
Samya Hussein Mohammad
Department of Zoology, Faculty of Science, Port Said University, Egypt
Pakistan Journal of Biological Sciences
Year: 2015 | Volume: 18 | Issue: 5 | Page No.: 204-214







ABSTRACT
The quality requisites of shellfishes are primarily dependent on the water quality, assuring a healthy product and a safe consumption. Water (moisture) content of five species from two different environments was conducted for a period of one year (July, 2010-June, 2011) for this purpose. Among the most exploitable and economically important Egyptian species, three brackish (the bivalves Cerastoderma glaucum, Fulvia fragilis and the gastropod Thais carinifera) and two salt water (the bivalves Donax semistriatus and Donax trunculus) shellfishes were chosen. Estimation on monthly basis revealed the incompatibility of moisture content with spawning period. Present results proved that water content varied according to the environmental factors rather than spawning seasons. Marine species had lower demand of water than brackish species. The differences in the average of the body water content between genera [Donax sp. (0.13), C. glaucum (0.75), F. fragilis (2.35) and T. carinifera (3.68)] was greater than that within the same genus [D. semistriatus (0.10) and D. trunculus (0.16)]. The gastropod T. carinifera had higher water content than the bivalves. Hierarchical clusters analysis was a good tool to differentiate moisture content between genera, as well as between species from different environments. The present study put in the perception of the probability of use water content in discrimination and classification between species.
PDF Abstract XML References Citation
Received: June 15, 2015;
Accepted: July 31, 2015;
Published: August 27, 2015
How to cite this article
Samya Hussein Mohammad, 2015. Water Content as a New Tool for Discrimination between some Shellfishes. Pakistan Journal of Biological Sciences, 18: 204-214.
DOI: 10.3923/pjbs.2015.204.214
URL: https://scialert.net/abstract/?doi=pjbs.2015.204.214
DOI: 10.3923/pjbs.2015.204.214
URL: https://scialert.net/abstract/?doi=pjbs.2015.204.214
INTRODUCTION
Moisture content is one of the most important biochemical constituents of the shellfishes. So, it is of great importance to know variations in the moisture content of them, as it may be correlated with spawning or other physiological activities or may reflect the impact of the surrounding environment on them. Most molluscs have open circulatory systems, which mean that their organs are surrounded by watery blood that contains nutrients and oxygen. Changes in watery environment represent a challenge for the tissues of these molluscs and alternatively on their water contents (Ruiz and Souza, 2008). So, we assume that moisture content may be used as a tool to differentiate between genera as well as between species from different environments (brackish and sea water). For this purpose, we select five shellfishes (the bivalves; C. glaucum, F. fragilis, D. semistriatus, D. trunculus and the gastropod T. carinifera) among the most exploitable species in Egypt and also worldwide.
The cockles are among the most familiar marine invertebrates, which include the largest and fastest growing living members of the Bivalvia class (Yonge, 1982). C. glaucum, commonly known as the lagoon cockle, is a species of Egyptian brackish water bivalve and is also quite common in Danish water, Baltic and in the Atlantic region: in the Netherlands and British Isles (Mohammad, 2002).
F. fragilis another cockle species among the brackish water bivalve and common in several other countries, such as; Turkey (Ozturk and Poutiers, 2005), Tunisia (Rifi et al., 2011), Italy, Spain (Zenetos et al., 2003), Malta (Goud and Mifsud, 2009) and Egypt (Mohammad et al., 2006).
The wedge clams, D. trunculus and D. semistriatus are one of the most economically important marine bivalves in Egypt. These saltwater species are widespread Mediterranean species. They occur usually on the coasts of Western Europe and north-western Africa. They inhabit the shallowest two meters of coastline and are commercially harvested for food (Alkaradawe et al., 2014).
