
Berna Elya
Faculty of Pharmacy, University of Indonesia, Postal Code, 16424, Depok, Indonesia
Rosita Handayani
Faculty of Pharmacy, University of Indonesia, Postal Code, 16424, Depok, Indonesia
Rani Sauriasari
Faculty of Pharmacy, University of Indonesia, Postal Code, 16424, Depok, Indonesia
Azizahwati
Faculty of Pharmacy, University of Indonesia, Postal Code, 16424, Depok, Indonesia
Uqie Shabrina Hasyyati
Faculty of Pharmacy, University of Indonesia, Postal Code, 16424, Depok, Indonesia
Idam Titis Permana
Faculty of Pharmacy, University of Indonesia, Postal Code, 16424, Depok, Indonesia
Yunita Indah Permatasari
Faculty of Pharmacy, University of Indonesia, Postal Code, 16424, Depok, Indonesia
Pakistan Journal of Biological Sciences
Year: 2015 | Volume: 18 | Issue: 6 | Page No.: 279-284







ABSTRACT
Active compounds of plants have potency as antidiabetic that can be used in diabetes mellitus treatment. One of the strategies is maintaining postprandial glucose level through inhibition of α-amylase and α-glucosidase and preventing the hydrolysis of Glucagon Like Peptide-1 (GLP-1) through inhibition of Dipeptidyl peptidase IV. So that pre-prandial and post-prandial glucose levels can be controlled properly. The aim of this study was to determine in vitro inhibitory activity of α-glucosidase, α-amylase and Dipeptidyl peptidase IV from 42 ethanolic extracts of selected Indonesian plants. Inhibitory activity was measured using spectrophotometric method. α-amylase activity was measured at λ = 540 nm, α-glucosidase activity and the DPP-IV activity were measured at λ = 405 nm. Eight extracts inhibit both α-glucosidase and α-amylase better than acarbose as a positive control. These extracts are Camelia sinensis (white, green, fermented), Pometia pinnata, Syzygium polyanthum, Artocarpus heteropyllus, Lagerstroemia speciosa and Persea Americana. The ethanolic extracts have DPP-IV inhibitory activity between 10-20%, lower than Diprotin as positive control, except Camelia sinensis (white) which gave the inhibition more than 30% at 62.5 μg mL–1. This study showed us that some extracts have inhibition activity of α-glucosidase, α-amylase and DPP-IV.
PDF Abstract XML References Citation
Received: June 06, 2015;
Accepted: September 09, 2015;
Published: October 14, 2015
How to cite this article
Berna Elya, Rosita Handayani, Rani Sauriasari, Azizahwati, Uqie Shabrina Hasyyati, Idam Titis Permana and Yunita Indah Permatasari, 2015. Antidiabetic Activity and Phytochemical Screening of Extracts from Indonesian Plants by Inhibition of Alpha Amylase, Alpha Glucosidase and Dipeptidyl Peptidase IV. Pakistan Journal of Biological Sciences, 18: 279-284.
