
M.M. Muharram
Department of Pharmacognosy, College of Pharmacy, Prince Sattam Bin Abdulaziz University, Alkharj 11942, Kingdom of Saudi Arabia
Department of Microbiology, College of Science, Al-Azhar University, Nasr City 11884, Cairo, Egypt
LiveDNA: 20.7742
Pakistan Journal of Biological Sciences
Year: 2016 | Volume: 19 | Issue: 3 | Page No.: 106-114







ABSTRACT
Recombinant methioninase (rMETase) is an enzyme that has antitumor activity. In this work, METase gene from Pseudomonas putida ATTCC 8209 was cloned to pT7-7 plasmid (yielded, PT7-METase-R7 clone) and expressed in E. coli strain BL21 (DE3). A protein band with a molecular massof 42 kDa was visualized by SDS-PAGE. The applied protocol yielded a total protein of 3.13 g with a recovery of 66.89% and a specific activity of 18.59 U mg–1 which considered as a low yield. However, when the METase gene was cloned to the vector (pTrc99A, clone: pTrc99A-MET-3) cells of E. coli JM109 yielded a total protein of 32.63 g with a recovery of 41.62% and a specific activity of 54.86 U mg–1 which revealed that the enhancement of METase gene expression by trc promoter was more than the T7 RNA polymerase promoter. The t1/2 of the rMETase was 2 h asanalyzed in mice by IV injection. Antitumor efficacy of rMETase was studied in five human cancer cell lines. At 1 U mL–1 the growth rate of treated colon cancer cell lines, Colo205 and SW620, with rMETase was 46 and 32% relative to control, respectively. With the ovarian cancer cell line (A2780) rMETase produced an inhibition effect of 54% at 1.5 U mL–1. In addition, the growth rate was reduced to 45 and 53% with the skin cancer cell line (A375) and the breast cancer cell line (MCF-7), respectively. These results indicate the feasibility of rMETase for use as a potent antitumor agent.
PDF Abstract XML References Citation
Received: November 28, 2015;
Accepted: January 12, 2016;
Published: February 15, 2016
How to cite this article
M.M. Muharram, 2016. Recombinant Engineering of L-Methioninase Using Two Different Promoter and Expression Systems and in vitro Analysis of Its Anticancer Efficacy on Different Human Cancer Cell Lines. Pakistan Journal of Biological Sciences, 19: 106-114.
DOI: 10.3923/pjbs.2016.106.114
URL: https://scialert.net/abstract/?doi=pjbs.2016.106.114
DOI: 10.3923/pjbs.2016.106.114
URL: https://scialert.net/abstract/?doi=pjbs.2016.106.114
INTRODUCTION
The complete dependence of the tumor cells on L-methionine for their proliferation was frequently reported by several researchers (Hoffman, 1984, 2015; Anderson, 1998; Hu and Cheung, 2009; Van Rite et al., 2011; Stone et al., 2012; Yano et al., 2014; Huang et al., 2015). Depletion of any essential amino will disrupt proteinsynthesis, but L-Met is also required for polyamine synthesis a known contributor to tumorigenesis (Megosh et al., 1995; Thomas and Thomas, 2003), furthermore it is the major methylation source for DNA and other molecules (Lu and Epner, 2000). When L-Met levels decrease from the normal human serum concentration of ~30 μM (Stabler et al., 1987) to a threshold around ~5 μM these tumors are unable to survive (Kokkinakis et al., 1997). In vivo, a significant decrease in serum L-Met can be achieved by the systemic application of rMETase from Pseudomonas putida that degrades L-methionine to α-ketobuty rate, methane thiol and NH3 resulting in drastic retardation of tumor growth in a variety of animal models and has been shown to have synergistic effects in combination with chemotherapeutic agents such as 5-fluorouracil and vincristine (Lishko et al., 1993; Tan et al., 1996, 1997; Yoshioka et al., 1998; Yang et al., 2004; Hu and Cheung, 2009).
L-Methioninase enzyme (METase; EC 4.4.1.11) is ubiquitous in almost of organisms, including bacteria, fungi, protozoa and plants except mammals (Cooper, 1983; El-Sayed, 2010). It is a pyridoxal 5-phosphate (PLP)-dependent multifunctional enzyme which catalyzes α, γ-elimination of L-methionine to α-ketobutyrate, methane thiol and ammonia (Tanaka et al., 1977). The L-methioninase has received affordable attention as a therapeutic agent against various types of methionine dependent tumors (Kokkinakis et al., 1997; Yoshioka et al., 1998; Yano et al., 2014; Huang et al., 2015). Regarding to its nature, L-methioninase was comprehensively characterized from many bacterial species as intracellular enzyme (Tanaka et al., 1976) and from fungal species as intracellular and extracellular enzyme (El-Sayed, 2009). The gene encoding METase from Pseudomonas putida was cloned and expressed in Escherichia coli (Inoue et al., 1995), the mechanism of action was characterized by using site-directed mutagenesis (Inoue et al., 2000) and the assay method was constructed on the basis of the reaction mechanism (Takakura et al., 2004).
