for the AA, AB and BB genotypes were 0.1, 0.78 and 0.12, respectively and the allele frequencies for A and B allele frequencies were 0.49 and 0.51. Statistical analysis revealed that there was a significant effect of GH genotypes on body weight. The AB genotype possessed higher body weight than the other 2 genotypes. Regarding MSTN gene, PCR-DraI digestion of 1346 bp fragment was monomorphic; where it yielded four fragments (505, 427, 321 and 93 bp) in all animals under study. Conclusion: The outcome of this study is that it highlights the effectiveness of GH/HpaII locus as candidate marker for body weight in cattle rather than MSTN/DraI.
PDF Abstract XML References Citation
Received: July 08, 2016;
Accepted: August 01, 2016;
Published: August 15, 2016
How to cite this article
S. M. Nasr, A. I. Ateya, K. M. Sadek and H. A. Radwan, 2016. Effect of Genetic Polymorphisms in GH/HpaII and MSTN/DraI Loci on Body Weight in Friesian Bull Calves. Pakistan Journal of Biological Sciences, 19: 338-344.
DOI: 10.3923/pjbs.2016.338.344
URL: https://scialert.net/abstract/?doi=pjbs.2016.338.344
DOI: 10.3923/pjbs.2016.338.344
URL: https://scialert.net/abstract/?doi=pjbs.2016.338.344
INTRODUCTION
Growth traits of animals are of principle concern for breeder due to their determinant and pivotal economical value1. Development of molecular techniques makes scientists able to attain more precise and proficient selection objective by marker-assisted selection (MAS). The association of candidate gene polymorphism with growth traits uses MAS to predict such traits and makes it a candidate for use in breeding programs. It could also be used as an implement for early selection of high growth rate animals and to prevent economic losses that are afforded by wasteful breeding of undesirable animals2. Candidate gene discipline permits discovering variation among individuals in the population for definite region of the DNA and accelerates rate of response to selection for trait of interest (e.g., body weight)3,4.
Bovine growth hormone consists of from 190-191 amino acids and has 22 kD molecular weight with 1800-bp length5-7, 5 exons and 4 introns8,9. Using restriction fragment length polymorphism (RFLP) genetic marker, many variant regions have been stated in different sites of bGH gene10. The most important reported polymorphisms are mutations at the third intron (transition T to C) and the fifth exon (transversion C to G) which are identified by MspI and Alul restriction enzymes, respectively11-13. The GH has anabolic role, major regulator of postnatal growth and development, tissue growth as well as lipid, protein and carbohydrate metabolism14. The GH role has been detected in numerous tissues, including bone, adipose tissue and muscle, so GH can be used as a candidate gene for marker-assisted selection for growth traits in several livestock species, including cattle15. Many studies indicated the association between GH/HpaII locus and growth traits16-19. However, most of that reported analysis revealed opposing results or fail to demonstrate relationship.
Myostatin (MSTN) is one of the transforming growth factor β (TGF-β) superfamily, which comprises proteins that are key events in cell growth and development. The MSTN represents a negative myogenesis regulator20 by obstructing the MyoD and Myo5 factors, which are associated with the mechanisms of precursor cells differentiation into myoblasts21. The MSTN gene knockout is the principle cause of double muscling phenotype that refers to increase mass of skeletal muscle that occurs due to the muscle fibers hypertrophy or hyperplasia22-25. Bovine MSTN gene is located on chromosome 2 (BTA2) at 3.1 cM (centimorgan) from the centromeric region, next to microsatellite TGLA4426. It consists of 2 introns and 3 exons, with 1840 and 2033 nucleotides in the two intron and 373, 374 and 381 nucleotides in the three exons27. The mRNA codes for a protein with 375 amino acids28. MSTN-DraI locus polymorphism relatedness with body weight is scare29. Therefore, the main objectives of this study were to investigate the effect of genetic polymorphisms in GH/HpaII and MSTN/DraI loci on Friesian bull calves body weight.
MATERIALS AND METHODS
Animals and experimental samples collection: In this study, 100 Friesian bull calves were used. The farm records revealed that; the weaning weight of the studied animals was 90 kg b.wt., the birth weight was 29-38 kg and the weaning age was 75-100 days. Disodium EDTA containing tubes were used to prevent coagulation of blood during collection of samples. Then, blood samples storage was carried out at -20°C till DNA extraction procedures. This study practice was established via the animal welfare and ethics committee and the faculty of veterinary medicine, Damanhour University.