Gastropods, on the other hand, represent ~2% of the marine molluscs harvested worldwide (Leiva and Castilla, 2001). Muricidae is a large family comprising 8 subfamilies represented by ~1300 species that are distributed worldwide (Houart, 2001). Despite that remarkable number of species, only a few muricids have commercial value. Among those, T. carinifera is widely distributed in the Indian Ocean, in the Red Sea, in Suez Canal in the Mediterranean Sea and in the Pacific Ocean (Ine, 1986; Gofas et al., 2001; Streftaris et al., 2005; Radwan et al., 2009), is also harvested for human consumption and has large commercial value due its cheapness and to the large amount of meat that containing.
These five species have a considerable economic importance, both in terms of aquaculture and harvested aquatic resource. Accordingly, the main purpose of this research was: (1) To evaluate water content and its percentage in these shellfishes and (2) To assess the possibility of discrimination and classification on the basis of water content.
MATERIALS AND METHODS
The study was conducted in the brackish (Timsah Lake, which is located on the Suez Canal at 30°34'N and 32°18'E) and salt-water (Idku that is located on the Mediterranean coast of Egypt, Latitude: 31°31'N, Longitude: 30°3'E). Average of salinity varied from 36-43 psu, for the brackish and saltwater, respectively. Specimens were collected at monthly intervals from July, 2010 -June, 2011. Shell measurements were made to the nearest 0.01 mm using a vernier caliper. The wet tissues were blotted and their weights (wet weights) were measured to the nearest 0.001 g with an electrical balance. Sexes were separated. The sexual dimorphism of C. glaucum and D. semistriatus is largely gonadal. Ovaries are green and dark red in females’ C. glaucum and D. semistriatus, respectively. Testes are creamy white-pale orange in males of both species. The dry weights were recorded after drying the tissue to a constant weight for 48 h at 80°C. The water (moisture) content was determined by weight difference before and after drying the samples. Percentage water (moisture) content was estimated, as percentage of dry weight to the wet weight.
Statistical analysis: Statistical analysis was evaluated by Microsoft Excel and SPSS 18 statistical packages. Mean of the different parameters were analyzed graphically and submitted to analysis of the variance (ANOVA) followed by multiple range LSD test to find any significant (p<0.05) fluctuations during the year or between the studied species. Primers for Windows (version 5.2.0) were used to determine the degree of similarity between the different species.
RESULTS
Figure 1-3 elucidate monthly significant variations (p<0.01) of water content and its percentage for both sexes of C. glaucum and D. semistriatus but the variation was not significant between sexes (p = 0.85 and 0.24, for them, respectively).
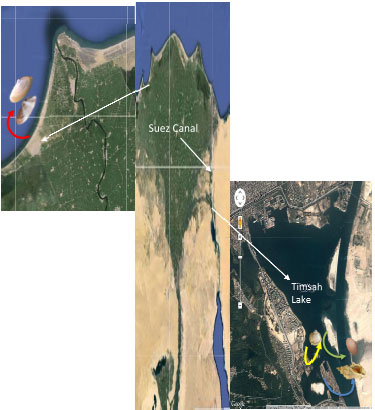 | |
| Fig. 1: | Map of the investigated area and monitoring sites in this study |
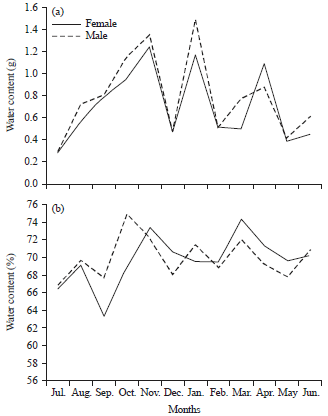 | |
| Fig. 2(a-b): | Monthly variation in (a) Water content and (b) Its percentage in both sexes of Cerastoderma glaucum |
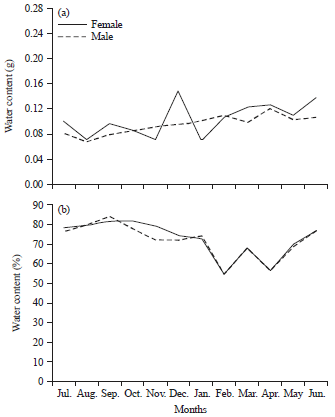 | |
| Fig. 3(a-b): | Monthly variation in (a) Water content and (b) Its percentage in Donax semistriatus |
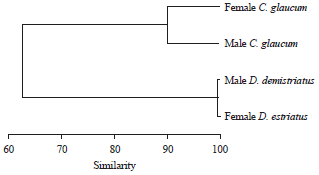
This was ascertained by the high similarity between sexes as shown in Fig. 4. However, moisture of males was slightly higher than females in C. glaucum and the reverse was true in D. semistriatus.