The enzyme composed of 398 amino acid residues with a calculated molecular weight of 42,626 Daltonscor responding to the subunit of the homotetramer (Inoue et al., 1995). The crystal structure of the enzyme was determined by X-ray diffraction analysis (Motoshima et al., 2000). Hori et al. (1996) also reported the cloning of a gene from P. putida and the peptide sequence deduced from the sequence of this gene has 398 amino acids with a molecular mass of 42,720 Daltons. However, this gene and corresponding protein differ significantly in sequence from that reported by Inoue et al. (1995). The first goal of this study was the cloning and expression of METase gene of Pseudomonas putida in E. coli. The second goal was to use a protocol for high-yield of rMETase and to investigate the antitumor efficacy of the expressed protein against human tumor cell lines in vitro.
MATERIALS AND METHODS
Materials: Restriction endonuclease, T4 DNA ligase and BL21 (DE3) competent cells were purchased from Strategene (San Diego, CA). The Gene Amp PCR reagent kit was purchased from Roche (Branchburg, NJ). The DNA purification kit was purchased from Promega (Madison, WI). The pT7 Blue T-vector was purchased from Novagen (Madison, WI). All other chemicals were obtained from Sigma Chemical Co. (St. Louis, MO). Cell Lines and strain of P. putida ATTCC 8209 was purchased from the American Type Culture Collection (Rockville, MD).
Cloning of the L-methioninase gene from Pseudomonas putida using the PT7Blue T-vector: Based on the sequence of the METase gene as determined by Inoue et al. (1995), two oligonucleotide primers were synthesized. The primers used were: P1, 5'GCCGGTCTGTGGAATAAGCT3' and P2, 5'CCAGGGTCGACTCCAGCGCC 3'. The PCR reaction conditions were as follows: First denaturation at 95°C for 10 min, then 5 cycles of denaturation at 94°C for 30 sec, annealing at 60°C for 30 sec and extension at 72°C for 2 min. This was followed by 25 cycles of denaturation at 94°C for 30 sec, 60°C for 30 sec, then extension at 72°C for 1.5 min, then a final extension at 72°C for 10 min. The PCR-amplified product 1365 bp band was purified and sequenced and contained the METase gene. Amplified METase gene was ligated to the PT7Blue T-vector (Novagene) at the EcoRV T-cloning site. The resulting plasmid PT7-METase-R was introduced to the cells of E. coli DH5-α using the standard transformation procedure. To construct an over expression vector, the PT7-METase-R was used as a template DNA with the following oligonucleotide primers for the PCR reaction P3-5'GGAATTCCATATGCACGGCTCCAACAAGC-3' and p4 5'AGTCATGGATCCTCAGGCACTCGCCTTGAGTGC-3'. The PCR-amplified product (1220 bp), which contained the METase gene with sites modified for the NdeI and BamH I restriction enzymes, was collected and purified. This DNA sequence was termed PT7-METase-R7. The PT7-METase-R7 was digested with the NdeI and BamH I-enzymes and ligated into the pT7-7 vector at the NdeI and BamH I-cloning sites using standard protocols. The resulting plasmid was then transformed into E. coli BL21 (DE3) competent cells. Positive clones were selected from ampicillin containing plates. After storage at 4°C for 24 h the clones which expressed high levels of rMETase had a distinct yellow-orange color due to high enrichment of the pyridoxal phosphate-containing rMETase. The overexpression clones were first selected by their color and then by activity assay. All yellow-orange colonies were positive for METase and non-colored clones were METase negative.
Cloning of the L-methioninase gene from Pseudomonas putida using Escherichia coli JM109 and pTrc99A vector: Cells of E. coli JM109 was used as host strain for expression of recombinant METase. METase gene from isolated chromosomal DNA of P. putida ATTCC 8209 was amplified by PCR with 5'-CCCGGTACCACGGCTCCAACAAGCTCCCAG-3' and reverse 5'-CTCGAGACGGGTTCAGGCACTCGCCTT-3' primers according to Inoue et al. (1995). Amplified METase gene was inserted into pMOS blue vector (Amersham Biosciences, Piscataway, NJ). The fragment treated with KpnI-T4 DNA polymerase and XbaI including METase gene to pTrc99A vector (Amersham Biosciences) yielding pTrc99A-MET-3.
Fermentation: It was started with 10 μL of IPTG induced cells of E. coli, harboring the vector PT7-METase-R7, into 5 mL LB medium with 100 μg ampicillin at 37°C at 400 rpm overnight. This culture was transferred to 800 mL Terrific Broth (TB) and grown overnight at 37°C at 400 rpm then transferred to 10 800 mL TB medium in 6 L flasks with shaking at 400 rpm and 37°C for 16 h at which the OD600 was 10. Incubation was continued for extra 6 h after the medium was changed with fresh TB until the OD600 reached to 20.