DNA extraction: The genomic DNA extraction was done via DNA purification kit procedures (Jena Bioscience, Germany). On 1.5% agarose gel, DNA quality was evaluated then examined in the UV transilluminator and bands were visualized and photographed. Moreover, to get high yield, enough concentration and purity of DNA, quantification of the extracted DNA was done using using nanodrop (UV-Vis spectrophotometer Q5000/USA).
PCR amplification: Amplification of fragments of GH gene and MSTN gene was carried out with expected amplicon sizes of 329 and 1346 bp7,29 respectively. The primer sequences are represented in Table 1.
The PCR was done in a reaction volume of 25 μL according with some modifications. The reaction consists of 2.5 μL of 10x Green buffer of Dream Taq (Thermo Scientific, Germany), 1 μL primer each primer forward and reverse (10 pmol), 0.5 μL of (10 mM) dNTP (Thermo Scientific, Germany), 0.5 μL Taq DNA polymerase (Thermo Scientific, Germany) and 17.5 μL ddH2O which finally added to 2 μL genomic DNA (20 ng μL–1).
| Table 1: | Primer sequences and size of the amplified fragments |
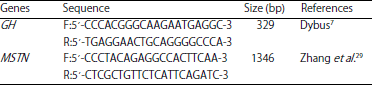 | |
The reactions were done in a thermal cycler (Sure cycler 8800, Malaysia) and thermal cycling program denaturizing at 94°C for 5 min, followed by 35 cycles at 94°C for 1min, annealing temperature 63°C for 1 min and extension at 72°C for 1 min, final step is the extension at 72°C for 10 min.
The PCR product of each sample (5 μL) and 100 bp DNA ladder (Thermo Scientific, Germany) were loaded in 2% (w/v) agarose gels in tris-borate-EDTA (TBE) buffer staining using ethidium bromide. The electrophoresis was carried out for 45 min at 100 V. The electrophoresis gel was examined on an UV transilluminator and bands were visualized and photographed.
Polymorphism detection: The PCR products of GH gene were cleaved by fast digest HpaII (Thermo Scientific, #FD0514) (isomer of MspI) at 37°C for 5 min; while, MSTN gene amplified fragments were digested with DraI (Thermo Scientific, #ER0221) at 37°C overnight. The reaction volume was 30 μL consisted of 10 μL PCR product, 17 μL H2o (dd water), 2 μL 10x buffer, 1 μL restriction enzyme. The polymorphism of the cleaved fragments recognition was carried out by agarose gel electrophoresis then their polymorphic pattern was obviously envisioned under U.V by gel documentation system.
Data analysis
Non-genetic factors adjustment: Based on established farm records, adjustment of the different recorded calf weaning body weight to 205 days of age body weight was done by deduction from birth weight, weaning weight and age using this equation30:
![]()
where, A, B, C and D are 205 days weight (kg), the weaning weight (kg), the birth weight (kg) and the weaning age (days) respectively.
Association of GH and MSTN genotypes with body weight: Statistical analysis was accomplished using Graphpad statistical software program (Graphpad prism for windows version 5.1, Graphpad software, Inc, Sandiego, CA, USA). General Linear Model (GLM) practice of the statistical analysis system package SAS31 was used for to elucidate association between GH and MSTN genotypes and corrected body weight.
Gene and genotypic frequencies: Gene and genotypic frequencies were calculated based on the electrophoresis results, by allele counting32 then chi-square was carried out for the observed counted genotypes for assessment Hardy-Weinberg equilibrium status and demonstration of genotype distribution in the studied population.
RESULTS
Specific primers were used to amplify precise DNA fragments 329 and 1346 bp of GH and MSTN genes, respectively. Restriction analysis of 329 bp PCR products of GH gene digested with HpaII revealed three genotypes; AA genotype possess undigested fragment (329 bp), AB genotype has three fragments (329, 224 and 105 bp) and BB genotype has two fragments (224 and 105 bp) (Fig. 1).