Water content of both sexes of C. glaucum and D. semistriatus increased with the increase in shell length, as shown in Fig. 5a and 6a. The increasing in water content of male C. glaucum was slightly higher than that of females (b = 1.87 and 1.49 for them, respectively). D. semistriatus showed also the reverse trend (b = 1.23 and 1.41 for male and female, respectively).
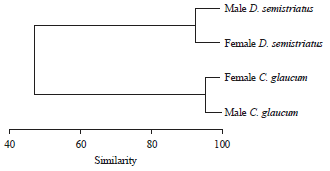 | |
| Fig. 4: | Dendrogram for hierarchical clusters analysis shows the similarity in water content between the studied species based on monthly variation during the year |
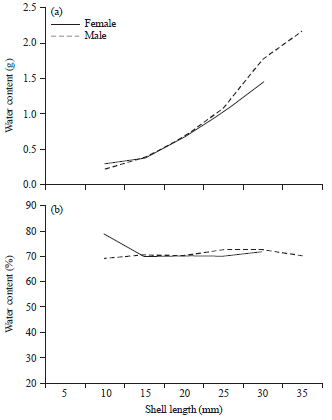 | |
| Fig. 5(a-b): | Variation in (a) Water content and (b) Its percentage with shell length in Cerastoderma glaucum |
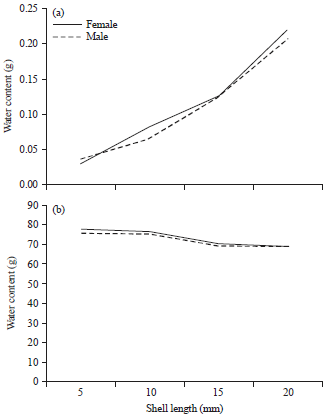 | |
| Fig. 6(a-b): | Variation in (a) Water content and (b) Its percentage with shell length in Donax semistriatus |
 | |
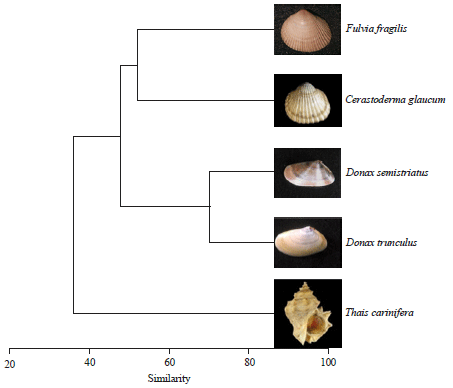
| Fig. 7: | Dendrogram for hierarchical clusters analysis shows the similarity in water content between the studied species according to the shell length |
However, there was no difference between sexes (p = 0.93 and 0.99 for C. glaucum and D. semistriatus, respectively). Generally, similarity between sexes in D. semistriatus was higher than that in C. glaucum as shown in Fig. 7. Reversely, the moisture percentage declined as the shell length of female C. glaucum, male and female D. semistriatus increased (b = -1.34, -2.58 and -3.23 for them, respectively) as shown in Fig. 5b and 6b. However, variation of moisture percentage with shell length was not noticeable in C. glaucum due to weak correlation (R2 = 0.3 for both sexes).
The bivalve water contents increased also with weight (Fig. 8-9). Females were more watery than males especially in the largest size.