DOI: 10.3923/pjbs.2015.279.284
URL: https://scialert.net/abstract/?doi=pjbs.2015.279.284
DOI: 10.3923/pjbs.2015.279.284
URL: https://scialert.net/abstract/?doi=pjbs.2015.279.284
INTRODUCTION
Research of active compounds that have antidiabetic activity should be developed in order to obtain better therapeutic outcomes. There are some mechanisms of antidiabetic drugs, such as suppress hepatic glucose production (biguanides), stimulate insulin secretion (sulfonylureas and glinides), delay digestion and absorption of intestinal carbohydrates to maintain postprandial glucose level (α-glucosidase and α-amylase inhibitors), improve the sensitivity of insulin receptor and peripheral glucose uptake (thiazolidinediones and metformin) or insulin (Krentz and Bailey, 2005; Verspohl, 2009). One therapeutic approach for treating diabetes is decreasing post-prandial hyperglycemia by delaying glucose absorption through carbohydrate-hydrolyzing enzymes inhibition, α-glucosidase and α-amylase in the digestive tract. Inhibitors of these enzyme delay carbohydrate digestion and prolong overall carbohydrate digestion time, causing a reduction in the rate of glucose absorption and consequently blunting the postprandial plasma glucose rise (Rhabasa-Lhoret and Chiasson, 2004; Bhandari et al., 2008; Ban et al., 2009). Recently, considerable interest has been generated by a novel class of antihyperglycemic agents that act at distinct levels of the incretin pathway. Incretins are a group of gastrointestinal hormones, predominantly Glucagon Like Peptide-1 (GLP-1) and Gastric Inhibitory Polypeptide (GIP), which increase post-prandial insulin release from pancreatic b-cells in a glucose-dependent manner (Ban et al., 2009). About 50-60% of the total insulin secreted during a meal results from GLP-1 and GIP (Creutzfeldt, 2005). Due to rapid cleavage and inactivation, a therapy with native GLP-1 administered parenterally is not feasible for the continuous treatment of type 2 diabetes and thus incretin mimetics that are resistant to cleavage by DPP-IV are being pursued (Thornberry and Gallwitz, 2009). Dipeptidyl peptidase IV (DPP-IV) is a serine protease that localizes on cell surfaces. It is responsible for the rapid degradation of incretins such as GLP-1 and GIP (Barnett, 2006). Inhibition of dipeptidyl peptidase IV (DPP-IV) is being used as a therapeutic strategy in the treatment of Type 2 diabetes (Drucker, 2006; Lacroix and Li-Chan, 2014; Nongonierma et al., 2013; McIntosh, 2008; McIntosh et al., 2006). This study was conducted to find inhibitory activity of α-amylase, α-glucosidase and DPP-IV from Indonesian plants. By inhibiting α-amylase, α-glucosidase and DPP-IV, glucose levels can be controlled to maintain normal blood sugar level.
MATERIALS AND METHODS
Materials: α-glucosidase 0.15 U mL–1 from Saccharomyces cerevisiae (Sigma Aldrich, Singapore), Potassium dihydrogen phosphate 2 M (Merck) pH 6.8, substrate 4-Nitrophenyl-α-D glucopyranoside (PNPG) 5 mM (Sigma Aldrich, Singapore), Sodium carbonate 200 mM (Merck), Acarbose (Sigma Aldrich, Singapore), α-amylase 1 U mL–1 from Porcine kidney (Sigma Aldrich, Singapore), Soluble starch 1 and 2% (Merck), buffer Sodium dihydrogen phosphate 20 mM pH 6.9, Sodium chloride 6.7 mM, 3.5-Dinitro salicylic acid 96 mM (Sigma Aldrich, Singapore), Sodium hydroxide 2 M, Sodium potassium tartrate (Merck), standard of maltose 0.2%, Dipeptidyl peptidase IV (DPP-IV) from porcine kidney (Sigma Aldrich, Singapore), substrate Gly-Pro-P-Nitroanilide (GPPN) (Sigma Aldrich, Singapore), Diprotin-A (Ile-Pro-Ile) (Sigma Aldrich, Singapore).
METHODS
Extraction: The powder of the dried plants (150 g) were extracted with 300 mL ethanol 70% using reflux method. The obtained extracts were evaporated with a rotary evaporator.
Phytochemistry screening: The viscous extracts (50 mg) were diluted using ethanol. Identification of the presence of alkaloids, flavonoids, tannins, saponins, glycosides, terpenoids and anthraquinone conducted in accordance with procedures of Indonesian Materia Medika and Harborne (MoH., 1995; Harborne, 1987).