Enzyme purification: The bacteria cells were harvested, after fermentation in production culture, by centrifugation at 4000 g and 4°C for 10 min. The pellet was suspended in extraction solution contained (20 mM potassium phosphate, pH 9.0, 10 mM pyridoxal phosphate and 0.01% b-mercapto ethanol) and disrupted with a homogenizer. Heat treatment of the homogenate was then carried out up to 50°C for 1 min. Suspension was centrifuged at 8000 g and 4°C for 30 min. The supernatant was then collected and filtrated by ultrafiltration using a Millipore (Bedford, MA) Prep/Scale-TFF PLHK 100 k, 2.5 ft2 cartridge with buffer (10 mM potassium phosphate, pH 8.3). The pH was adjusted to 7.2 by ultrafiltration.
DEAE sepharose chromatography: First column: DEAE Sepharose FF (pH 7.2) was 100 mm diameter and 30 cm height, with a volume of 2400 mL of DEAE sepharose FF (Pharmacia, Uppsala, Sweden). The elution flow rate was 15-50 mL min–1. Forty to eighty grams of total protein (10- 20 g mL–1 was applied on the column. The column was washed by with 40 mM potassium chloride in PPM buffer contained (10 mM potassium phosphate, pH 7.2, 10 μM pyridoxal phosphate and 0.01% β-mercaptoethanol) for 10 vol until the until the OD280 dropped below 0.1 dropped below 0.1. The protein was then eluted with a linear gradient of 40-300 mM potassium chloride in PPM buffer. Elution fractions of 500 mL were collected. The fractions containing rMETase were identified by yellow color and activity assay. Second column: DEAE sepharose FF has a height of 25 cm, with a volume of 500 mL. The elution flow rate was 6-8 mL min–1. Ten grams of total protein (2-5 mg mL–1) were applied on the second DEAE sepharose FF column after 24 h dialysis in 80 mM potassium chloride and 10 mM potassium phosphate (pH 8.3). After loading, the column was washed with 80 mM potassium chloride and 10 mM potassium phosphate (pH 8.3) for about 4 volumes until the OD280 dropped below 0.1. rMETase was eluted with a buffer of linear gradient of (80-300 mM potassium chloride in 10 mM potassium phosphate buffer (pH 8.3). Elution fractions of 300 mL were collected and the the fractions containing rMETase were identified by yellow color and activity assay.
Enzyme activity: It was carried out according to the method of Esaki and Soda (1987) with the slight modification that previously described (Takakura et al., 2004). The enzyme solution was prepared by appropriate dilution with 100 mM potassium phosphate buffer (pH 8.0) containing 0.01% dithiothreitol, 1 mM EDTA, 10 μM pyridoxal 5'phosphate (PLP) and 0.05% Tween 80. One milliliter of the substrate solution consisting of 100 mM potassium phosphate buffer (pH 8.0), 25 mM L-methionine and 10 μM PLP was preincubated at 37°C in a glass tube with a sealed cap. The enzymatic reaction was initiated by adding 50 μL of the enzyme solution to the preincubated substrate solution at 37°C. After incubating the reaction mixture for precisely 10 min in a glass tube with a sealed cap, the enzymatic reaction was terminated by adding 100 μL of 50% trichloroacetic acid. Subsequently, the chemical reaction was initiated by adding both 1.6 mL of 1 M sodium acetic acid buffer (pH 5.0) and 0.6 mL of 0.1% 3-methyl-2-benzothiazolinone hydrazone hydrochloride monohydrate (MBTH) into 0.8 mL of the above enzymatic reaction mixture. After incubating the mixture at 50°C for 40 min in a glass tube with a sealed cap and cooling it down to 25°C, the amount of α-ketobutyrate produced was measured spectrophotometrically. One unit of METase was defined as the amount of enzyme that produced 1 μmol of α-ketobutyrate per minute at infinite concentration of L-methionine. The specific activity was calculated as units/mg protein.
ActiClean endotoxin affinity: To eliminate endotoxin, 20 mg protein/mL of purified rMETase in 250 mL was applied on an 800 mL ActicleanEtox column (Sterogen, CA). The protein was eluted with a buffer contained (0.12 M sodium chloride in 10 mM sodium phosphate, pH 7.2), at a flow rate of 1 mL min–1. Enzyme fractions, identified by yellow color and activity assay, were collected. The final eluant was concentrated with 30 k AmiconCentriprep concentrators by centrifugation at 4000 g 4°C for 30 min. Sterilization was performed with 0.2 μm Nylon filter (Nalgene).
