
Naveen Kumar
National Institute of High Security Animal Diseases, Bhopal, Madhya Pradesh, India
Yashpal Singh Malik
Division of Biological Standardization, Indian Veterinary Research Institute (IVRI), Izatnagar, 243122, Uttar Pradesh, India
Kuldeep Sharma
Division of Biological Standardization, Indian Veterinary Research Institute (IVRI), Izatnagar, 243122, Uttar Pradesh, India
Vinayagamurthy Balamurugan
Animal Disease Monitoring and Surveillance, H A Farm, Hebbal, Bengaluru, 560024, Karnataka, India
Sathish Bhadravati Shivachandra
Division of Virology, Indian Veterinary Research Institute (IVRI), Mukteswar, 263138, Nainital, Uttarakhand, India
Kuldeep Dhama
Division of Pathology, Indian Veterinary Research Institute (IVRI), Izatnagar, 243122, Uttar Pradesh, India
Pakistan Journal of Biological Sciences
Year: 2015 | Volume: 18 | Issue: 5 | Page No.: 247-251







ABSTRACT
Rotaviruses of group A (RVA) are foremost cause of diarrhoeal diseases in neonates of animals and humans worldwide leading to substantial economic losses. The RVA non-structural protein-4 (NSP-4), a viral enterotoxin, is known to be associated with infantile gastroenteritis/secretory diarrhoea by inducing pathological changes in the mature enterocytes. In this study, the carboxyl terminus of NSP4 protein (73M to 175M) from a bovine RVA was expressed in Escherichia coli Tuner (DE3) pLysS cells. The fusion protein (rNSP4ct, ~31 kDa) with hexa-histidine tags on its both termini was purified by affinity chromatography under native condition using Nickel-Nitrilotriacetic acid (Ni-NTA) agarose resin. The purified soluble recombinant NSP4ct was confirmed by Western blot. The structural analysis of rNSP4 protein revealed similarity between bovine RVA and human RVA (central tetrameric coiled-coil region) and confirmed that it was composed of mainly alpha helix (85%), lacking the beta strands. The rNSP4ct protein of bovine RVA has the potential of being used in developing diagnostics, assessing the biological activity (enterotoxin property) of rNSP4ct in understanding the pathogenesis in intestinal mucosa which would reveal the role of anti-NSP4 antibodies in protection against rotavirus infection and stimulation of mucosal immunity in animal model.
PDF Abstract XML References Citation
Received: June 22, 2015;
Accepted: August 15, 2015;
Published: August 27, 2015
How to cite this article
Naveen Kumar, Yashpal Singh Malik, Kuldeep Sharma, Vinayagamurthy Balamurugan, Sathish Bhadravati Shivachandra and Kuldeep Dhama, 2015. Expression Analysis of a Highly Soluble Region of an Enterotoxigenic Non-structural Protein of Bovine Rotavirus. Pakistan Journal of Biological Sciences, 18: 247-251.
DOI: 10.3923/pjbs.2015.247.251
URL: https://scialert.net/abstract/?doi=pjbs.2015.247.251
DOI: 10.3923/pjbs.2015.247.251
URL: https://scialert.net/abstract/?doi=pjbs.2015.247.251
INTRODUCTION
Rotaviruses of group A (RVA) are foremost cause of diarrhoeal diseases in neonates of animals and humans worldwide leading to considerable economic losses specifically in developing countries (Estes and Kapikian, 2007). Rotaviruses (RVs) belong to the family Reoviridae and possess a genome of 11 segments of double stranded RNA, encoding six structural (VP1-6) and six non-structural proteins (NSP1-6). The NSPs are known to play an essential and diverse role such as in virus replication, morphogenesis and pathogenesis and most of them exhibit multifunctional properties. The NSPs are also good targets for antivirals and development of diagnostics. The RVAs E2 genotype is among the most diverse E types detected in NSP4 gene, which correspond to the KUN-like (genotype A) and has been widely isolated from several species of animals including bovines (Matthijnssens et al., 2008). Identification of the NSPs of RVA, especially NSP4 as the first viral enterotoxin, has contributed significantly towards understanding of its structure and mechanism of action in inducing secretory diarrhoea in neonates (Malik et al., 2014).