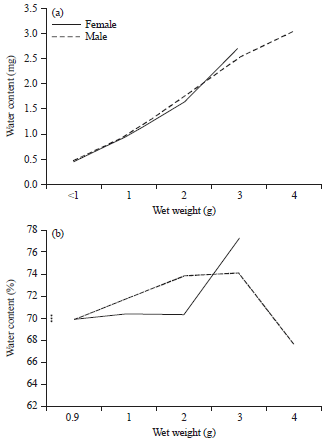 | |
| Fig. 8(a-b): | Variation in (a) Water content and (b) Its percentage with wet weight in Cerastoderma glaucum |
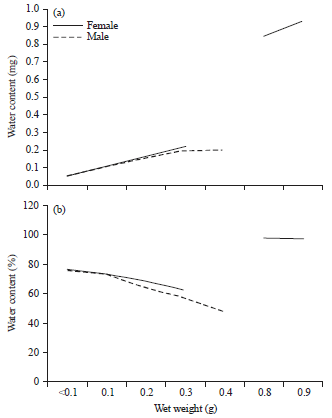 | |
| Fig. 9(a-b): | Variation in (a) Water content and (b) Its percentage with wet weight in Donax semistriatus |
 | |
| Fig. 10(a-b): | Total average of (a) Water content and (b) Its percentage in Cerastoderma glaucum and Donax semistriatus |
This was clear from the negative relationship of its percentage with size that appeared in male C. glaucum and male D. semistriatus (b = -0.22 and -7.33 for them, respectively).
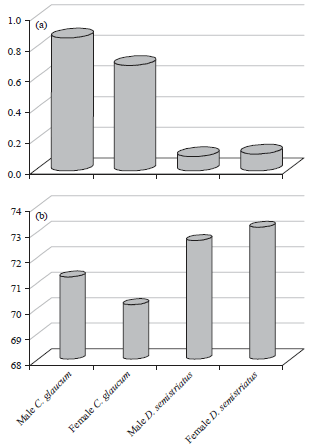
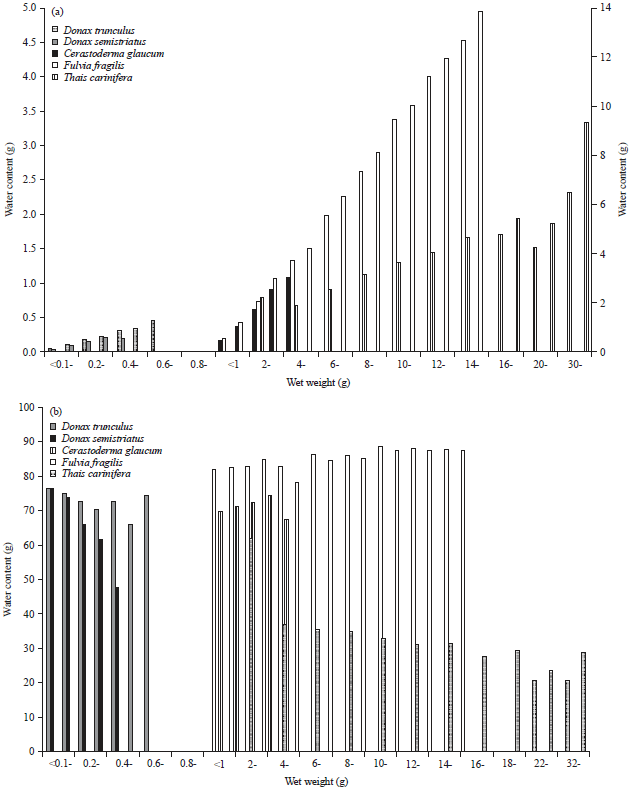
Generally, the average water content and its percentage in both sexes of the two bivalves were graphed in Fig. 10. The difference between sexes was little and non significant (p>0.05). Hence, the data was pooled and compared with the other molluscs. The gastropod T. carinifera had the highest water content as seen in Fig. 11a. The maximum and minimum water contents of the studied species were 6.56 (Nov.) and 2.33 (Sep.), 2.58 (Jan.) and 0.84 (May), 1.36 (Jan.) and 0.27 (Jul), 0.46 (May) and 0.04 (Nov.) and 0.12 (Apr.) and 0.06 (Aug.) for T. carinifera, F. fragilis, C. glaucum, D. trunculus and D. semistriatus, respectively. Significant variations (p<0.01) were detected between all species except that between D. trunculus and each of C. glaucum (p = 0.17) and D. semistriatus (0.86). Water content percentage of F. fragilis exhibited monthly little variation with a distinct drop (70.18%) in Oct. (Fig. 11b). The highest percent of water content of D. semistriatus (83.31) appeared in Sep. while the lowest (54.94 and 56.49) in Feb. and Apr. Moisture percentage ranged between 56.20 and 76.78, 65.42 and 73.51 and 21.61 and 68.85 in D. trunculus, C. glaucum and T. carinifera, respectively.