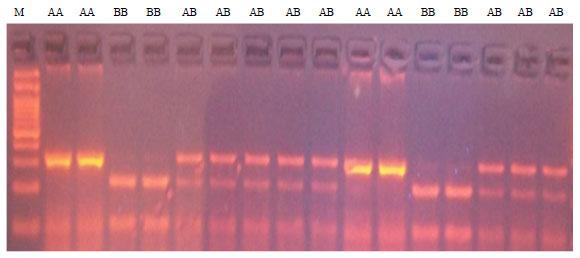 | |
| Fig. 1: | Representative HpaII restriction fragment pattern of GH gene (329 bp). AA: Restriction fragment of 329 bp, AB: Restriction fragment of 329, 224 and 105 bp, BB: Restriction fragment of 224 and 105 bp and M: DNA ladder 100 bp |
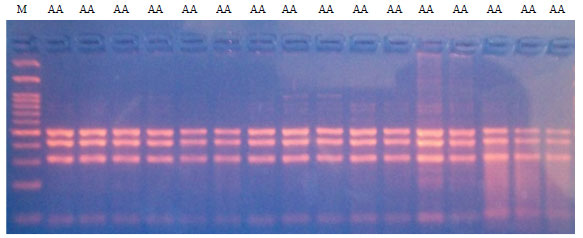 | |
| Fig. 2: | Representative DraI restriction fragment pattern of MSTN gene (1346 bp). AA: Restriction fragment of 505, 427, 321 and 93 bp, M: DNA ladder 100 bp |
| Table 2: | Frequency of genotypes and alleles in the GH locus |
 | |
| Chi square calculated (χ2) = 31.43 high significant differences, chi square tabulated (χ2) at DF 1 and p<0.05 = 3.84 | |
| Table 3: | Associations of GH genotypes with corrected body weight (Mean±SE) |
 | |
| Means of different levels within the same column having different superscripts are significantly different | |
Meanwhile, restriction analysis of 1346 bp PCR products of MSTN gene digested with DraI was monomorphic; where it yielded four fragments (505, 427, 321 and 93 bp) in all animals under study (Fig. 2).
The GH genotypes incidence and alleles frequency were calculated. For the 100 Friesian bull calves, genotypic frequencies for the AA, AB and BB genotypes were 0.1, 0.78 and 0.12, respectively and the allele frequencies for A and B allele frequencies were 0.49 and 0.51. The χ2-test exhibited that the GH genotypes distribution in the cattle population was not in Hardy-Weinberg equilibrium (p<0.05) (Table 2). Statistically significant GH genotypes effect on body weight was declared; where AB genotype possessed higher body weight trait than the other 2 genotypes. However, no significant variation was detected between AA and BB genotypes in the studied trait (body weight) (Table 3).
DISCUSSION
Growth rate represents economically important quantitative trait that affect carcass quantity in cattle. Genetic improvement of such trait has been accomplished using traditional methods of selection based on phenotypic information. However, candidate gene approach and molecular genetic markers could help to overcome some of the limitations of the traditional methods applied by animal breeder, genetic selection in this way has been very successful. The candidate gene approach refers to interpretation of polymorphisms of genes whose proteins are key enzymes involved vital physiological processes pathways33. When these genes are verified to be associated with particular economic trait, the existing traditional methods of trait selection have been substituted by the use of polymorphic specific genes that represent molecular detectable markers for economic traits of animals.
In this study, amplification of GH gene generated fragment length of 329 bp. The attained DNA cleaved fragments for GH-HpaII digestion were: Digested (224 and 105 bp) fragments for BB genotype, three fragments (329, 224 and 105 bp) for AB genotype and undigested fragment (329 bp) for AA genotype. The frequencies of the detected various GH-MspI genotypes in the 100 Friesian bull calves were 0.1, 0.78 and 0.12 for the AA, AB and BB genotypes respectively. The frequencies for A and B alleles were 0.49 and 0.51. The χ2-test presented the obtained GH genotypes distribution in the cattle population not in Hardy-Weinberg equilibrium (p<0.05). Genotype counts for the observed and the expected values showed high significant difference indicating the population is not balanced and not follow Hardy- Weinberg equilibrium. This may be attributed to small sample size and/or artificial selection of parents for high body weight34. The GH-HpaII locus was significantly associated with the desired trait of interest (body weight); where AB genotype was higher in body weight than the other 2 genotypes (AA and BB). Apposing results were attained by previous studies to determine the association of GH/HapaII variant and body weight in various cattle breed.
The GH/MspI genetic diversity within and between 8 different breeds of beef cattle was reported35-39. On contrast, MspI site in GH gene of Aceh cattle was monomorphic40. The frequency of AA genotype was 1.00 and same as A allele frequency.