Limulus Amebocyte Lysate (LAL) test endotoxin assay: A sample was mixed with the LAL and incubated at 37°C for 10 min. A substrate solution supplied with the kit was then mixed with the sample and incubated at 37°C for an additional 6 min. The reaction was stopped with stop reagent supplied with the kit. The absorbance of the sample was determined with a spectrophotometer at 410 nm. The concentration of endotoxin was calculated from a standard curve which was constructed from the endotoxin supplied in the kit at concentrations from 0.1-1 EU mL–1.
In vivo determination of the half-life of rMETase: This test of the half-life of rMETase was analyzed in mice. Forty units of pure rMETase were injected into the tail vein. Blood samples were collected before the injection and at 20 min, 40 min, 1, 2, 3, 5, 6, 7, 8 and 9 h after injection. Blood samples were collected from three mice at each time point.
In vitro effect of rMEtase on different human cancer cell lines: Human cancer cell lines of various origins (colon cancer; Colo 205 and SW 620; ovarian cancer: A2780; breast cancer: MCF7 and skin cancer: A375 were cultured in DMEM or RPMI 1640 (Life Technologies, Inc.) containing l0% dialyzed horse serum in 96 well plates at an initial density of 1×I05 cells mL–1 for cancer cell lines. The cells were treated with the purified enzyme at 0-2.5 U mL–1. The viability of cultured cell lines was measured spectrophotometrically with 3-(4.5- dimethylthiazol-2-yl)-2.5-diphenyl-tetrazolium bromide and with erythrocin B dye exclusion method, respectively, after a 4 day incubation. The growth of cells treated with different concentrations of the enzyme was expressed as the percentage of growth compared to that in cells without treatment.
Miscellaneous: All DNA manipulation protocols, restriction analysis, electro-elution of DNA fragments, ligation and DNA sequence determination were performed by the standard methods as described in Sambrook et al. (1989). Polyacrilamide gel electrophoresis (SDS-PAGE) as described by Laemmli (1970). Protein concentration was determined according to Bradford (1976). The prestained protein markers (Biolabs, England), broad range 14.4-116.2 kDa were used in detection of molecular masses. Immuno-detection of proteins was conducted according to Towbin et al. (1979).
RESULTS
PCR cloning of rMETase gene from Pseudomonas putida in Escherichia coli: Chromosomal DNA of P. putida ATTCC 8209 was used to amplify the segment coding for the METasegene by PCR as described in the experimental procedures. The PCR amplification yielded a DNA band of 1220 bp (Fig. 1) which was identified by HI fragment DNA sequencing as the METase gene. The PCR product was introduced to the PT7Bue T-vector as NdeI/Bam HI fragment L-fragment to yield the PT7-METase-R7 clone. This clone was used for the transformation of E. coli BL21 (DE3).
Expression the rMETase: Positive clones which expressed high levels of rMETase had a distinct yellow-orange color when the transformation plates were incubated for 24 h at 4°C due to high enrichment of the pyridoxal phosphate-containing rMETase. The overexpression clones were first selected by their color (Fig. 2) and then by activity assay. Expressed rMETaserepresented 11.68% of the total protein based on the specific activity of 18.59 U mg–1. A protein band with a molecular masscorresponding to ~42 kDa was visualized by SDS-PAGE (Fig. 3, lane 2).
 | |
| Fig. 1: | PCR amplification of METase gene, chromosomal DNA of P. putida ATTCC 8209 served as a template DNA. Three microliter of PCR product was analyzed on 0.7% agarose: Lane 1, DNA standard (Roche, Mannheim) lane 2 and PCR product (1220 bp) |
Fermentation and purification of rMETase: Fermentation was started with 10 μL of IPTG induced E. coli cells BL21 (DE3), harboring the vector PT7-METase-R7, into 5 mL LB medium with 100 μg ampicillin at 400 rpm and 37°C overnight. This culture was transferred to 1 L mL TB broth in 6 L flask and grown overnight at 37°C at 400 rpm and transferred to 6 L fermentor. Incubation was continued and accompanied by refreshing the TB medium until the OD600 reached to 20. Bacterial cells were then harvested by centrifugation and then lysed. The rMETase was then purified by heating to 50°C for 1 min and ultrafiltration followed by chromatography on two DEAE Sepharose Fast Flow columns one at pH 7.2 and the other at 8.3, as described under procedures. Purification analysis of rMETase by SDS-PAGE showed a single protein band of approximately 42 kDa (Fig. 4, Lane 3).
As seen in Table 1, the purification procedure of rMETase yielded a total protein of 17.65 g after heat treatment step and the ultrafiltration with a recovery of 93% and specific activity of 4.60 U mg–1.
Specific activity increased from 4.6-18.59 U mg–1 after elution on the second DEAE Sepharose FFcolumn step. ActiClean endotoxin affinity step could reduce endotoxin levels from 59-0.43 ng mg–1 protein.