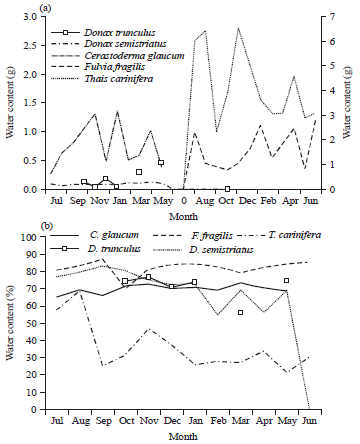 | |
| Fig. 11(a-b): | Monthly variation in (a) Water content and (b) Its percentage in the studied species |
No significant variations were detected between the bivalves C. glaucum and D. trunculus (p = 0.57), C. glaucum and D. semistriatus (p = 0.17) and D. trunculus and D. semistriatus (p = 0.63), but the others exhibited significant variations (p<0.05).
The water content increased as the shell length increased (Fig. 12a). T. carinifera showed a different trend, water content increased till shell length 60 mm, after which it declined up to 70 mm, thereafter, it began to increase again. However, F. fragilis showed the maximum increase in water content (b = 2.59), as the shell length increased by 1 mm. It was followed by C. glaucum (b = 1.77) and the lowest was D. semistriatus (b = 1.36). No obvious variation was detected in the moisture percentage with the increase of length in C. glaucum and F. fragilis (Fig. 12b). T. carinifera exhibited negative correlation with the shell size, the maximum water percentage was recorded in the smallest snail (62.24%), it declined to 28.95% in the largest size. Negative correlation with size was also recorded for the clams D. trunculus and D. semistriatus. These results are well represented in Fig. 13. It separated the investigated species into three groups. The first group was the brackish bivalves (F. fragilis and C. glaucum) that had a positive correlation with shell length, the second group was the marine bivalves (D. trunculus and D. semistriatus) that had a negative correlation and isolated the gastropod T. carinifera in the third group.