Association between GH/MspI locus genetic polymorphism and Grati dairy cow’s body weight was studied16. The results showed that frequencies of A and B allele were 0.34 and 0.66, respectively. There was significant association between GH/MspI genotypes and body weight (p<0.05). Also, GH/MspI variant effect on body weight was identified in Ongole-crossbred cattle17. Results showed that cows and their subsequent progenies possessing the AB genotype performed the favorable body weight compared with BB and AA homozygous genotypes. Another study on GH/MspI polymorphism and body weight, intron 3 of the GH gene polymorphism and body weight at first estrus and first calving in Holstein heifers in Antioquia was investigated18. Allele frequencies for the alleles (A) and (B) were 0.91 and 0.09, respectively. The genotype frequencies were 0.77, 0.2 and 0.03 for the genotypes (BB), (AB) and (BB), respectively. There was an association between genotype and weight at first estrus and first calving (p<0.01). On contrary, GH/MspI locus did not significantly affect the body weight and measurements of West Sumatera Pesisir cattle19.
In the present study, PCR-DraI digestion of 1346 bp of MSTN gene was monomorphic; where it yielded four fragments (505, 427, 321 and 93 bp) in all animals under study; suggesting that MSTN-DraI locus did not affect the body weight. The MSTN-DraI locus polymorphism relatedness with body weight is scare. In three Chinese cattle breeds, PCR-DraI digestion of 1346 bp of MSTN gene was polymorphic29. The following DNA fragments were attained according this previous study; fragment lengths of 505, 427, 321 and 93 bp for genotype AA, 505, 365, 321, 93 and 62 bp for genotype BB and 505, 427, 365, 321, 93 and 62 bp for genotype AB. According to results, MSTN-DraI locus could affect body weight. Also, MSTN gene g-371 T>A promoter polymorphism and its association with carcass trait was screened in Holstein and Korean cattle breeds. Significant differences (p<0.05) were found between the MSTN genotypes and all carcass traits, except the live weight, back fat thickness, eye muscle area, carcass weight, marbling score, or meat color index41 .
The limitation of the present study should be acknowledged. First, small sample size may not allow obtaining concrete conclusion. Second, limited number of candidate gene markers may influence the conclusion. Accordingly, such shortcoming should be considered in further investigations.
CONCLUSION
This pilot study highlights the significance effect of GH/HpaII locus as candidate for body weight in cattle rather than MSTN/DraI. Moreover, GH/HapaII locus can be utilized as a marker for early selection of high body weight animals and early culling of lower body weight animal’s results in preventing economic losses afforded by the latter. Further studies need to be done on a large sample size and wide range of cattle breed to establish the association between GH/HpaII and MSTN/DraI gene polymorphisms and body weight.
ACKNOWLEDGMENT
Many cardinal thanks from the authors to all members of department of animal husbandry and animal wealth development, faculty of veterinary medicine, Damanhour university, Egypt for their kind continuous cooperation .
REFERENCES
- Hua, G.H., S.L. Chen, J.N. Yu, K.L. Cai and C.J. Wu et al., 2009. Polymorphism of the growth hormone gene and its association with growth traits in Boer goat bucks. Meat Sci., 81: 391-395.
CrossRefDirect Link - Pal, A., A.K. Chakravarty, T.K. Bhattacharya, B.K. Joshi and A. Sharma, 2004. Detection of Polymorphism of growth hormone gene for the analysis of relationship between allele type and growth traits in Karan Fries cattle. Asian-Austr. J. Anim. Sci., 17: 1334-1337.
CrossRefDirect Link - Rothschild, M., C. Jacobson, D. Vaske, C. Tullge and L. Wang et al., 1996. The estrogen receptor locus is associated with a major gene influencing litter size in pigs. Proc. Natl. Acad. Sci. USA., 93: 201-205.
Direct Link - Parmentier, I., D. Portetelle, N. Gengler, A. Pradi and C. Bertozzi et al., 1999. Candidate gene markers associated with somatotropic axis and milk selection. Domestic Anim. Endocrinol., 17: 139-148.
PubMedDirect Link - Lingappa, V.R., A. Devillers-Thiery and G. Blobel, 1977. Nascent prehormones are intermediates in the biosynthesis of authentic bovine pituitary growth hormone and prolactin. Proc. Nat. Acad. Sci., 74: 2432-2436.
Direct Link - Hediger, R., S.E. Johnson, W. Barendse, R.D. Drinkwater, S.S. Moore and J. Hetzel, 1990. Assignment of the growth hormone gene locus to 19q26-qter in cattle and to 11q25-qter in sheep by in situ hybridization. Genomics, 8: 171-174.