α-glucosidase activity assay: A 36 μL of phosphate buffer solution, 30 mL sample solution with various concentrations (10, 25, 50, 100 and 150 μg mL–1) and 17 μL PNPG substrate at concentration of 5 mM were put in 96 well microplate. This mixture was incubated at 37°C for 5 min. After 5 min, 17 μL of α-glucosidase solution 0.15 U mL–1 was added in each well to obtain total volume of 100 mL. The mixture was incubated for 15 min to get the complete hydrolysis reaction. After 15 min, the reaction was stopped by adding 100 μL of Sodium Carbonate (Na2CO3) 200 mM. Absorbance was measured at 405 nm using a microplate reader. Each test was repeated twice (Sigma-Aldrich, 1995; Dewi et al., 2007).
Calculation of percent inhibition: The obtained data from the activity test was processed using the following formula in Eq. 1:
| (1) |
| A1 | : | Blanko absorbance (B*)-control of blanko absorbance (KB**) |
| A2 | : | Sample absorbance (S)-control of sample absorbance (KS***) |
| * | : | Blanko contains substrate + enzyme, without extract |
| ** | : | Control of blanko contains substrate and buffer, without enzyme and extract |
| *** | : | Control of sample contains substrate + extract with the addition of the enzyme after incubation |
An experiment used 5 variation of sample concentration. Percent inhibition obtained in each sample was processed in form of graph and compared to the percent inhibition of acarbose as a positive control.
Calculation of IC50: The IC50 was calculated using linear regression equation in which the concentration of the sample as the x-axis and percent inhibition as the y-axis. From the equation y = a+bx, IC50 values can be calculated using the following formula in Eq. 2:
| (2) |
α-amylase activity assay: A 250 μL of 500 μg mL–1extract, 250 μL of starch 2.0% (w/v) and 250 μL of 1 U mL–1 α-amylase solution were homogeneously mixed into a test tube. After incubated at 20°C for 3 min, 500 μL of color reagent (Dinitrosalicylic acid) was added to stop the enzymatic reaction. The mixture was put into boiled water and 250 μL α-amylase 1 U mL–1 was added immediately. Heating process was continued for 15 min. Then the solution was removed from heating process and cooled at room temperature for 3 min. A 4500 μL aqua dest was added to obtain total volume of 6000 μL. The solution was homogenized using a vortex. The α-amylase activity was determined at 540 nm using spectrophotometry to measure product absorbance (maltose) which reduces DNS. The produced absorbance was compared with blank. Percent inhibition was calculated using the Eq. 1 (Sigma-Aldrich, 2014; Ali et al., 2006).
DPP IV activity assay: A 25 μL extract was added with 50 μL DPP-IV (500 μg mL–1). The mixture was incubated at 37°C for 5 min. A 100 μL GPPN (2 mM) was added to the wells containing extract and enzyme. Incubation was continued for 15 min. The reaction was terminated by adding 25 μL acetic acid glacial 25%. The absorbance was measured at λ = 405 nm (Chakrabarti et al., 2011).
RESULTS
Phytochemistry screening: Phytochemistry screening conducted on 7 secondary metabolites presence: alkaloids, flavonoids, tannins, glycosides, saponin, terpene/steroids and anthraquinone. There was 42 samples tested, 40 plants contain alkaloids, flavonoids and glycosides (Table 1).
Enzymatic activity assay: The test results of α-glucosidase presented with IC50 values of samples using Eq. 2, while for the testing of α-amylase and DPP-IV test, data is shown only through the percent inhibition values were calculated based on Eq. 1.