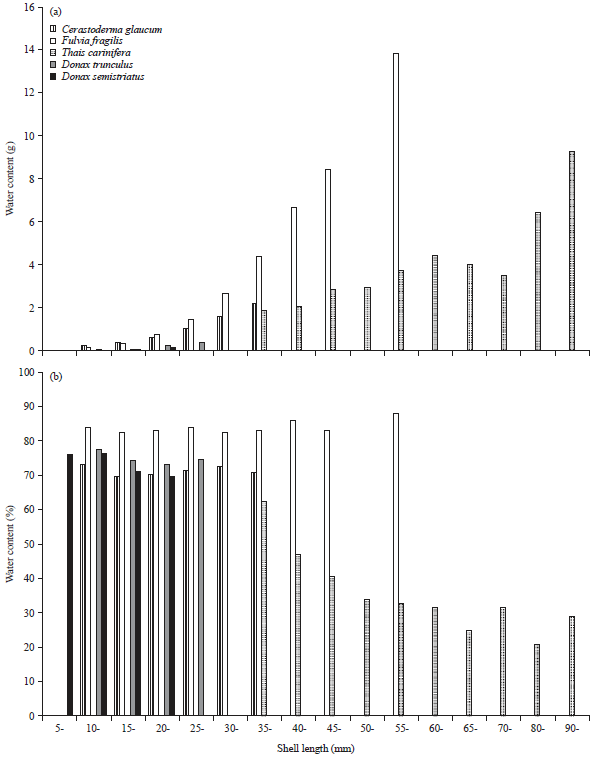 | |
| Fig. 12(a-b): | Variation in (a) Water content and (b) Its percentage with shell length in the studied species |
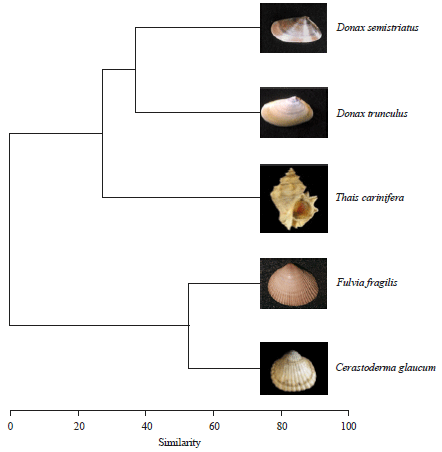
Depending on weight, there was obvious positive correlation with the wet weight (Fig. 14a). F. fragilis exhibited the highest moisture content specially in the heaviest animals while D. trunculus and D. semistriatus contained the lowest. Hierarchical clusters analysis (Fig. 15) also isolate F. fragilis and C. glaucum in a group dissimilar with Donax sp. (similarity = 0). The percentage of water content (Fig. 14b) shows little variations with the increasing in weight except T. carinifera that exhibited a negative relationship (b = -1.24) with weight.
The least values of water content were estimated in the marine bivalves (0.10 and 0.16 for D. semistriatus and D. trunculus, respectively) as shown in Fig. 16a. It gradually increased to 0.75 and 2.35 g in the brackish bivalves C. glaucum and F. fragilis, then it reached to the highest (3.68 g) in the gastropod T. carinifera. This variation between genera was highly significant (p<0.01) except that between D. semistriatus and D. trunculus (p = 0.75). Meanwhile, percentage of water content of T. carinifera represented the minimum value (32.89 g) and the maximum value was represented by F. fragilis (Fig. 16b).
 | |
| Fig. 13: | Dendrogram for hierarchical clusters analysis shows the similarity in water content between the studied species based on shell length |
Also, the variation between species was highly significant except that between D. semistriatus and D. trunculus (p = 0.50).
DISCUSSION
The present study revealed that monthly variation in water content of male C. glaucum was slightly higher than female, while the reverse was the case in D. semistriatus. This means that the monthly water demand differs between sexes of brackish water than that of the sea. This result was ascertained from the hierarchical clusters analysis that separated the brackish from marine genera at similarity of 50%.
Both sexes of marine and brackish water showed increase in water content as the length and weight increased. However, their moisture percent declined with them. Female C. glaucum was the only exception that showed a positive correlation. This may mean that females’ C. glaucum became more watery at the larger size than males.
Generally, variation in the water content between sexes was not significant. This was previously noticed by Jayabal and Kalyani (1987), who stated that different body fractions of both male and female M. meretrix showed similar trend in water content. So, this data was pooled and compared with the other genera. The present results revealed monthly significant variation between T. carinifera and F. fragilis from one side and between them and the other bivalves from the other hand. This was clear from the highest water content that recorded in T. carinifera followed by F. fragilis then C. glaucum. Hence, the maximum water content was recorded in brackish shellfishes (T. carinifera, F. fragilis and C. glaucum), while, the least water content was estimated in the marine bivalves (D. semistriatus and D. trunculus). We can relate this to the surrounding environment. As D. semistriatus and D. trunculus live in higher salinity (43 pus), it may accumulate less water than that leave in the brackish water (36 pus) according to their osmoregulation. A similar result was previously recorded by Jayabal and Kalyani (1987). They mentioned that, at lower salinity, water accumulated into the body.