NSP4 (175aa), encoded by genome segment 10, is a trans-membrane, Endoplasmic Reticulum (ER)-specific glycoprotein with pleotropic functions including viral morphogenesis and pathogenesis. It contains a signal sequence at the N-terminus followed by three hydrophobic domains namely H1 (7-21 aa), H2 (29-47 aa), H3 (67-85 aa) and a coiled α-helical domain (95-137 aa) (Estes and Kapikian, 2007). Its amino-terminal hydrophobic region (1-44 aa) is located in the lumen of the endoplasmic reticulum, whereas its cytoplasmic carboxyl-terminal region (45-175 aa) constitutes the Cytoplasmic Tail (CT) that exhibits all the known important biological properties associated with the protein. These include alteration of Ca2+ homeostasis by the release of Ca2+ from the endoplasmic reticulum, membrane permeabilization, Ca2+ and VP4-binding, double-layered particle-interaction for transport in the lumen of the endoplasmic reticulum for maturation into triple-layered particles and diarrhoea induction in newborn mouse pups. A secreted form of NSP4 containing residue 112-175 aa was known to possess diarrhoea-inducing ability similar to that of full-length protein (Zhang et al., 2000).
Though, full length NSP4 protein has been expressed in Lactococcus lactis (Enouf et al., 2001), insect cell lines (Rodriguez-Diaz et al., 2003), mammalian cell lines (Newton et al., 1997), but expression of full length NSP4 protein in prokaryotic system has not been successful because of presence of membrane-destabilizing domain on NSP4 protein (Enouf et al., 2001). Moreover, prokaryotic expression system was preferred in the production of large quantities of recombinant NSP4, as non-glycosylated NSP4 is also biologically active (Mori et al., 2002; Sharifi et al., 2005). Therefore, in this study, effort was made to express and purify the highly soluble recombinant carboxyl terminus of NSP4 protein of bovine RVA excluding the N-terminus membrane-destabilizing domain in E. coli.
MATERIALS AND METHODS
Vector and host cells: Four different strains {BL21 (DE3) pLysS, Origami (DE3) pLysS, Rosetta-gami (DE3) pLysS and Tuner (DE3) pLysS} of E. coli procured from M/s Novagen (Germany), were used for the expression of carboxyl terminus of NSP4 protein. For cloning, pET-32a (+) vector under the transcriptional control of the bacteriophage T7 promoter with lac operator was used. All the cells were grown in Luria Bertani (LB) broth (HiMedia, India) on shaker cum incubator at 37°C (180 rpm). Standard procedure for transformation was followed as per the manufacturer instructions. Overnight grown cultures in LB broth, supplemented with appropriate antibiotics, were diluted 1:10 in fresh LB broth supplemented with appropriate antibiotics. Cells were induced with (0.2-2 mM) isopropyl β-D-1-thiogalactopyranoside (IPTG) after 1-2 h incubation at 37°C (180 rpm) when optical density was approximately 0.6-0.8.
Cloning of partial NSP4 gene sequences: During the study, bovine RVA positive diarrhoeic faecal sample (C91) retrieved from Enteric Virus Laboratory, Indian Veterinary Research Institute, Mukteswar, India, was processed for viral RNA extraction using TriReagent-LS (Sigma, St. Louis, USA) followed by cDNA synthesis as described previously (Zhang et al., 2000; Matthijnssens et al., 2008; Malik et al., 2012). A primer set targeting carboxyl terminus of NSP4 protein (217-525 nt; 73-175 aa) of bovine RVA (genotype E2) was designed based on its available gene sequences (GenBank accession No. JN626229). The primers for gene amplification were synthesized (IDT-DNA, USA), which had added restriction enzyme sites (underlined) for EcoRI and XhoI at 5´ end of forward and reverse primer, respectively with primer tags (small letters) were as follows: NSP4ct[+]: 5´-GAACGAATTCATGGTAACAATTTTTAACACG-3´; NSP4ct[-]: 5´-TCAGCTCGAGCTTACATTGC TGCAG-3´.