| Table 1: | Phytochemistry screening result |
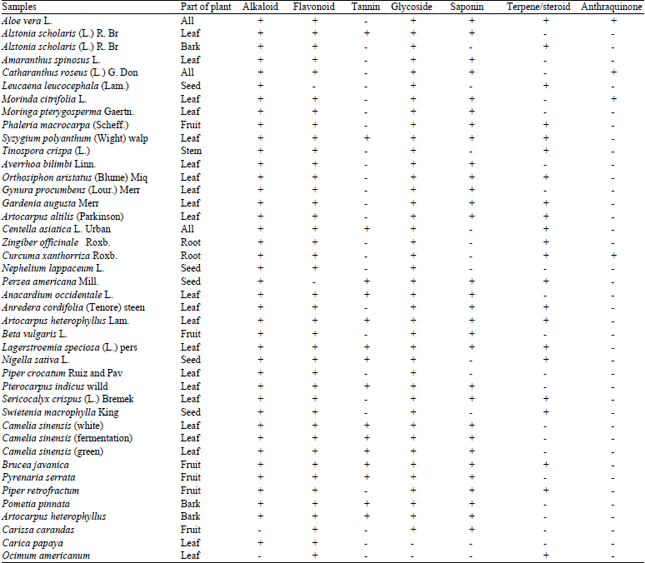 | |
This was done because the α-glucosidase inhibition was linear (extract concentration is proportional to the percent inhibition produced), while the α-amylase and DPP-IV inhibition properties are non-linear, the increase of the extract concentration does not increase the percent inhibition. Therefore, the inhibition of α-amylase and the DPP-IV can not be analyzed by using linear regression equation (Eq. 2).
DISCUSSION
In vitro α-glucosidase inhibitory studies demonstrated that all samples had inhibitory activity (Table 2). The percentage inhibition at 10, 25, 50, 100 and 150 μg mL–1 concentrations of samples showed a concentration dependent on percentage inhibition. The highest inhibitory activity of samples was obtained from Camelia sinensis (white) extracts with IC50 value 10.54 μg mL–1. The IC50 values of Camelia sinensis (white) were much smaller than the IC50 of Acarbose as a positive control (IC50 Acarbose = 36.98 μg mL–1). It shows that Camelia sinensis (white) inhibits the activity of α-glucosidase better than acarbose in a smaller concentration. Eight other samples also showed better results than Acarbose, which are Camelia sinensis (white tea), Camelia sinensis (green tea), Camelia sinensis(with fermentation), Pometia pinnata, Syzygium polyanthum, Artocarpus heterophyllus, Lagerstroemia speciosa and Persea Americana.
| Table 2: | Assay of enzyme inhibition activity |
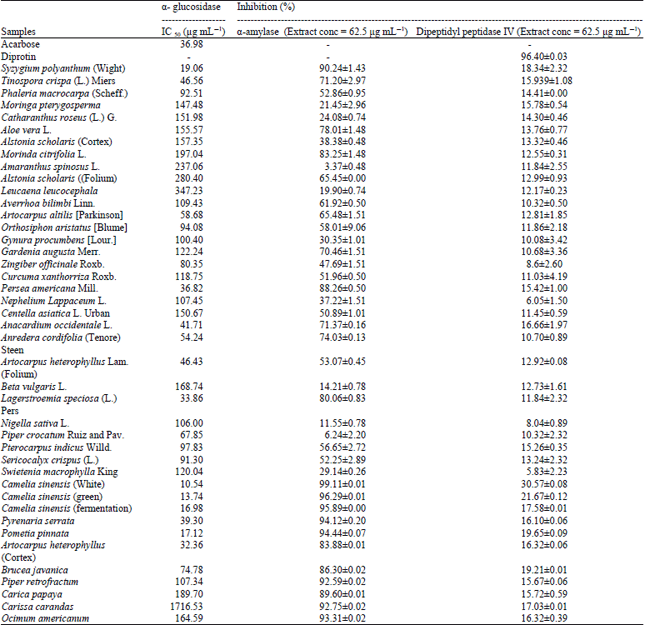 | |
All of them contain tannin. In recent years, tannin has been reported as non-specific inhibitors for several hydrolytic enzymes such as lipases, α-glucosidases, α-amylases and invertase (Barret et al., 2013; Goncalves et al., 2011; Grussu et al., 2011; Ikarashi et al., 2011; Kusano et al., 2011; Yilmazer-Musa et al., 2012; Zajacz et al., 2007). This hypothesis can be proved by further experiments such as tannin isolation and activity assay of isolate that have been separated from other compounds.