Generally the present study revealed a distinct monthly pattern of variation in the water content of all studied species. Some authors related this variation to the spawning season. Ogogo (2004) attributed the fall in moisture to the spawning of the animal. Meanwhile, Manhas et al. (2013) observed that moisture content was high during the peak spawning. Taylor and Venn (1979) previously interpreted this. The rapid increase in moisture weight corresponds with the animal absorbing more water to fill the lumen created in the gonad after the release of the gamete. Recently, Udoh and Abiaobo (2014) confirmed that as they found the shellfish species absorbed more water after spawning. The present studied species characterized by continuous spawning. Spawning of C. glaucum occurred most of the year, without prolonged periods of reproductive inactivity (Mohammad, 2002; Machreki-Ajmi et al., 2013). The continuity of spawning was also recorded for the other shellfishes F. fragilis (Mohammad, 2002), T. carinifera (Radwan et al., 2009), D. semistriatus and D. trunculus (Ali et al., 2009; Alkaradawe, 2014).
 | |
| Fig. 14(a-b): | Variation in (a) Water content and (b) Its percentage with wet weight in the studied species |
The irregular annular trends in the water content of the present studied species did not coincide with the spawning season. So, the present results disagree with the previous assumptions but consistent with Gharsallah et al. (2010). They mentioned that among the biochemical component analyzed in the Hexaplex truculus, the protein, lipid and carbohydrate contents showed marked variations throughout the reproductive cycle, whereas, the moisture content did not show significant oscillations. Moreover, we assume that the gonad did not represent a large fraction of the body and hence, the change in its water content did not impact on the total water content. This assumption coincides with the early finding of Jayabal and Kalyani (1987). They estimated water content in four selected body parts (digestive diverticula, adductor muscle, foot and gonad) of M. meretrix and concluded that gonad had the minimum water content among the four body fractions.
 | |
| Fig. 15: | Dendrogram for hierarchical clusters analysis shows the similarity in water content between the studied species based wet weight |
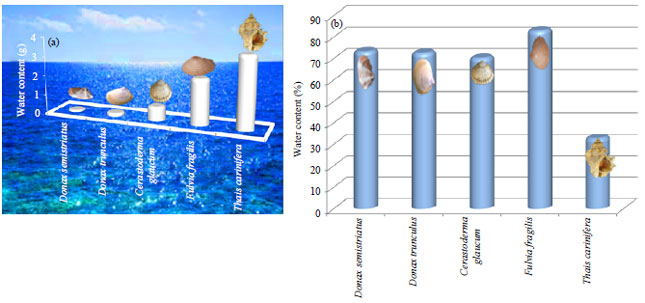 | |
| Fig. 16(a-b): | Average variation in (a) Water content and (b) Its percentage in the studied species |
Accordingly, we can attribute the variation in the water content of the present studied shellfishes to the environmental factors rather than spawning seasons. Water content was influenced by food availability and salinity (Jayabal and Kalyani, 1987). Starvation was also among factors that affected water content. Riley (1976) mentioned that starvation involved dry weight losses and a general increase in the water content of the body of Crassostrea gigas.
CONCLUSION
In a conclusion, demand of water varies according to the surrounding environment and genera. The marine species contained lower water content than the brackish species. The gastropod had higher water content than the bivalves and the variation was not significant between the species of the same genus or between sexes. Allah has created every (living) creature from water and of them are those that move on their bellies and of them are those that walk on two legs and of them are those that walk on four. Allah creates what He wills. Indeed, Allah is over all things competent (Surat An-Nūr (The Light), 45).