The Reverse Transcriptase-Polymerase Chain Reaction (RT-PCR) reaction conditions were optimized and amplification resulted in an amplicon of approximately 308 bp as visualized in 2% agarose gel electrophoresis (data not shown). The amplified PCR product digested with EcoRI and XhoI, was gel eluted using QIAquick gel extraction kit (Qiagen, Germany) and ligated to a linearized pET32a (Novagen, USA). The resultant recombinant plasmid (pNSP4ct) was initially transformed in to E. coli TOP10 cells. The positive colonies containing recombinant plasmid (pNSP4ct) were confirmed by colony PCR and restriction enzyme analysis after isolation of plasmid DNA by using GeneJET plasmid miniprep kit (Fermentas, USA) and sequencing. Subsequently, plasmid (pNSP4ct) was transformed into four expression host cells of E. coli, as mentioned under vector and host cells.
Expression and purification of truncated NSP4 protein under native conditions: For expression, recombinant cells were grown in LB broth with 1% glucose containing antibiotics [ampicillin (50 μg mL–1) and chloramphenicol (35 μg mL–1)] and kept in shaking incubator at temperature ranges (25-37°C) before induction with (0.2-2 mM) IPTG (Sigma-Aldrich, USA). After 3 h post induction, cells were harvested by centrifugation at 4025×g for 20 min and cell pellet was re-suspended in a BugBuster protein extraction reagent (Novagen, Germany) as per the manufacturer’s instructions for cell lysis. Following cell lysis, lysate was centrifuged at 28,621×g for 20 min at 4°C to get the clear supernatant containing soluble recombinant NSP4ct protein. The recombinant NSP4ct protein was purified by affinity chromatography using Ni-NTA agarose resin (Qiagen, Germany) as per the standard manufacturer’s protocol. The soluble recombinant truncated NSP4 protein after purification was dialyzed by using cellulose membrane (Mol wt. 12 kDa cut off) (Sigma, USA) overnight at 4°C with phosphate-buffered saline (pH 7.4) and concentrated by polyethylene glycol-6000. The purified protein was quantified using Nanodrop-1000 Spectrophotometer (Thermo-Scientific, USA).
Characterization and in-silico secondary structure prediction: For detection, recombinant NSP4ct protein with hexa-histidine tag was transferred on to the polyvinylidene difluoride (PVDF) membrane (MDI, India) using, semi-dry immunoblot system (Amersham pharmacia, USA). The recombinant protein was detected by chromogenic method using monoclonal anti-polyhistidine peroxidase conjugate (Sigma, USA) diluted at the rate of 1:1000 in 3% bovine serum albumin and 3,3’-diaminobenzidine (Sigma, USA) as the substrate. The NSP4 gene of bovine RVA genotype E2 (GenBank accession No. JN626229) was used for prediction of secondary structure for truncated NSP4 using Phyre 2.0 (Kelley and Sternberg, 2009).
RESULTS
Optimization of RT-PCR: The RT-PCR reaction conditions optimized with denaturation at 95°C for 5 min followed by cyclic denaturation at 94°C for 30 sec, annealing at 54°C for 20 sec and extension at 72°C for 30 sec for repeated 30 cycles and a final extension at 72°C for 5 min yielded specific amplicon of 308 bp cytoplasmic tail region (217-525 nt) of NSP4 gene of bovine RVA. The PCR product was visualized after resolving in ethidium bromide containing 2% agarose gel.