Expression of rMETase using the Escherichia coli JM109 and pTrc99A vector: Data presented in Table 1 showed a fantastic protocol of laboratory-scale preparation of rMETase. However, for therapeutic use of recombinant METase, the enzyme needs to be manufactured on a large-scale, in high yields and quality and with low cost.
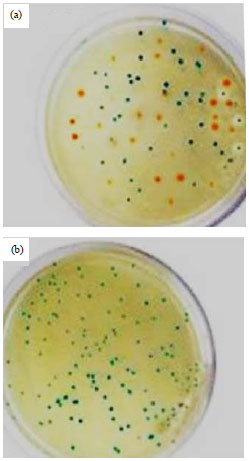 | |
| Fig. 2(a-b): | Expression of rMETase in Escherichia coli cells of BL21 (DE3). Positive clones which expressed high levels of rMETase had a distinct yellow-orange color due to high enrichment of (a) cells transformed with the empty vector of (PT7Blue) were used as a control plate and (b) Pyridoxal phosphate-containing rMETase |
 | |
| Fig. 3: | Stained SDS-PAGE with coomassie brilliant blue showing the expression of the rMETase, lane, 1 and 4; protein marker, lane 2; extracted total protein after IPTG induction while lane, 3; without induction, arrow indicates to the expressed protein band at 42 kDa |
| Table 1: | Purification of the recombinant METase from Escherichia coli BL21 (DE3) with PT7-METase-R7 |
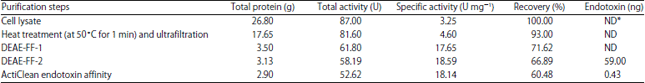 | |
| *ND: Not determined | |
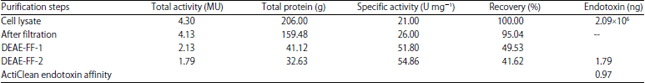
| Table 2: | Purification of rMETase from Escherichia coli JM109 with pTrc99A-MET-3 |
 | |
 | |
| Fig. 4: | Stained SDS-PAGE with coomassie brilliant blue showing the purification of the rMETase; lane 1; protein marker, lane 2; supernatant of the cell lysate of the IPTG induced Escherichia coli cells that harboring the vector PT7-METase-R7, lane 3; purified protein of rMETase at about 42 kDa |
In spite of the promising data and the applicable protocol of gram-scale preparation of rMETase, it is difficult to be applied in the industrial and commercial production using the T7 RNA promoter in Escherichia coli.
In a trial to overcome this issue, cells of E. coli BL21 (DE3) were replaced by E. coli JM109 as a host strain for expression of rMETase. However, results of E. coli JM109 with PT7-METase-R7 were not significantly different from those of E. coli BL21 (DE3) (data not shown). Therefore, a new expression plasmid (pATG3131), carrying the trc promote, was constructed according to a previous study by Inoue et al. (1995).
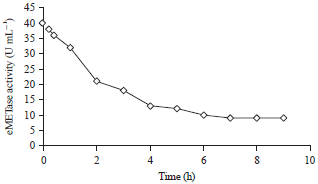 | |
| Fig. 5: | In vivo determination of the half-life of rMETase |
In this plasmid, amplified METase gene by PCR was treated with KpnI and XbaI and ligated to pTrc99A vector resulting in pTrc99A-MET-3. Escherichia coli JM109 cells which harboring pTrc99A-MET-3 were cultivated with shaking and the pH was maintained at 7.2. IPTG was added 4 h after fermentation (30 L) at a final concentration of 0.30 mM. Table 2 summarizes the purification results of rMETase from E. coli JM109 with pTrc99A-MET-3. The experiment revealed that the trc promoter in the pTrc99A vector enhanced the METase gene expression several of times more than the PT7Bue T vector based on the total and specific activities. Half-life of rMETase was analyzed in mice. Upon administration of rMETase by iv injection in mice, half life time of the rMETaset1/2 was 2 h as shown in Fig. 5.
Effect of rMETase on different human cancer cell lines. Five human cancer cell lines were tested and all of them were sensitive to inhibition by rMETase. At the value of 1 U mL–1, the growth rate of Colo 205 and SW 620 cancer cells treated with the rMETase resulted in 46 and 32% relative to control, respectively. Inhibitory effect of the rMETase on ovarian cancer was conducted with the cell line A2780 over a concentration 1.5 U mL–1 where it produced an inhibition effect of 54%.
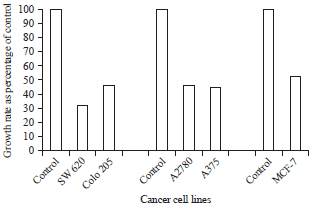 | |
| Fig. 6: | In vitro effect of rMETase on different human cancer cell lines |
As given in Fig. 6, the growth rate was reduced with rMETase to 45 and 53% in case of the skin cancer cell line (A375)and the breast cancer cell line (MCF-7), respectively.