CrossRefDirect Link - Gordon, D.F., D.P. Quick, C.R. Erwin, J.E. Donelson and R.A. Maurer, 1983. Nucleotide sequence of the bovine growth hormone chromosomal gene. Mol. Cell. Endocrinol., 33: 81-95.
CrossRefDirect Link - Zakizadeh, S., G. Rahimi, S.R. Mirae-Ashtiani, A. Nejati-Javaremi and M. Moradi-Shahrbabak et al., 2006. Analysis of bovine growth hormone gene polymorphisms in three Iranian native breeds and Holstein cattle by RFLP-PCR. Biotechnology, 5: 385-390.
CrossRefDirect Link - Lucy, M.C., S.D. Hauser, P.J. Eppard, G.G. Krivi, J.H. Clark, D.E. Bauman and R.J. Collier, 1993. Variants of somatotropin in cattle: Gene frequencies in major dairy breeds and associated milk production. Domest. Anim. Endocrinol., 10: 325-333.
PubMedDirect Link - Zhang, H.M., D.R. Brown, S.K. DeNise and R.L. Ax, 1993. Rapid communication: Polymerase chain reaction-restriction fragment length polymorphism analysis of the bovine somatotropin gene. J. Anim. Sci., 71: 2276-2276.
PubMedDirect Link - Yao, J., S.E. Aggrey, D. Zadworney, J.F. Hayes and U. Kuhnlein, 1996. Sequence variations in the bovine growth hormone gene characterized by single-strand conformation polymorphism (SSCP) analysis and their association with milk production traits in Holsteins. Genetics, 144: 1809-1816.
PubMedDirect Link - Ayuk, J. and M.C. Sheppard, 2006. Growth hormone and its disorders. Postgrad. Med. J., 82: 24-30.
CrossRefPubMedDirect Link - Beauchemin, V.R., M.G. Thomas, D.E. Franke and G.A. Silver, 2006. Evaluation of DNA polymorphisms involving growth hormone relative to growth and carcass characteristics in Brahman steers. Genet. Mol. Res., 5: 438-447.
PubMedDirect Link - Maylinda, S., 2011. Genetic polymorphism of growth hormone locus and its association with bodyweight in Grati dairy cows. Int. J. Biotechnol. Mol. Biol. Res., 2: 117-120.
Direct Link - Paputungan, U., L. Hakim, G. Ciptadi and H.F.N. Lapian, 2013. Polymorphism of growth hormone msp1 enzyme-restriction associated with production performance of ongole-crossbred cattle mated by artificial insemination technique. J. Basic. Applied Sci. Res., 3: 581-589.
Direct Link - Arango, J., J.J. Echeverri and A. Lopez, 2014. Association between a polymorphism in intron 3 of the bovine growth hormone gene and growth traits in Holstein heifers in Antioquia. Genet. Mol. Res., 13: 6191-6199.
Direct Link - Jakaria, J., D. Duryadi, R.R. Noor and B. Tappa, 2007. The relationship of MspI growth hormone gene polymorphism and body weight and body measurements of west Sumatera Pesisir cattle. J. Indonesian Trop. Anim. Agric., 32: 33-40.
Direct Link - Lee, S.J. and A.C. McPherron, 2001. Regulation of myostatin activity and muscle growth. Proc. Natl. Acad. Sci. USA., 98: 9306-9311.
CrossRefPubMedDirect Link - McPherron, A.C. and S.J. Lee, 1997. Double muscling in cattle due to mutations in the myostatin gene. Proc. Nat. Acad. Sci. USA., 94: 12457-12461.
PubMedDirect Link - Charlier, C., W. Coppieters, F. Farnir, L. Grobet and P.L. Leroy et al., 1995. The mh gene causing double-muscling in cattle maps to bovine chromosome 2. Mammalian Genome, 6: 788-792.
CrossRefDirect Link - Kambadur, R., M. Sharma, T.P.L. Smith and J.J. Bass, 1997. Mutations in Myostatin (GDF8) in double-muscled belgian blue and piedmontese cattle. Genome Res., 7: 910-915.
PubMedDirect Link - Cappucio, I., C. Marchitelli, A. Serracchioli, A. Nardone, F. Filippini, P. Ajmone-Marsan and A. Valentini, 1998. A G-T transversion introduces a stop codon at the mh locus in hypertrophic Marchigiana beef subjects. Anim. Genet., 29: 51-51.