Expression and purification of truncated NSP4 protein: It was established after optimization that when Tuner (DE3) pLysS cells were induced at an optical density of 0.6-0.8, significant improvement in expression resulted at IPTG (1 mM), incubation temperature post-induction (28°C) and length of time cultures grown following induction (3 h). Following induction with 1 mM IPTG, the recombinant E. coli expressed a NSP 4ct protein (12 kDa, 104 aa) along with hexa-histidine tags region (19 kDa, 175 aa) on its both N- and C-terminus, accounting for a total molecular weight of ~31 kDa (279 aa) as observed on 10% SDS-PAGE (Fig. 1, Panel-D and Lane 2). Solubility analysis of over-expressed NSP4ct protein following cell lysis indicated the presence of high amount of expressed protein in soluble fraction. Further, the rNSP4 protein was purified under native condition though elution and dialysis, which resulted in purified single band of recombinant NSP4ct protein on 10% SDS-PAGE at expected size of approximately 31 kDa (Fig. 1, Panel-D and Lane 3). Immunoblot using anti-His antibodies detected the presence of hexa-histidine tag in recombinant NSP4ct protein by development of brown color in a chromogenic reaction on PVDF membrane (Fig. 1, Panel-D and Lane 4).
In-silico secondary structure prediction: On prediction, recombinant truncated NSP4 protein composed of mainly alpha helix (85%) and lack beta strand. The various biological important domains within truncated NSP4 protein are portrayed in Fig. 1, Panel C. A protein model was also constructed using this web based server which is depicted in Fig. 1, Panel E. Structurally, the NSP4 of bovine RVA was similar to that of human RVA partially (central tetrameric coiled-coil region) solved NSP4 structure (Chacko et al., 2011). Although, it was noted that it forms a tetramer by a two fold rotation of dimer having two chains, there is a still considerable debate on its oligomeric state and respective biological activities. Hence, there is a need to express, purify and crystallize the NSP4 of different animal origin to elucidate their role in pathogenesis. However, studies on solving the complete NSP4 structure were mired by inability to express full length NSP4 in heterologous expression systems either due to poor expression, aggregation and/or insolubility.
DISCUSSION
This study reports for the first time expression and purification of carboxyl terminus of NSP4 protein of bovine RVA in E. coli. The identification of biological and enterotoxigenic properties in NSP4 protein of RVs has created interest in scientific community to understand its structure and mechanism of action in inducing secretory diarrhea in suckling mice. This 175 amino acids containing protein is responsible for different biological activities mapped to different regions (Browne et al., 2000).
The NSP4 proteins being glycosylated in nature, with two potential N-glycosylation sites in all mammalian strains, it is suggested to express in eukaryotic expression system like Sf9 cells (Rodriguez-Diaz et al., 2003). Nonetheless, expression of full length NSP4 in Sf9 cells was found to be associated with increased intracellular calcium levels (Tian et al., 1995). Surprisingly, the expression of full length NSP4 did not influence the permeability of the plasma membrane in Sf9 cells, but changed plasma membrane permeability in mammalian cells (Browne et al., 2000; Newton et al., 1997; Tian et al., 1996). Though, very high level expression of NSP4 was linked with cytotoxicity in Sf9 cells, levels typically detected in rotavirus-infected cells did not affect cell viability (Tian et al., 1996).
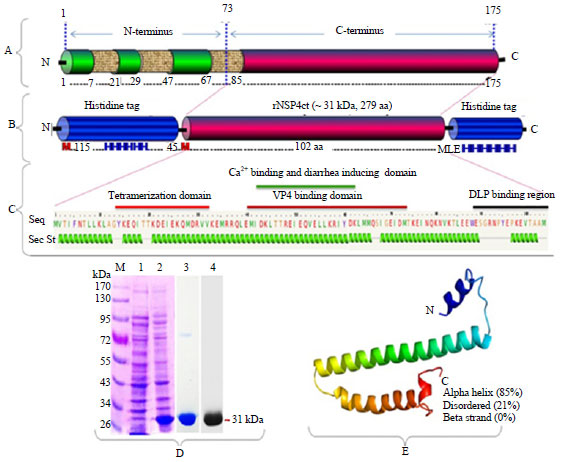 | |
| Fig. 1: | Design of construct, expression, purification and Western blot of recombinant partial NSP4 protein of bovine rotavirus, Panel A: Full length NSP4 protein of bovine rotavirus genotype E2, Panel B: Carboxyl terminus of NSP4 protein with hexa-histidine tags at N- and C-terminus, Panel C: Protein sequence of NSP4ct region with predicted secondary structure and domains, Panel D: Analysis of truncated NSP4 protein expression, purification and Western blot, Lanes-M: PageRuler™ Prestained Protein Ladder (MBI Fermentas), Lane 1: E. coli cell lysate before induction, Lane 2: E. coli cell lysate after induction with 1 M IPTG showing rNSP4ct (31 kDa), Lane 3: Purified rNSP4ct protein by affinity chromatography using Ni-NTA agarose resin, Lane 4: Immunoblot using monoclonal anti-polyHistidine peroxidase conjugate and Panel E: Three dimensional model of NSP4ct region, N: Amino terminus, C: Carboxyl terminus, Numericals in Panel A and B indicate position of amino acids in NSP4, Seq: Sequence of NSP4ct, SecSt: Secondary structure |
However, purified NSP4 protein expressed in E. coli has also shown to possess enterotoxigenic activity in sucking mice (Mori et al., 2002).