DISCUSSION
Methionine is necessary for cancer cells compared to normal cells both in vitro and in vivo (Kreis and Goodenow, 1978; Ghoshal et al., 1986; Howarth and Wang, 2013; Sinha et al., 2014; Hoffman, 2015; Strekalova et al., 2015). The rMETase can be used to target some malignancies with minimal toxicity based on previous reports suggesting that it has antitumor activity (Tan et al., 1996, 1997; Zhao et al., 2006). This enzyme was searched in various bacterial species but the yield of pure enzyme was very low (Lishko et al., 1993). Therefore, the large-scale production of rMEtase was necessary.
In this work , we used a protocol for high-yield, pure and effective rMETase by high-level gene expression, large-scale fermentation and purification. The results revealed that yield of the rMETase reached to 11.68% (Table 1) of the total protein with cells of the E. coli BL21 and the pT7-7 overexpression plasmid which contains the strong T7 RNA polymerase promoter, based on the specific activity (18.59 U mg–1). Approximate accordance could be noticed with the results of the expressed rMETase from pAC1 clone (Tan et al., 1997) which gave rMETaseyield of 20 U mg–1 and 2.6 g pure protein (13.47%).
After fermentation in the production culture, Purification analysis of rMETase by SDS-PAGE and ultrafiltration chromatography on two DEAE sepharose fast flow columns showed a single protein band of approximately 43 kDa (Fig. 4). Although this clone showed a comparable results of laboratory-scale preparation of rMETase, the enzyme needs to be manufactured on a large-scale, in high yields and quality and with low cost. To overcome this issue, cells of E. coli BL21 (DE3) were replaced by E. coli JM109 as a host strain for expression of rMETase. However, results of E. coli JM109 with PT7-METase-R7 were not significantly different from those of E. coli BL21 (DE3) (data not shown). Therefore a new expression plasmid (pATG3131), carrying the trc promote, was used.
High-level expression and the purification process of the enzyme provided a cost-effective manufacturing process. Table 2 summarizes the yield of rMETase expressed in E. coli JM109 with pATG3131 plasmid that carrying the trc promoter. Using a 30 L L-fermentor, the developed clone, promoter pTrc99A-MET-3, yielded total protein of 159.48 g with a specific activity of 26 U mg–1 and a recovery of 95%. As seen in Table 2, the purification protocol of rMETase yielded a total protein of 32.63 with a recovery of 41.62% and specific activity of 54.86 U mg–1 after elution on the second DEAE Sepharose FFcolumn step. Applied protocol of rMETase revealed that the trc promoter in the pTrc99A vector enhanced the yield of rMETase (54.86 U mg–1) more than the PT7Bue T vector (18.59/mg). Produced rMETase produced in this study had a half-life of approximately 2 h in accordance with the result of Tan et al. (1997) and Takakura et al. (2006).
The relative cytotoxicity of a cancer therapeutic agent is a major concern for any cancer therapy. In order to determine the anticancer efficacy of the rMETase, purified proteins from the clone pTrc99A-MET-3 was examined with five different human cancer cell lines. The growth rate of the colon cancer, Colo 205 and SW 620 was 54 and 68% relative to control, respectively. In comparison with a previous report (Tan et al., 1999), the growth rate of Colo 205 and SW 620 cancer cells treated rMETase resulted in a growth rate of 73 and 88%, respectively. Cancer cells of the ovarian cell line A2780 exhibited a significant sensitivity to rMETase where their growth rate was inhibited to a value of 54% relative to the control. The methioninase protein inhibited the growth rate of the cancer cell lines of the skin (A375) and the breast (MCF-7) to 45 and 53%, respectively. Interestingly, inhibitory effects of rMETaseon cancer cell lines of A375 skin, A2780ovary and MCF-7 breast from previous studies (Palwai et al., 2009) reached to approximate value of 12, 23 and 11%, respectively.
The differences between the growth rate of the treated cells with rMETase and control demonstrated that rMETase could deplete tumor methionine levels.
CONCLUSION
The applied protocol for high-yield production, low toxicity and high efficacy of rMETase indicates the feasibility of this protein for use as a selective antitumor agent with high potential.
REFERENCES
- Anderson, M.E., 1998. Glutathione: An overview of biosynthesis and modulation. Chem. Biol. Interac., 111-112: 1-14.
CrossRefDirect Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Cooper, A.J.L., 1983. Biochemistry of sulfur-containing amino acids. Annu. Rev. Biochem., 52: 187-222.
CrossRefDirect Link - El-Sayed, A.S.A., 2009. L-methioninase production by Aspergillus flavipes under solid-state fermentation. J. Basic Microbiol., 49: 331-341.
CrossRefDirect Link - El-Sayed, A.S.A., 2010. Microbial L-methioninase: Production, molecular characterization and therapeutic applications. Applied Microbiol. Biotechnol., 86: 445-467.