ACKNOWLEDGMENTS
The author expresses her deepest gratitude to E. Soliman K. for useful comments and valuable suggestions on the final manuscript. I also wish to thank E. Abd El Rahman S. who collaborated in the organizing the data. Special thanks to Dr. Rabab, I. and Dr. Nancy, A. for their valuable assistance. The author would like to thank the valuable comments of two anonymous referees.
REFERENCES
- Alkaradawe, R.M.I., S.H. Mohammad, S.Z. Mohamed and Z.S. Morsy, 2014. Population structure, condition index and age structure of the Mediterranean carpet shell clams Donax semistriatus and Donax trunculus (Mollusca: Bivalvia) in Idku, Egypt. Egypt. J. Aquat. Biol. Fish., 18: 25-48.
Direct Link - Goud, J. and C. Mifsud, 2009. Fulvia fragilis (Forsskal in Niebuhr, 1775) (Bivalvia: Cardiidae), an alien species new to the Maltese malacofauna. Aquatic Invasions, 4: 389-391.
CrossRefDirect Link - Jayabal, R. and M. Kalyani, 1987. Seasonal variations in biochemical constituents of different body components of Meretrix meretrix (L.). Mahasagar, 20: 65-69.
Direct Link - Leiva, G.E. and J.C. Castilla, 2001. A review of the world marine gastropod fishery: Evolution of catches, management and the Chilean experience. Rev. Fish Biol. Fish., 11: 283-300.
CrossRefDirect Link - Machreki-Ajmi, M., T. Rebai and A. Hamza-Chaffai, 2013. Reproductive strategy in a littoral population of the cockle Cerastoderma glaucum from the Gulf of Gabes area (Southeastern Tunisia). J. Shellfish Res., 32: 733-738.
CrossRefDirect Link - Manhas, P., S. Langer and G. Singh, 2013. Studies on water and lipid distribution pattern in Paratelphusa masoniana (Henderson) (female), an edible freshwater crab from Jammu region of j&k (India). Int. J. Adv. Res., 1: 245-251.
Direct Link - Ozturk, B. and J.M. Poutiers, 2005. Fulvia fragilis (Bivalvia: Cardiidae): A lessepsian mollusc species from Izmir bay (Aegean sea). J. Mar. Biol. Assoc. UK., 85: 351-356.
CrossRefDirect Link - Radwan, N.A., S.H. Mohammad, S.Z. Mohammed and A.E. Yaseen, 2009. Reproduction and gonad development of gastropod thais carinifera in Lake Timsah, Suez Canal, Egypt. Egypt J. Aquat. Biol. Fish., 13: 53-67.
Direct Link - Rifi, M., G. Le Pennec, M.B. Salem and J. Ben Soissi, 2011. Reproductive strategy of the invasive cockle Fulvia fragilis in the Bay of Tunis (Tunisia). J. Mar. Biol. Assoc. UK., 91: 1465-1475.
CrossRefDirect Link - Ruiz, J.L. and M.M. Souza, 2008. Osmotic stress and muscle tissue volume response of a freshwater bivalve. J. Comparat. Biochem. Physiol. Part A, 151: 399-406.
CrossRefDirect Link - Streftaris, N., A. Zenetos and E. Papathanassiou, 2005. Globalisation in marine ecosystems: The story of non-indigenous marine species across European seas. Oceanogr. Mar. Biol. Ann. Rev., 43: 419-453.
Direct Link - Taylor, A.C. and J.J. Venn, 1979. Seasonal variation in weight and biochemical composition of the tissues of the queen scallop, Chlamys opercularis, from the Clyde sea area. J. Mar Biol. UK., 59: 605-621.
CrossRefDirect Link - Udoh, J.P. and N.O. Abiaobo, 2014. Condition index, meat yield and population structure of the marine gastropod, Thais coronata, off cross river estuary, Nigeria. J. Adv. Life Sci. Technol., 23: 24-31.
Direct Link - Yonge, C.M., 1982. Functional morphology and evolution in the Tridacnidae (Mollusca: Bivalvia: Cardiacea). Rec. Aust. Mus., 33: 735-777.
CrossRefDirect Link