In view of known fact that N-terminal hydrophobic domain and membrane destabilizing domain located within the residues 55-69 aa of NSP4 protein, the expression becomes difficult owing to cytotoxic effect in E. coli and mammalian cells (Browne et al., 2000; Newton et al., 1997). Therefore, expression primers were designed to amplify the carboxyl terminal region (73-175 aa) having all the important biological properties of protein. The prokaryotic expression system was preferred because of easy availability, low cost, high-productivity, well-characterized genetics and ability of the bacterial host to accumulate recombinant proteins up to 80% of its dry weight. The partial viral enterotoxin (NSP4) protein of bovine RVA was successfully expressed in E. coli with majority of it in the soluble fraction of cell lysate. The high solubility of rNSP4ct could be due to the highly hydrophilic α-helix regions of NSP4 coupled with soluble stretch of amino acids provided by pET32a vector. Hence, the recombinant NSP4ct protein was purified under native conditions (Fig. 1, Panel C and Lane 3). Previously, many researchers have noticed the positive effect of the hydrophilic nature of the histidine tag on enhanced expression and high solubility of expressed recombinant fusion proteins (Ahuja et al., 2012). It is also presumed that the native conformation of NSP4 was preserved as it was held up on both sides by additional residues along with histidine regions that could have prevented it from forming an oligomer or tetramer (Chacko et al., 2011). A total yield of approximately 36 mg L–1 was obtained under described purification conditions. The purified rNSP4ct protein was devoid of non-specific proteolytic degradation and aggregations. The results showed that Tuner (DE3) pLysS expression host cells are suitable for the production of high quantity of rNSP4ct protein of bovine RVA, due to their intrinsic capability for higher turn over which might be associated with the lac permease mutation that probably allowed the uniform entry of IPTG inside the cells to give maximum expression.
CONCLUSION
Conclusively, the study indicated the higher expression and solubility of rNSP4ct protein due to the presence of hydrophilic histidine tags on both termini. The recombinant NSP4ct protein produced in this study has multiple applications viz., (1) It could be used in crystallization studies to understand the oligomeric state and structural features in relation to pathogenesis, (2) It could be potentially used in the development of rapid diagnostic assay for detecting coproantibodies in bovines and (3) It could also be used in assessing the biological activity (enterotoxin property) of rNSP4ct in understanding the pathogenesis in intestinal mucosa, which would reveal the role of anti-NSP4 antibodies in protection against rotavirus infection and stimulation of mucosal immunity in animal model.
ACKNOWLEDGMENTS
The authors wish to thank Director, Indian Veterinary Research Institute (IVRI), Izatnagar, India for the financial and infrastructural support. This work was supported by a grant (BT/168/NE/TBP/2011) to YPS from Department of Biotechnology, New Delhi, India and National Fellow project from Division of Education, Indian Council of Agricultural Research, Govt. of India, New Delhi.
REFERENCES
- Ahuja, A., A. Sen, R. Yogisharadhya, K.K. Rajak and S.B. Shivachandra, 2012. Prokaryotic expression and purification of highly soluble partial Glycoprotein Erns of Indian strain of classical swine fever virus. Indian J. Virol., 23: 397-401.