CrossRefDirect Link - Esaki, N. and K. Soda, 1987. L-methionine γ-lyase from Pseudomonas putida and Aeromonas. Methods Enzymol., 143: 459-465.
CrossRefPubMedDirect Link - Ghoshal, A.K., D.S. Sarma and E. Farber, 1986. Ethionine in the Analysis of the Possible Separate roles of Methionine and Choline Deficiencies in Carcinogenesis. In: Essential Nutrients in Carcinogenesis, Poirier, L.A., P.M. Newberne and M.W. Pariza (Eds.). Springer, USA., ISBN: 978-1-4612-9025-4, pp: 283-292.
- Hoffman, R.M., 1984. Altered methionine metabolism, DNA methylation and oncogene expression in carcinogenesis: A review and synthesis. Biochimica Biophysica Acta (BBA)-Rev. Cancer, 738: 49-87.
CrossRefDirect Link - Hoffman, R.M., 2015. Development of recombinant methioninase to target the general cancer-specific metabolic defect of methionine dependence: A 40-year odyssey. Expert Opin. Biol. Ther., 15: 21-31.
CrossRefDirect Link - Hori, H., K. Takabayashi, L. Orvis, D.A. Carson and T. Nobori, 1996. Gene cloning and characterization of Pseudomonas putida L-methionine-α-deamino-γ-mercaptomethane-lyase. Cancer Res., 56: 2116-2122.
Direct Link - Howarth, G.S. and H. Wang, 2013. Role of endogenous microbiota, probiotics and their biological products in human health. Nutrients, 5: 58-81.
CrossRefDirect Link - Hu, J. and N.K.V. Cheung, 2009. Methionine depletion with recombinant methioninase: In vitro and in vivo efficacy against neuroblastoma and its synergism with chemotherapeutic drugs. Int. J. Cancer, 124: 1700-1706.
CrossRefDirect Link - Huang, K.Y., H.Y. Hu, Y.L. Tang, F.G. Xia, X.Q. Luo and J.Z. Liu, 2015. High-level expression, purification and large-scale production of L-methionine γ-lyase from Idiomarina as a novel anti-leukemic drug. Mar. Drugs, 13: 5492-5507.
CrossRefDirect Link - Inoue, H., K. Inagaki, M. Sugimoto, N. Esaki, K. Soda and H. Tanaka, 1995. Structural analysis of the L-methionine γ-lyase gene from Pseudomonas putida. J. Biochem., 117: 1120-1125.
Direct Link - Inoue, H., K. Inagaki, N. Adachi, T. Tamura, N. Esaki, K. Soda and H. Tanaka, 2000. Role of tyrosine 114 of L-methionine γ-lyase from Pseudomonas putida. Biosci. Biotechnol. Biochem., 64: 2336-2343.
CrossRefDirect Link - Kokkinakis, D.M., S.C. Schold Jr., H. Hori and T. Nobori, 1997. Effect of long‐term depletion of plasma methionine on the growth and survival of human brain tumor xenografts in athymic mice. Nutr. Cancer, 29: 195-204.
CrossRefDirect Link - Kreis, W. and M. Goodenow, 1978. Methionine requirement and replacement by homocysteine in tissue cultures of selected rodent and human malignant and normal cells. Cancer Res., 38: 2259-2262.
Direct Link - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Lishko, V.K., O.V. Lishko and R.M. Hoffman, 1993. Depletion of serum methionine by methioninase in mice. Anticancer Res., 13: 1465-1468.
PubMed - Lu, S. and D.E. Epner, 2000. Molecular mechanisms of cell cycle block by methionine restriction in human prostate cancer cells. Nutr. Cancer, 38: 123-130.
CrossRefDirect Link - Megosh, L., S.K. Gilmour, D. Rosson, A.P. Soler, M. Blessing, J.A. Sawicki and T.G. O'Brien, 1995. Increased frequency of spontaneous skin tumors in transgenic mice which overexpress ornithine decarboxylase. Cancer Res., 55: 4205-4209.
Direct Link - Motoshima, H., K. Inagaki, T. Kumasaka, M. Furuichi and H. Inoue et al., 2000. Crystal structure of the pyridoxal 5'-phosphate dependent L-methionine γ-lyase from Pseudomonas putida. J. Biochem., 128: 349-354.
CrossRefDirect Link - Palwai, N.R., X.P. Zang, R.G. Harrison, D. Benbrook and J.T. Pento, 2009. Selective growth inhibition of cancer cells by L-methioninase-containing fusion protein targeted to the urokinase receptor. Pharmacology, 84: 271-275.
CrossRefDirect Link - Sambrook, J., E.F. Fritsch and T.A. Maniatis, 1989. Molecular Cloning: A Laboratory Manual. 2nd Edn., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, USA., ISBN-13: 9780879695774, Pages: 397.