In α-amylase in vitro study, all extracts showed inhibition value between 3-99%. There were 14 samples that have inhibition values more than 80%. All of them contain glycosides. The glycosides present in the crude extracts acts as a substrate for the α-glucosidase enzyme and may be responsible for the inhibitory activity (Elya et al., 2012). Amylases degrade starch by cleaving glycosidic bonds. Glycoside was found from the sample has glycosidic bonds so it’s changing the role of starch as a substrate. With this mechanism, the starch in the body was not changed to form a disaccharide. It could help the work of glucosidase which converts disaccharide into monosaccharides (glucose) and the level of glucose can be controlled.
In DPP-IV in vitro activity assay, inhibition activity of extracts were relatively lower compared to Diprotin as positive control. In total concentration of 62.5 mg mL–1, diprotin inhibited DPP-IV with percent inhibition is 96.40±0.03%, while the plant extracts inhibited DPP-IV with percent inhibition 10-20%. Camelia sinensis (white tea) showed the highest inhibition of all extracts in this study with percent inhibition is 30.57±0.08%.
Different from alpha glucosidase, inhibition activity of alpha amylase and DPP-IV were not dependent on concentration. IC50 calculations on the inhibition of α-amylase and DPP-IV are not done because the extract has a non-linear inhibition profile.
Our in vitro studies demonstrated an appreciable α-glucosidase, α-amylase and DPP-IV inhibitory activity present in 42 samples, where further experiments can be performed on animal models to confirm the hypoglycemic activity.
CONCLUSION
All of the tested extracts show inhibition activity of α-glucosidase, α-amylase and DPP-IV. Camelia sinensis (white), Camelia sinensis (green), Camelia sinensis (with fermentation), Pometia pinnata, Syzygium polyanthum, Artocarpus heterophyllus, Lagerstroemia speciosa, dan Persea Americana have better inhibition activity of α-glucosidase than Acarbose in concentration 62.5 μg mL–1. Plant extract which contain tannin (polyphenol) and glycoside have better inhibition activity of α-amylase in concentration 62.5 μg mL–1. All of the tested extracts showed inhibition activity, but they have shown relatively lower inhibition of DPP-IV than Diprotin in concentration 62.5 μg mL–1.
ACKNOWLEDGMENT
This work was supported by Hibah Grant Cluster University of Indonesia.
REFERENCES
- Ali, H., P.J. Houghton and A. Soumyanath, 2006. α-Amylase inhibitory activity of some Malaysian plants used to treat diabetes; with particular reference to Phyllanthus amarus. J. Ethnopharmacol., 107: 449-455.
CrossRefDirect Link - Ban, K., S. Hui, D.J. Drucker and M. Husain, 2009. Cardiovascular consequences of drugs used for the treatment of diabetes: Potential promise of incretin-based therapies. J. Am. Soc. Hyperten., 3: 245-259.
CrossRefDirect Link - Barret, A., T. Ndou, C.A. Hughey, C. Straut, A. Howell, Z. Dai and G. Kaletunc, 2013. Inhibition of α-amylase and glucoamylase by tannins extracted from cocoa, pomegranates, cranberries and grapes. J. Agric. Food Chem., 61: 1477-1486.
CrossRefDirect Link - Barnett, A., 2006. DPP-4 inhibitors and their potential role in the management of type 2 diabetes. Int. J. Clin. Pract., 60: 1454-1470.
CrossRefPubMedDirect Link - Bhandari, M.R., N. Jong-Anurakkun, G. Hong and J. Kawabata, 2008. α-Glucosidase and α-amylase inhibitory activities of nepalese medicinal herb. Food Chem., 106: 247-252.
CrossRef - Chakrabarti, R., S. Bhavtaran, P. Narendra and N. Varghese, 2011. Dipeptidyl Peptidase-IV inhibitory activity of Berberis aristata. J. Natural Prod., 4: 158-163.
Direct Link - Creutzfeldt, W., 2005. The [pre-] history of the incretin concept. Regulatory Peptides, 128: 87-91.