CrossRefDirect Link - Browne, E.P., A.R. Bellamy and J.A. Taylor, 2000. Membrane-destabilizing activity of rotavirus NSP4 is mediated by a membrane-proximal amphipathic domain. J. Gen. Virol., 81: 1955-1959.
Direct Link - Chacko, A.R., M. Arifullah, N.P. Sastri, J. Jeyakanthan and G. Ueno et al., 2011. Novel pentameric structure of the diarrhea-inducing region of the rotavirus enterotoxigenic protein NSP4. J. Virol., 85: 12721-12732.
CrossRefDirect Link - Enouf, V., P. Langella, J. Commissaire, J. Cohen and G. Corthier, 2001. Bovine rotavirus nonstructural protein 4 produced by Lactococcus lactis is antigenic and immunogenic. Applied Environ. Microbiol., 67: 1423-1428.
CrossRefDirect Link - Kelley, L.A. and M.J.E. Sternberg, 2009. Protein structure prediction on the web: A case study using the Phyre server. Nat. Protocols, 4: 363-371.
CrossRefDirect Link - Malik, Y.S., K. Sharma, N. Vaid, S. Chakravarti and K.M. Chandrashekar et al., 2012. Frequency of group A rotavirus with mixed G and P genotypes in bovines: Predominance of G3 genotype and its emergence in combination with G8/G10 types. J. Vet. Sci., 13: 271-278.
CrossRefPubMedDirect Link - Malik, Y.S., N. Kumar, K. Sharma, S. Ghosh and K. Banyaic et al., 2014. Molecular analysis of non structural rotavirus group A enterotoxin gene of bovine origin from India. Infect. Genet. Evol., 25: 20-27.
CrossRefPubMedDirect Link - Matthijnssens, J., M. Ciarlet, E. Heiman, I. Arijs and T. Delbeke et al., 2008. Full genome-based classification of rotaviruses reveals a common origin between human Wa-Like and porcine rotavirus strains and human DS-1-like and bovine rotavirus strains. J. Virol., 82: 3204-3219.
CrossRefDirect Link - Mori, Y., M.A. Borgan, N. Ito, M. Sugiyama and N. Minamoto, 2002. Sequential analysis of nonstructural protein NSP4s derived from Group A avian rotaviruses. Virus Res., 89: 145-151.
CrossRefDirect Link - Newton, K., J.C. Meyer, A.R. Bellamy and J.A. Taylor, 1997. Rotavirus nonstructural glycoprotein NSP4 alters plasma membrane permeability in mammalian cells. J. Virol., 71: 9458-9465.
Direct Link - Rodriguez-Diaz, J., P. Lopez-Andujar, A. Garcia-Diaz, J. Cuenca, R. Montava and J. Buesa, 2003. Expression and purification of polyhistidine-tagged rotavirus NSP4 proteins in insect cells. Protein Exp. Purif., 31: 207-212.
CrossRefDirect Link - Sharifi, Z., B. Yakhchali and M.S. Shahrabadi, 2005. Expression and one step purification of the full-length biologically active, Nsp4 of human rotavirus WA strain. Int. J. Mol. Med. Adv. Sci., 1: 206-212.
Direct Link - Tian, P., J.M. Ball, C.Q. Zeng and M.K. Estes, 1996. The rotavirus nonstructural glycoprotein NSP4 possesses membrane destabilization activity. J. Virol., 70: 6973-6981.
Direct Link - Tian, P., M.K. Estes, Y. Hu, J.M. Ball, C.Q. Zeng and W.P. Schilling, 1995. The rotavirus nonstructural glycoprotein NSP4 mobilizes Ca2+ from the endoplasmic reticulum. J. Virol., 69: 5763-5772.
Direct Link - Zhang, M., C.Q.Y. Zeng, A.P. Morris and M.K. Estes, 2000. A functional NSP4 enterotoxin peptide secreted from rotavirus-infected cells. J. Virol., 74: 11663-11670.
CrossRefDirect Link