Direct Link - Sinha, R., T.K. Cooper, C.J. Rogers, I. Sinha and W.J. Turbitt et al., 2014. Dietary methionine restriction inhibits prostatic intraepithelial neoplasia in TRAMP mice. Prostate, 74: 1663-1673.
CrossRefDirect Link - Stabler, S.P., P.D. Marcell, E.R. Podell and R.H. Allen, 1987. Quantitation of total homocysteine, total cysteine and methionine in normal serum and urine using capillary gas chromatography-mass spectrometry. Anal. Biochem., 162: 185-196.
CrossRefDirect Link - Stone, E., O. Paley, J. Hu, B. Ekerdt, N.K. Cheung and G. Georgiou, 2012. De novo engineering of a human cystathionine-γ-lyase for systemic L-methionine depletion cancer therapy. ACS Chem. Biol., 7: 1822-1829.
CrossRefDirect Link - Strekalova, E., D. Malin, D.M. Good and V.L. Cryns, 2015. Methionine deprivation induces a targetable vulnerability in triple-negative breast cancer cells by enhancing trail receptor-2 expression. Clin. Cancer Res., 21: 2780-2791.
CrossRefDirect Link - Takakura, T., K. Mitsushima, S. Yagi, K. Inagaki and H. Tanaka et al., 2004. Assay method for antitumor L-methionine γ-lyase: Comprehensive kinetic analysis of the complex reaction with L-methionine. Anal. Biochem., 327: 233-240.
CrossRefDirect Link - Takakura, T., T. Ito, S. Yagi, Y. Notsu and T. Itakura et al., 2006. High-level expression and bulk crystallization of recombinant L-methionine γ-lyase, an anticancer agent. Applied Microbiol. Biotechnol., 70: 183-192.
CrossRefDirect Link - Tan, Y., X. Sun, M. Xu, X. Tan and A. Sasson et al., 1999. Efficacy of recombinant methioninase in combination with cisplatin on human colon tumors in nude mice. Clin. Cancer Res., 5: 2157-2163.
PubMedDirect Link - Tan, Y., M. Xu, X. Tan, X. Tan and X. Wang et al., 1997. Overexpression and large-scale production of recombinantl-methionine-α-deamino-γ-mercaptomethane-lyase for novel anticancer therapy. Protein Expr. Purif., 9: 233-245.
CrossRefDirect Link - Tan, Y., M. Xu, H. Guo, X. Sun, T. Kubota and R.M. Hoffman, 1996. Anticancer efficacy of methioninase in vivo. Anticancer Res., 16: 3931-3936.
Direct Link - Tanaka, H., N. Esaki and K. Soda, 1977. Properties of L-methionine γ-lyase from Pseudomonas ovalis. Biochemistry, 16: 100-106.
CrossRefDirect Link - Tanaka, H., N. Esaki, T. Yamamoto and K. Soda, 1976. Purification and properties of methioninase from Pseudomonas ovalis. FEBS Lett., 66: 307-311.
CrossRefDirect Link - Thomas, T. and T.J. Thomas, 2003. Polyamine metabolism and cancer. J. Cell. Mol. Med., 7: 113-126.
CrossRefDirect Link - Towbin, H., T. Staehelin and J. Gordon, 1979. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications. Proc. Natl. Acad. Sci. USA., 76: 4350-4354.
PubMedDirect Link - Van Rite, B.D., Y.A. Lazrak, M.L. Pagnon, N.R. Palwai, L.F. Neves, P.S. McFetridge and R.G. Harrison, 2011. Enzyme prodrug therapy designed to target L-methioninase to the tumor vasculature. Cancer Lett., 301: 177-184.
CrossRefDirect Link - Yang, Z., J. Wang, Q. Lu, J. Xu and Y. Kobayashi et al., 2004. PEGylation confers greatly extended half-life and attenuated immunogenicity to recombinant methioninase in primates. Cancer Res., 64: 6673-6678.
CrossRefDirect Link - Yano, S., S. Li, Q. Han, Y. Tan, M. Bouvet, T. Fujiwara and R.M. Hoffman, 2014. Selective methioninase-induced trap of cancer cells in S/G2 phase visualized by FUCCI imaging confers chemosensitivity. Oncotarget, 5: 8729-8736.
CrossRefPubMedDirect Link - Yoshioka, T., T. Wada, N. Uchida, H. Maki and H. Yoshida et al., 1998. Anticancer efficacy in vivo and in vitro, synergy with 5-fluorouracil and safety of recombinant methioninase. Cancer Res., 58: 2583-2587.
PubMedDirect Link - Zhao, R., F.E. Domann and W. Zhong, 2006. Apoptosis induced by selenomethionine and methioninase is superoxide mediated and p53 dependent in human prostate cancer cells. Mol. Cancer Ther., 5: 3275-3284.
CrossRefPubMedDirect Link