Direct Link - Grobet, L., L.J.R. Martin, D. Poncelet, D. Pirottin and B. Brouwers et al., 1997. A deletion in the bovine myostatin gene causes the double-muscled phenotype in cattle. Nat. Genet., 17: 71-74.
CrossRefPubMedDirect Link - Smith, T.P.L., N.L. Lopez-Corrales, S.M. Kappes and T.S. Sonstegard, 1997. Myostatin maps to the interval containing the bovine mh locus. Mammalian Genome, 8: 742-744.
CrossRefDirect Link - Jeanplong, F., M. Sharma, W.G. Somers, J.J. Bass and R. Kambadur, 2001. Genomic organization and neonatal expression of the bovine myostatin gene. Mol. Cell. Biochem., 220: 31-37.
CrossRefDirect Link - Shibata, M., K. Ohshima, T. Kojima, T. Muramoto and K. Matsumoto et al., 2003. Nucleotide sequence of myostatin gene and its developmental expression in skeletal muscles of Japanese Black cattle. Anim. Sci. J., 74: 383-390.
CrossRefDirect Link - Zhang, R.F., H. Chen, C.Z. Lei, C.L. Zhang and X.Y. Lan et al., 2007. Association between polymorphisms of MSTN and MYF5 genes and growth traits in three Chinese cattle breeds. Asian-Austr. J. Anim. Sci., 20: 1798-1804.
Direct Link - Szabo, F., E. Szaboand and S. Bene, 2012. Statistic and genetic parameters of 205-day weaning weight of beef calves. Arch. Tierz., 6: 552-561.
Direct Link - Falconer, D.S. and T.F.C. Mackay, 1996. Introduction to Quantitative Genetics. 4th Edn., Prentice Hall, Harlow, England, ISBN-13: 9780582243026, Pages: 464.
Direct Link - Womack, J.E., 1993. The goals and status of the bovine gene map. J. Dairy Sci., 76: 1199-1203.
CrossRefDirect Link - Wang, J., G. Li, M.A. Elzo, L. Yan, S. Chen, X. Jia and S. Lai, 2015. A novel single nucleotide polymorphism of the POU1F1 gene associated with meat quality traits in rabbits. Ann. Anim. Sci., 15: 611-620.
CrossRefDirect Link - Sutarno, S., 1999. Genetic diversity within and between breeds of beef cattle 2. Growth hormone gene. BioSMART: J. Biol. Sci., 1: 1-7.
Direct Link - Zhou, G.L., Q.I. Zhu, H.G. Jin and S.L. Guo, 2006. Genetic variation of growth hormone gene and its relationship with milk production traits in China holstein cows. Asian-Aust. J. Anim. Sci., 19: 315-318.
Direct Link - Maylinda, S., W. Busono, H. Nugroho, Marijono and E. Romjali, 2008. Genetic polymorphism in growth hormones locus and its relationship with digest ability of low quality feed. J. Ilum-Ilmu Hayati (Life Sci.), 20: 63-70.
Direct Link - Kmiec, M., E. Krzecio, A. Terman, M. Kocwin-Podsiadla and T. Grzelak, 2010. Differentiation of fatteners carcass quality traits depending on GH/MspI polymorphism. Annales Universitatis Mariae Curie-Skłodowska Sectio EE Zootechnica, 28: 31-40.
Direct Link - Paputungan, U., L. Hakim, G. Ciptadi and H.F.N. Lapian, 2012. The allele frequencies of growth hormone gene on the parental and progeny of Ongole-crossbreed cattle population in the North Sulawesi of Indonesia using PCR-RFLP. J. Evolutionary Biol. Res., 4: 52-58.
CrossRefDirect Link - Putra, W.P.B., T. Hartatik and S. Sumadi, 2013. Growth hormone gene genotyping by MspI restriction enzyme and PCR-RFLP methods in aceh cattle breed at indrapuri district of aceh province. J. Indonesian Trop. Anim. Agric., 38: 207-211.
Direct Link - Han, S.H., I.C. Cho, M.S. Ko, E.Y. Kim, S.P. Park, S.S. Lee and H.S Oh, 2012. A promoter polymorphism of MSTN g.-371T> A and its associations with carcass traits in Korean cattle. Mol. Biol. Rep., 39: 3767-3772.
CrossRefDirect Link