CrossRefDirect Link - Dewi, R.T., Y.M. Iskandar, M. Hanafi, L.B.S. Kardono, M. Angelina, I.D. Dewijanti and S.D.S. Banjarnahor, 2007. Inhibitory effect of Koji Aspergillus terreus on α-glucosidase activity and postprandial hyperglycemia. Pak. J. Biol. Sci., 10: 3131-3135.
CrossRefPubMedDirect Link - Drucker, D.J., 2006. Enhancing the action of incretin hormones: A new whey forward. Endocrinology, 147: 3171-3172.
Direct Link - Goncalves, R., N. Mateus and V. de Freitas, 2011. Inhibition of α-amylase activity by condensed tannins. J. Food Chem., 125: 665-672.
CrossRefDirect Link - Grussu, D., D. Stewart and G.J. McDougall, 2011. Berry polyphenols inhibit α-amylase in vitro: Identifying active components in rowanberry and raspberry. J. Agric. Food Chem., 59: 2324-2331.
CrossRefDirect Link - Ikarashi, N., R. Takeda, K. Ito, W. Ochiai and K. Sugiyama, 2011. The inhibition of lipase and glucosidase activities by acacia polyphenol. Evidence-Based Complement. Alternative Med.
CrossRefDirect Link - Krentz, A.J. and C.J. Bailey, 2005. Oral antidiabetic agents: Current role in type 2 diabetes mellitus. Drugs, 65: 385-411.
PubMedDirect Link - Kusano, R., S. Ogawa, Y. Matsuo, T. Tanaka, Y. Yazaki and I. Kouno, 2011. α-amylase and lipase inhibitory activity and structural characterization of acacia bark proanthocyanidins. J. Nat. Prod., 74: 119-128.
CrossRefDirect Link - Lacroix, I.M.E. and E.C.Y. Li-Chan, 2014. Overview of food products and dietary constituents with antidiabetic properties and their putative mechanisms of action: A natural approach to complement pharmacotherapy in the management of diabetes. Mol. Nutr. Food Res., 58: 61-78.
CrossRefDirect Link - McIntosh, C.H.S., H.U. Demuth, S.J. Kim, J.A. Pospisilik and R.A. Pederson, 2006. Applications of dipeptidyl peptidase IV inhibitors in diabetes mellitus. Int. J. Biochem. Cell Biol., 38: 860-872.
CrossRefDirect Link - McIntosh, C.H.S., 2008. Incretin-based therapies for type 2 diabetes. Can. J. Diabetes, 32: 131-139.
CrossRefDirect Link - Nongonierma, A.B., C. Mooney, D.C. Shields and R.J. Fitzgerald, 2013. Inhibition of dipeptidyl peptidase IV and xanthine oxidase by amino acids and dipeptides. Food Chem., 141: 644-653.
CrossRefDirect Link - Thornberry, N.A. and B. Gallwitz, 2009. Mechanism of action of inhibitors of dipeptidyl-peptidase-4 (DPP-4). Best Pract. Res. Clin. Endocrinol. Metab., 23: 479-486.
CrossRefDirect Link - Yilmazer-Musa, M., A.M. Griffith, A.J. Michels, E. Schneider and B. Frei, 2012. Grape seed and tea extracts and catechin 3-gallates are potent inhibitors of α-amylase and α-glucosidase activity. J. Agric. Food Chem., 60: 8924-8926.
CrossRefDirect Link - Zajacz, A., G. Gyemant, N. Vittori and L. Kandra, 2007. Aleppo tannin: Structural analysis and salivary amylase inhibition. Carbohyd. Res., 342: 717-723.
CrossRefDirect Link - Elya, B., K. Basah, A. Mun'im, W. Yuliastuti, A. Bangun and E.K. Septiana, 2012. Screening of α-glucosidase inhibitory activity from some plants of Apocynaceae, Clusiaceae, Euphorbiaceae and Rubiaceae. J. Biomed. Biotechnol.
CrossRefDirect Link